Biologie evolutivă a dezvoltării
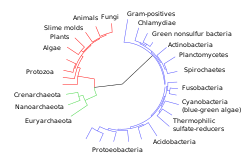
Biologia evolutivă a dezvoltării (în engleză Evolutionary Developmental Biology [1] [2] , din care termenul popular evo-I) este disciplina științifică care analizează o structură evolutivă și o funcție a genomului (adică setul complet cu tot ADN - ul conținut în o celulă). Se ocupă cu investigarea relației dintre dezvoltarea embrionară și fetală a unui organism ( ontogeneza ) și evoluția populației sale ( filogenia ) [3] .
Această disciplină se bazează pe ipoteza că mutațiile care afectează genele implicate în controlul dezvoltării embrionare (ontogeneza) pot da naștere la noi caractere la adult ( fenotip ), care prin reproducere va transmite noilor caractere descendenților, începând astfel procesul evolutiv al acestor personaje în populație. Prin urmare, un rol principal în evoluția biologică este atribuit genelor care controlează și reglează dezvoltarea embrionului; diversificarea progresivă ( biodiversitatea ) ființelor vii ar fi explicată prin schimbări funcționale în controlul și reglarea genelor care reglementează dezvoltarea embrionară.
Evoluția biologică prin selecție nu mai este privită pur și simplu ca schimbarea prin generații succesive de genotipuri și fenotipuri prin mutații, încrucișări și fluxuri migratorii, ci și ca o schimbare în reglarea genelor implicate în procesele care reglementează dezvoltarea. Biologia evolutivă a dezvoltării este considerată capabilă să răspundă la întrebarea privind trăsăturile neadaptative prezente în populații [4] .
Acest lucru a provocat o schimbare fundamentală în paradigma evolutivă: în trecut se credea că biodiversitatea depinde de modificări mai mult sau mai puțin semnificative în numărul și secvența genelor, de la începutul anilor nouăzeci , comunitatea științifică a fost de acord că apariția de noi personaje ( prin urmare, apariția unor funcții noi) în organismele unei populații, precum și cauzate de apariția unor gene noi, este cauzată și de modificări ale reglării acelorași gene deja existente în genomul diferitelor organisme [5] . Prin urmare, schimbarea modului de reglare a expresiei genelor ar duce la apariția de noi funcții în organismele vii [4] [6] [7] [8] [9] [10] [11] .
Nașterea biologiei evolutive a dezvoltării este de obicei urmărită în anii 1990 și este de obicei atribuită unificării biologiei evoluției și a biologiei dezvoltării .
Principiile biologiei dezvoltării evolutive
Biologia evolutivă a dezvoltării nu este o alternativă la teoria evoluției prin selecție naturală: selecția este văzută ca mecanismul de control a posteriori al variantelor aleatorii care pot apărea în cadrul unei populații. Selecția este considerată ca un mecanism de bază, care nu poate fi înlocuit de mecanismele descoperite ulterior ( deriva genetică , fluxul de gene , efectul fondator etc.) [12] .
Biologia evolutivă a dezvoltării teoretizează că, pe lângă forțele externe care acționează asupra organismului (forțele de mediu), există și forțe care pot modifica organismele vii din interior (forțe genomice).
Așa-numitele forțe „interne“ sunt urmărite înapoi la genomice cifra de afaceri , ceea ce reprezintă o rearanjare și / sau remodelare a genomului cauzate de proprietățile unor elemente ale acestora; aceste elemente pot:
- trecerea de la o parte a genomului la alta ( transpunere );
- convertirea în gene noi (conversie genetică);
- convertirea spontană la diferite baze azotate ( dezaminare , metilare );
- distribuție inegală la descendenți în timpul duplicării celulelor germinale în meioză ( încrucișare inegală);
- răspund la stimulii de mediu interacționând cu aceștia prin calea transducției semnalelor de mediu către genom .
- mutații spontane care nu pot fi atribuite acțiunii mutagenilor externi sau a agenților intercalatori.

Temporar, fenomenele de rotație pot apărea în orice moment din viața organismului. Pentru ca efectele lor să fie transmise descendenților, aceste fenomene trebuie să apară în celulele liniei germinale , în special la nivelul celulelor responsabile de formarea gametilor ( gametogeneza ). Ele pot apărea și în timpul formării zigotului .
Fenomenele de rotație genomică, la nivel de genom, pot duce la:
- reutilizarea secvențelor de nucleotide, destinate modulelor; modulele din timpul dezvoltării ar fi utilizate diferit în diferite grupuri taxonomice;
- cooptarea genelor, adică utilizarea diferită a acelorași gene de către diferite organisme;
- promovarea schimbărilor evolutive, cum ar fi: duplicarea genelor , divergența între gene și transferul orizontal de gene - transferul ne-ereditar al aceleiași gene de la un organism la altul.
Prin urmare, fluctuația genomică, dacă se produce, ar contribui la originarea unor variații genetice care, la nivelul populației, ar duce la diferențierea ființelor vii și, prin urmare, la biodiversitate .
Domenii de studiu

Biologia evolutivă a dezvoltării se concentrează pe studiul variațiilor care pot apărea în timpul dezvoltării embrionare a unui organism, în special asupra erorilor și mutațiilor care pot apărea și care ar putea conduce la reprogramarea dezvoltării embrionare, originând noi structuri sau noi căi metabolice la adult.
De asemenea, investighează evoluția ființelor vii în raport cu dezvoltarea embrionară a acestora, concentrându-se pe embriologia comparativă, pentru a clarifica diferențele și asemănările dintre evoluțiile embrionare ale diferitelor organisme.
Studiile de teren ale biologiei evolutive a dezvoltării, privind dezvoltarea embrionară, vezi Xenopus laevis (un broască africană) printre protagoniști: acest lucru se datorează dimensiunii mari a celulelor sale de ou, care facilitează studiul și observarea empirică.; Xenopus laevis , împreună cu alte organisme, este, de asemenea, organismul model al studiilor privind evoluția sistemului imunitar și funcționarea căilor care reglează stresul celular.
Dovezi despre Evo-devo
Coopțiune genică

Cooptarea genică este un mecanism evolutiv în care aceleași gene, în aranjamente diferite, dau naștere unor funcții diferite [5] . Gena nu este nici modificată, nici mutată. Cooptarea este în general recunoscută prin secvențierea genomului și observarea prezenței genei într-o poziție diferită de cea a genomului de referință. Alte condiții de cooptare sunt date de mutații care modifică funcționarea elementelor care reglează expresia genei, cum ar fi: factori de transcripție , promotori , potențatori , ARN nucleolar mic și, de asemenea, molecule de semnal reglator , modificări post-translaționale .
Exemple de cooptare genică sunt:
- Gena homeotică Distalless, implicată în dezvoltarea membrelor muștei fructului Drosophila melanogaster ; la unii fluturi este cooptat pentru dezvoltarea unor pete în formă de ochi pe aripi [7] [13] .
- Câștigul funcțional specific omului, un element de reglementare aparținând categoriei de potențatori , a suferit nouă mutații cumulative; aceste mutații au cooptat toate genele responsabile de dezvoltarea membrelor anterioare ale protoprimatelor, rezultând mai degrabă dezvoltarea mâinii umane decât dezvoltarea mâinii primatelor [14] . De fapt, genele pentru construirea mâinii primatelor sunt identice cu cele ale omului, se consideră că modificarea câștigului funcțional specific omului este principala cauză a diferențelor dintre mâna umană și mâna primatelor [15] [16 ] . ] .
Mecanismul cooptării genetice a dat naștere la noi programe de cercetare științifică, deschizând astfel noi căi pentru înțelegerea proceselor evolutive, în special a clarificat numeroase puncte referitoare la evoluția sistemului imunitar și funcționarea căilor care reglează stresul. [17] .
Mecanismele cooptării s-au dovedit importante și în sectorul medical, în care au contribuit la clarificarea funcționării fenomenului angiogenezei , adică a capacității tumorilor de a-și crea propria rețea de vase de sânge, necesare supraviețuirii a tumorilor în sine. Celulele, în timpul transformării lor în tumori, suferă cooptări ale funcțiilor genelor prezente în celula sănătoasă și, din studii medicale, se pare că înțelegerea acestor opțiuni este una dintre cheile luptei împotriva cancerului [18] .
Dezvoltare conservatoare și modulară
Biologia evolutivă a dezvoltării evidențiază modul în care diferențele dintre un organism și altul nu se pot atribui prezenței simple a unor gene specifice pentru unul sau altul organism, ci mai degrabă depind de modul în care este reglementată expresia, în cursul dezvoltării embrionare și timp evolutiv , al acelorași gene prezente în diferite organisme.
Secvențele genetice sunt văzute ca module, deși aceste module sunt întotdeauna aceleași, sunt aranjate și organizate diferit în diferite organisme. Prin urmare, genomul unui organism este structurat în module interdependente, variația unui singur modul afectează întregul organism, de exemplu: Antennapedia este un modul Hox care controlează poziționarea picioarelor în dezvoltarea embrionară a insectei Drosophila melanogaster , ectopica sa expresia, adică expresia acestei forme, în timpul dezvoltării embrionare, într-o regiune anatomică diferită de cea normală, conduce la formarea picioarelor deplasate [19] .


Toate aceste module reprezintă „cutia de instrumente” disponibilă pentru fiecare organizație; fiecare organism, în timpul fazei sale de dezvoltare embrionară, folosește aceste module ca „instrumente” pentru a-și construi propria formă ( fenotip ), un organism va dezvolta un fenotip în loc de altul în funcție de modul în care va folosi „instrumentele” la nivelul său dispoziție în timpul dezvoltării sale embrionare, de exemplu: în Drosophila melanogaster al treilea segment toracic constă dintr-o pereche de picioare și o pereche de bascule , variația genei ultrabithorax , o altă genă Hox, transformă al treilea segment toracic într-un al doilea segment toracic echipat cu aripi în loc de balansoare [20] , deoarece genomul insectelor , fiind organizat prin module care dau interacțiuni tipice pentru a forma un fenotip de forme de insecte, conține posibilitatea dezvoltării a 2 sau 4 aripi, în diferitele subdiviziuni taxonomice, de ex. familii , această organizare a genomului a apărut în cursul evoluției insectelor , iar coca a „stabilit” să dezvolte un anumit fenotip caracteristic pentru fiecare grupare taxonomică. Faptul că această „cutie de instrumente” este practic identică la diferiți taxoni pentru insecte (această asemănare se numește conservare sau forme comune ) arată în favoarea unei origini comune sau a strămoșilor comuni pentru toate insectele [21] .
În cursul evoluției, divergențele biologice și modificările morfologice, care au condus la biodiversitatea actuală, sunt însoțite de un grad ridicat de conservare a modulelor genetice care determină anumite caractere.
Exemplele sunt date de Hox , care sunt module genetice responsabile de dezvoltarea segmentară a corpului animalului, dezvoltarea corectă a axei corpului și poziția anatomică în care organismul își va dezvolta apendicele (picioare, aripi, antene, membre etc.) ..); au rămas stabile în aproape toate speciile, chiar și în organismele care s-au diversificat de milioane de ani.
Unele dintre Hox recunoscute și studiate și care mărturisesc bine această conservare sunt:
- Pax6 (mamifere) și Eyeless / Toy (artopode) , sunt module genetice aparținând categoriei Hox ; Structura, conformația și funcția Pax6 sunt identice cu cea a Eyeless / Toy, de fapt ambele sunt responsabile de formarea ochilor la animale, aceste două module genetice sunt nealterate, la toate speciile de animale; S-a arătat că înlocuind Eyeless / Toy din Drosophila (muscă) cu cel al șoarecelui , numit Pax6, se obține și dezvoltarea corectă a ochiului [22] . Se crede că aceasta este o dovadă genetică că mamiferele și insectele posedă un strămoș comun îndepărtat, de la care aceste două filuri au moștenit aceeași formă. Acest modul a fost numit Pax6 la mamifere și artropode fără ochi / jucărie, deoarece fiecare model experimental are propria sa nomenclatură pentru denumirea secvențelor de nucleotide, în plus, se efectuează descoperirea secvenței de nucleotide și atribuirea numelui la secvență. funcţie.
- Distalless , implicat în dezvoltarea membrelor, este întotdeauna prezent și la șerpi , dar a suferit o variație a modului său de exprimare, deci nu mai este exprimat în regiunile în care picioarele ar trebui să se dezvolte, ca urmare a lipsei sau insuficienței sale de exprimare . Din această cauză, șerpii nu mai sunt capabili să dezvolte picioare [23] .
Importanța medicală a conservării
Fenomenele de conservare, prin procesul evolutiv , ale modulelor genetice în dezvoltarea organismelor vii [24] , conduc la noi dovezi în sectorul medical, de exemplu: descoperirea funcționării modulului nodal (un modul genetic ) a fost fundamental pentru înțelegerea și prevenirea adecvată a multor boli de inimă.
nodal este un modul genetic responsabil pentru spirală normală sau inversată în anatomia corpului organismelor vii [25] .
Spiralizarea este procesul prin care organele sunt poziționate în direcția corectă în interiorul corpului [26] , de exemplu: la mamifere inima este înclinată longitudinal cu vârful spre stânga, ficatul este poziționat spre dreapta și splina pe stânga .
Structura, conformația și funcția nodalului sunt identice la gastropode , broaște [27] , la om [28], de fapt reprezintă o formă clasică numită comună sau conservată; în special în gastropode este responsabil pentru spirala spre dreapta cochiliei.
O modificare a nodului provoacă o spirală inversată în timpul dezvoltării organismului [29] [30] , de exemplu: dacă acest lucru se întâmplă la om, inima, în direcția longitudinală, va fi înclinată cu vârful spre dreapta, ficatul o splină stângă și dreaptă.
Gastropodele care posedă un modul nodal inversat își construiesc învelișul rotindu-se în sens invers acelor de ceasornic în locul acelor de ceasornic.
Oamenii care posedă inversat fața modulului nodală un risc mai mare de boli si probleme cardiace cardiovasculare [31] (ca toate organele sunt poziționate în spate, ca și în cazul în care acestea au fost în oglindă), identificarea acestei spirale Inverted nodală , în general , se produce atunci când este prea târziu, atunci când persoana suferă primele probleme cardiace, identificarea timpurie a purtătorilor nodali inversați le-ar permite acestor indivizi să întreprindă o prevenire precisă și, prin urmare, un risc mai mic de probleme cardiovasculare.
Constrângeri de dezvoltare

Organizarea dezvoltării embrionare prin intermediul acestor module implică limite dezvoltării embrionare a organismelor, aceste limite pot fi de natură anatomică, de exemplu: la mamiferele euplacentate fătul trebuie să treacă prin canalul vaginal pentru a se naște, dimensiunea organism la naștere nu pot depăși cele ale canalului vaginal, din acest motiv, de exemplu: capul ființelor umane nu poate deveni „prea” mare și de fapt unele organisme, precum ființa umană, își pun capăt dezvoltării în afara uterului matern; Moartea în timpul unei nașteri naturale a unui făt care s-a dezvoltat cu un cap prea mare a împiedicat, în cursul evoluției umane, orice posibilă dezvoltare umană către această caracteristică. Sau pot fi de natură genomică, de exemplu: mamiferele au toate un număr de vertebre cervicale egal cu șapte, o variație a modulului genetic responsabil de dezvoltarea numărului de vertebre, care în timpul dezvoltării embrionare a dus la o variație a numărului vertebrelor cervicale, ar avea repercusiuni într-o variație a celorlalte module interdependente, rezultatul este întotdeauna letal la mamifere, din acest motiv vertebrele gâtului pot ajunge la dimensiunea de 70 de centimetri în lungime ca în girafă sau pot scădea până la 1 sau 2 milimetri ca la delfin, dar la toate mamiferele trebuie să apară în mod necesar în număr de șapte.
Personaje precum: numărul variabil de vertebre cervicale la mamifere sau ființe umane cu un cap enorm, sunt de fapt prevenite de tipul de dezvoltare embrionară la care sunt supuse aceste organisme, aceste impedimente iau denumirea generică a constrângerilor de dezvoltare, dat fiind că posibilul apariția acestor trăsături în timpul dezvoltării unui organism este incompatibilă cu termenul dezvoltării în sine, aceste trăsături nu pot fi prezente în rezerva genetică a populațiilor, prin urmare nu pot fi supuse selecției sau propagării. În funcție de tipul, modalitățile și perioada de dezvoltare care disting diferitele specii, fiecare specie are propriile constrângeri de dezvoltare, posibila apariție a unei presiuni de mediu selective extrem de negative împotriva unui caracter legat de o constrângere de dezvoltare are ca efect migrația sau dispariția populației.
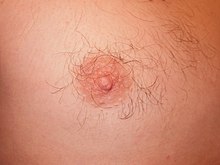
Deoarece modulele care reglementează dezvoltarea sunt interdependente, o variație a unui modul afectează toate celelalte, de exemplu o variație care a împiedicat (în timpul dezvoltării embrionare) un mamifer să dezvolte mameloane, indiferent de sexul său, ar fi incompatibilă cu sfârșitul dezvoltării în sine, aceste caractere, independent de presiunile selective la care se poate întâlni organismul, nu pot lipsi la fiecare individ al populației; Din acest motiv, găsim, în populațiile naturale, caractere care nu corespund nicio adaptare, precum prezența mameloanelor vestigiale la masculii de mamifere .
Constrângerile dezvoltării pot fi depășite, prin depășire ne referim la o modificare substanțială ( mutație ) care duce la reorganizarea modulelor genomului , o modificare care permite organismului să-și termine dezvoltarea și să-și completeze ciclul de viață cu reproducere , eludând efectiv constrângerea în sine, aceasta implică o schimbare substanțială în dezvoltarea organismului și reorganizarea modulelor care alcătuiesc genotipul acestuia; De exemplu: dacă o libelula ( Odonata ), prin intermediul mutației ultrabithorax , a suferit formarea de rockeri, în locul celei de-a doua perechi de aripi, s-ar transforma într-o insectă mai asemănătoare cu un dipter ( Diptera ); Din acest motiv, depășirea unei constrângeri de dezvoltare nu implică neapărat manifestarea unei noi trăsături în fondul genetic sau formarea unei noi specii, ci mai degrabă implică un salt evolutiv, salturile evoluate sunt evenimente care apar proporțional cu timpul de generație. de indivizi și durează sute de mii de ani să apară [32] , odată cu formarea unui nou Phylum .
Economia evolutivă
Prin acest termen înțelegem probabilitatea mai mare ca ființele vii să suporte variații de funcționalitate sau în perioada de exprimare (adică atunci când o genă începe și se exprimă și, prin urmare, își produce propria proteină sau ARN , până când gena se oprește), sau în cantitatea de proteine produse, de gene deja existente, mai degrabă decât să apară noi gene pentru a satisface noi nevoi vitale care nu au fost îndeplinite anterior.
„Când?” și „unde?”
Studiile științifice au arătat că dimensiunea genomului (adică structura completă a întregului ADN conținut într-o celulă) a unui organism ( valoarea C ) nu este legată de complexitatea organismului, dar o mare parte a biodiversității ar fi datorată la modularea diferită a expresiei acelorași gene.
S-ar datora diferențele dintre organisme în funcție de când? si unde? o genă se exprimă:
- Pentru când? înseamnă în ce fereastră de timp sau în ce perioadă de timp a dezvoltării embrionare și fetale a organismului, o genă se activează și se oprește, pentru cât timp funcționează.
- Unde? vrem să spunem în ce poziție a genomului se află acea genă, unde este situată în nucleul celulei și, prin urmare, cu ce elemente nucleare poate interacționa.
Prin urmare, aceleași gene, exprimate în perioade de timp diferite și în poziții diferite, în timpul dezvoltării embrionare a diferitelor organisme, ar fi responsabile pentru diferențele care se găsesc între organismele vii.
Relația cu biologia evolutivă
Din punct de vedere istoric, grupurile de cercetare ale biologilor în dezvoltare și biologilor evoluționisti au lucrat în paralel, evitând în același timp interacțiunile reciproce, în special în ceea ce privește concepțiile teoretice fundamentale; bibliografic, biologii de dezvoltare și biologii evoluționisti publică în diferite reviste și urmează diferite paradigme explicative.
Programele de cercetare în biologia dezvoltării evolutive încep să definească noi instrumente conceptuale pentru înțelegerea procesului evolutiv și să redefinească concepte deja cunoscute, precum: constrângeri de dezvoltare , paramorfisme sau co-opțiune genică .
Notă
- ^ Redi CA și colab. , 2004.
- ^ A scuola di evo-devo - „Școală de vară pe biologie evolutivă a dezvoltării” , pe pikaia.eu . Adus pe 7 iulie 2010 .
- ^ Raff RA, Wallace A, Carroll SB, Coates MI și Wray G, Cronicând nașterea unei discipline , în Evolution & Development 1999; 1 (1): 1-2 . Adus la 3 februarie 2021 (arhivat din original la 15 martie 2016) .
- ^ a b Pievani T, The theory of evolution today: the rediscovery of darwinian pluralism ( PDF ), la Universitatea din Basilicata - Inaugurarea anului academic , 2009. URL accesat la 6 iulie 2010 (arhivat de la adresa URL originală la 20 septembrie 2015) .
- ^ a b Mandrioli M, Aceleași gene, forme diferite , pe Pikaia, portalul evoluției . Adus la 3 februarie 2021 (Arhivat din original la 29 aprilie 2013) .
- ^ Fusco G., Minelli A., Gene, segmente și picioare: Evo-Devo în Padova ( PDF ) [ link întrerupt ] , în Notiziario al Universității din Padova 2005; 1: 10-12 .
- ^ a b P. Beldade și Paul M. Brakefield, The Genetics and Evo - Devo of butterfly wing pattern ( PDF ), în Nature 2002; 442 (3): 442-452 (arhivat din original la 27 iunie 2010) .
- ^ Brakefield, Paul M; Franceză, Vernon , 2005.
- ^ Sean B. Carrol , 2006.
- ^ Boncinelli 2007 .
- ^ Gregory 2005 .
- ^ Minelli S., Darwin în labirintul evo-devo , în Il Sole 24 ore , 20 aprilie 2010.
- ^ (EN) G. Panganiban și colab. , Rolul genei Distal-less în dezvoltarea și evoluția membrelor insectelor , în Current Biology , vol. 4, nr. 8, 1994, pp. 671-675, DOI : 10.1016 / S0960-9822 (00) 00151-2 . Adus la 26 iulie 2010 .
- ^ (EN) Shyam Prabhakar și colab. , Câștig de funcție specific omului într-un potențial de dezvoltare , în Știință , vol. 321, n. 5894, 2008, pp. 1346-1350, DOI : 10.1126 / science.1159974 . Adus la 26 iulie 2010 .
- ^ (EN) L. Duret, Galtier, N., Comentariu la „Câștigul funcției specific uman pentru un dezvoltator de dezvoltare” (PDF), în Știință, vol. 323, 2009, pp. 714c. Adus la 26 mai 2010 .
- ^ (EN) S. Prabhakar și colab. , Răspuns la comentariul despre „Câștigul funcțional specific omului într-un potențial de dezvoltare” ( PDF ), în Știință , vol. 323, 2009, pp. 714d. Adus la 26 mai 2010 .
- ^ (EN) GW Duff, Durum, SK, Cytokine , în Cytokine, vol. 1, nr. 1, 1989, pp. iv, DOI : 10.1016 / 1043-4666 (89) 91042-9 .
- ^ (EN) Liekens S., De Clercq, E.; Neyts, J., Angiogenesis: regulatori și aplicații clinice , în Biochimie Farmacologie , vol. 61, nr. 3, 2001, pp. 253-270, DOI : 10.1016 / S0006-2952 (00) 00529-3 .
- ^ Russell Peter J., Genetică , Edises, 1998, ISBN 88-7959-154-1 .
- ^ FlyBase Gene Report: Dmel \ Ubx , la flybase.org , FlyBase , 20 martie 2009. Accesat la 23 aprilie 2009 .
- ^ Bateson William, Materiale pentru studiul variației: tratat cu o atenție specială la discontinuitate în originea speciilor , Cambridge University Press, 1894.
- ^ (EN) PX, Xu, Woo, I., Her, H. Beier, DR, Maas, RL, omologii de șoarece Eya ai ochilor Drosophila absenți necesită gena Pax6 pentru exprimare în cristalin și placod nazal, în Development, vol. 124, 1997, pp. 219-231.
- ^ (EN) MJ Cohn, Tickle, C., Baza de dezvoltare a limbilității și modelarea axială la șerpi, în Nature, vol. 399, 1999, pp. 474-479.
- ^ ( EN ) Levin M, et al, Evolutionary conservation of mechanisms upstream of asymmetric Nodal expression: reconciling chick and Xenopus , in Dev Genet , 2998, 23:185-193.
- ^ Entrez Gene database , su ncbi.nlm.nih.gov .
- ^ ( EN ) Hamada H, et al, Establishment of vertebrate left-right asymmetry , in Nature Reviews Genetics , 2002, 10.1038/nrg732.
- ^ ( EN ) Yu JK, et al., An amphioxus nodal gene (AmphiNodal) with early symmetrical expression in the organizer and mesoderm and later asymmetrical expression associated with left-right axis formation , in Evolution & Development , 2002, 4(6):418-25.
- ^ ( EN ) Conlon FL, et al., A primary requirement for nodal in the formation and maintenance of the primitive streak in the mouse , in Development , 1994, 120: 1919-1928.
- ^ ( EN ) Zhou X, et al., Nodal is a novel TGF-beta-like gene expressed in the mouse node during gastrulation , in Nature , 1993, 361: 543-547.
- ^ ( EN ) Beck S, et al., Extraembryonic proteases regulate Nodal signalling during gastrulation. , in Nature Cell Biology , 2002, 4: 981-985.
- ^ ( EN ) Mohapatra B, et al., Identification and functional characterization of NODAL rare variants in heterotaxy and isolated cardiovascular malformations , in Human Molecular Genetics , 2009, 1;18(5):861-71.
- ^ Telmo Pievani, La variabile tempo nella teoria dell'evoluzione , su videoscienza.it , 15 marzo 2010 (archiviato dall' url originale il 5 febbraio 2013) .
Bibliografia
Libri
- Barnes RS, Calow P., Olive PJ, Invertebrati una nuova sintesi , Zanichelli, 1990, ISBN 88-08-07206-1 .
- Edoardo Boncinelli , Perché non possiamo non dirci darwinisti , Rizzoli, 2009, ISBN 978-88-17-03425-8 .
- Boncinelli E., Tonelli C., Dal moscerino all'uomo: una stretta parentela , Sperling & Kupfer, 2007, ISBN 978-88-6061-071-3 .
- Carroll SB, Infinite forme bellissime. La nuova scienza dell'Evo-Devo , Codice Edizioni, 2006, ISBN 978-88-7578-037-1 .
- editore: Codice Edizioni Carroll SB, Al di là di ogni ragionevole dubbio. La teoria dell'evoluzione alla prova dell'esperienza , ISBN 978-88-7578-089-0 .
- Capanna E., Pievani T., Redi CA, Chi ha paura di Darwin? , Ibis, 2006, ISBN 978-88-7164-209-3 .
- Richard Dawkins , Il più grande spettacolo della terra , Mondadori, 2010, ISBN 978-88-04-59552-6 .
- Girotto V., Telmo Pievani , Vallortigara G., Nati per credere. Perché il nostro cervello sembra predisposto a fraintendere la teoria di Darwin , Codice, 2008, ISBN 88-7578-110-9 .
- Gould SJ, La struttura della teoria dell'evoluzione , Codice Edizioni, 2008, ISBN 978-88-7578-000-5 .
- ( EN ) Gregory TR, The evolution of the genome , Elsevier/Academic Press, 2005, ISBN 0-12-301463-8 . URL consultato il 7 luglio 2010 (archiviato dall' url originale il 23 novembre 2010) .
- Neil A. Campbell, Jane B. Reece, Eric J. Simon, L'essenziale in biologia , Mondadori, 2008, ISBN 978-88-7192-399-4 .
- Pievani T., La teoria dell'evoluzione. Attualità di una rivoluzione scientifica , Il Mulino, 2006, ISBN 88-15-10975-7 .
- Scott F. Gilbert, Biologia dello sviluppo , Zanichelli, 2005(3 ed.), ISBN 88-08-07275-4 .
Articoli
- Raineri, M., Darwin e la nuova Evo Devo. Percorso storico, nodi irrisolti, punti di vista alternativi [ collegamento interrotto ] , in Emmeciquadro , agosto 2007, pp. 7-17. URL consultato il 26 luglio 2010 .
- CA Redi, Zuccotti, M.; Garagna, S., Evo/Devo: la storia evolutiva dei genomi , in Le Scienze , ottobre 2004.
- ( EN ) A. Abzhanov, Protas, M.; Grant, BR; Grant, PR; Tabin, CJ, Bmp4 and Morphological Variation of Beaks in Darwin's Finches , in Science , vol. 305, n. 5689, 2004, pp. 1462-1465, DOI : 10.1126/science.1098095 . URL consultato il 26 luglio 2010 .
- ( EN ) P Beldade, French, V.; Brakefield, PM, Developmental and genetic mechanisms for evolutionary diversification of serial repeats: eyespot size in Bicyclus anynana butterflies , in Journal of experimental zoology. Part B, Molecular and developmental evolution , 310B, n. 2, 2008, pp. 191-201, DOI : 10.1002/jez.b.21173 .
- ( EN ) PM Brakefield, French, V., Evolutionary developmental biology: how and why to spot fly wings , in Nature , vol. 433, n. 7025, 2005, pp. 466-7, DOI : 10.1038/433466a .
- ( EN ) PM Brakefield, Beldade, P.; Zwaan, BJ, Injection of chemicals into pupae of the African butterfly Bicyclus anynana , in Cold Spring Harbor protocols , n. 5, 2009, DOI : 10.1101/pdb.prot5215 .
- ( EN ) PM Brakefield, Beldade, P.; Zwaan, BJ, Hemolymph extraction from various developmental stages of the African butterfly Bicyclus anynana , in Cold Spring Harbor protocols , n. 5, 2009, DOI : 10.1101/pdb.prot5214 .
- ( EN ) Laubichler, M., A Constrained View of Evo-Devo's Roots , in Science , vol. 309, 2005, pp. 1019-1020, DOI : 10.1126/science.1112990 .
- ( EN ) STE Lommen, Saenko, SV, Tomoyasu, Y.; Brakefield, PM, Development of a wingless morph in the ladybird beetle, Adalia bipunctata , in Evolution & development , vol. 11, n. 3, 2009, pp. 278-89, DOI : 10.1111/j.1525-142X.2009.00330.x .
- ( EN ) Pennisi, E., Evolution of Developmental Diversity Meeting: RNAi Takes Evo-Devo World by Storm , in Science , vol. 304, n. 5669, 2004, p. 384, DOI : 10.1126/science.304.5669.384 . URL consultato il 26 luglio 2010 .
- ( EN ) SV Saenko, French, V.; Brakefield, PM; Beldade, P., Conserved developmental processes and the formation of evolutionary novelties: examples from butterfly wings ( PDF ), in Philosophical transactions of the Royal Society of London. Series B, Biological sciences , vol. 363, n. 1496, 2008, pp. 1549-55, DOI : 10.1098/rstb.2007.2245 . URL consultato il 26 luglio 2010 .
Voci correlate
Collegamenti esterni
- Sito di EvoDevo journal , su evodevojournal.com .
- "EVO-DEVO" - testo tratto dal libro "Chi ha paura di Darwin" (citato in bibliografia)
- Video conferenza di Sean Carroll sull'evoluzionismo. [ collegamento interrotto ] , su pikaia.eu . URL consultato il 7 luglio 2010 .
| Controllo di autorità | GND ( DE ) 7663051-1 |
|---|