Cerebel
| Cerebel | |
|---|---|
 | |
 | |
| Anatomia lui Gray | (RO) Pagina 788 |
| Sistem | Nevrasse |
| Artera | Arterele vertebrale |
| Venă | venele cerebeloase superioare și venele cerebeloase inferioare |
| Identificatori | |
| Plasă | A08.186.211.132.810.428.200 |
| TA | A14.1.07.001 |
| FMA | 67944 |
| ID NeuroLex | birnlex_1489 |
Cerebelul (în latină cerebellum care înseamnă „creierul mic”) este o parte a sistemului nervos central și mai precis a creierului posterior și este prezent la toate vertebratele . Deși, de obicei, este mai mic decât creierul , iar la unele animale precum peștii Mormyridae capătă și mai mare. [1] În ciuda creierului constituie aproximativ 10% din masa cerebrală din „ homo sapiens” , acesta conține aproximativ 80% din numărul total de neuroni din creier [2] , care este de aproximativ 70 miliarde de neuroni (69,03 ± 6,65) comparativ cu 16 miliarde în Coaja (16:34 ± 2,17) și 0,69 ± 0,12 miliarde de trunchi cerebral .
Cerebelul joacă un rol important în controlul mișcării. De asemenea, este implicat în funcții cognitive , cum ar fi „ atenția , memoria și limbajul , precum și în reglarea răspunsurilor la frică sau plăcere [3]. Cu toate acestea, funcțiile sale motorii sunt cele mai fiabile și studiate din punct de vedere istoric, deși este se estimează că doar 20% din funcțiile cerebeloase sunt de natură motorie [4] . Lobii cerebeloși sunt de fapt implicați în întregul spectru al domeniilor neuropsihologice [5] [6] . În sine, cerebelul uman nu inițiază mișcarea, ci contribuie la coordonarea, precizia și sincronizarea exactă a acesteia [7] : primește intrarea din sistemele senzoriale ale măduvei spinării și din alte zone ale creierului și integrează aceste semnale în optimizarea activitatea motorie. [8] O afectare cerebelară la om produce deficite în ordinea mișcării, în „ echilibru , postură și învățare motorie și, în plus, o varietate de deficiențe încărcate de limbajul domeniului, atenția și memoria sunt bine documentate în Neuropsihologie clinică și în cercetare în neuroștiințe [9] [10] [11] [12] [13] [8] [14] .
Anatomic, cerebelul uman este prezentat ca un antrenament împărțit aproximativ ovoid în două emisfere (emisfere cerebeloase), cântărind aproximativ 130-140 grame [14] și este dispus în fosa craniană posterioară ascunsă sub emisferele cerebrale. Suprafața sa corticală este acoperită de caneluri paralele fin distanțate, spre deosebire de circumvoluțiile neregulate tipice ale cortexului cerebral . Aceste caneluri paralele provin din faptul că cortexul cerebelos este un strat subțire de țesut strâns pliat ca un acordeon. În cadrul acestui strat subțire, există mai multe tipuri de neuroni cu un aranjament foarte regulat, cele mai importante fiind celulele Purkinje și celulele granulare. Această organizare neuronală complexă permite o capacitate mare de procesare a semnalului, dar aproape toată prelucrarea cortexului cerebelos trece printr-o serie de mici nuclee profunde care se află în substanța albă din cerebel. [15]
În plus față de rolul său direct în controlul mișcării, cerebelul este necesar pentru diferite tipuri de învățare motorie, în special în adaptarea la schimbările din relațiile senzorimotorii. Au fost dezvoltate mai multe modele teoretice pentru a explica calibrarea senzomotrică în termeni de plasticitate sinaptică în cerebel. Aceste modele sunt derivate din cele formulate de David Marr și James Albus , pe baza observației că fiecare celulă Purbellje cerebelară primește două tipuri de intrare total diferite: una include mii de intrări slabe din fibrele paralele ale celulelor granulare; cealaltă este o intrare extrem de puternică dintr-o singură fibră de cățărare . [16] Conceptul de bază al teoriei Marr-Albus este că fibra rampiacantă acționează ca un „semnal de predare”, ceea ce induce o schimbare durabilă a puterii intrărilor în fibra paralelă. Observațiile depresiei pe termen lung în intrările din fibrele paralele au oferit suport pentru aceste teorii, deși validitatea lor rămâne încă controversată. [16]
Rapoarte și configurație externă

În ansamblu, cerebelul pare a fi alcătuit dintr-o parte centrală, numită vierme, și două expansiuni laterale convexe, emisferele cerebeloase. Suprafața din jurul cerebelului, la fel ca telencefalul , prezintă numeroase brazde la o tendință predominant transversă care delimitează teritorii specifice, lobii și lobulii. În general are fața superioară, fața inferioară și circumferința.
Fața superioară este acoperită de lobii occipitali de care este separată de tentoriul cerebelului, un sept al durei mater care este dispus orizontal. Această lamă care se inserează pe scara „ osului occipital ”, ajută la delimitarea sinusului venos și continuă înainte pentru a forma un spațiu oval mare care delimitează inferior porțiunea supratentorială a creierului . Pe linia mediană, fața superioară se ridică în relief, fața superioară a viermelui, formată de viermele însuși.
Fața inferioară este în relație cu spatele cranian și cu cisterna magna (sau cerebelos-medular), un rezervor mare care conține lichid cefalorahidian . Acest rezervor este în comunicație cu cavitatea celui de-al patrulea ventricul prin trei găuri: una centrală, foramenul lui Magendie și două foramine laterale ale lui Luschka. În partea centrală a feței inferioare se află valecula cerebelului (Reil's), o depresiune care găzduiește un alt sept al durei mater, secera cerebeloasă.

Circumferința este acoperită în față de o depresiune, hilul cerebelos, delimitat de trei pedunculi cerebeloși direcți către trunchiul cerebral . Hilul este delimitat în sus de valva Vieussens și de valva Tarin care se alătură hilului formând un unghi acut, fastigium, deschis în față și formând vârful acoperișului celui de-al patrulea ventricul. Valva Vieussens, în special, este o folie care leagă centrul medular de tecti lamina (folie cvadrigeminală) și este formată din două straturi: un vârf al substanței albe , velul medular superior (sau frontal), care ocupă spațiul între pedunculii superiori și un strat mai profund de substanță cenușie care continuă cu fața inferioară a viermelui. Valva Tarin sau velul medular inferior (sau posterior) este un ependim al laminei, continuarea membranei tectoriale, care ocupă spațiul dintre pedunculii inferiori.
Unghiul cerebelos
Unghiul cerebelos este o zonă triunghiulară delimitată posterior și dorsal de cerebel și pod , anterior și lateral de osul temporal petros. Este plasat în fața flocului. În această zonă sunt concentrate nucleele nervilor cerebrali faciale , intermediare și vestibulare-cohleare care apar învelite de o teacă meningeală comună. Importanța acestei zone este pur clinică și datorită relațiilor intime și delicate care există între elementele prezente; alterarea, de fapt, care afectează această zonă prezintă un renumit cadru anatomoclinico sub formă de sindrom unghiular ponto-cerebelos .
Pedunculii cerebelosi
Pedunculii sunt căi de legătură între cerebel și trunchiul cerebral (și, prin urmare, cu întregul sistem nervos central) alcătuit din mănunchiuri de fibre nervoase atât aferente, cât și eferente.
- Pedunculii cerebeloși inferiori sau corpurile restiformi, se extind de la hilul cerebelului până la jumătatea superioară a bulbului și conțin fibre predominant aferente la cerebel de la bulb și măduva spinării .
- Pedunculele cerebeloase medii sau brahia pontis se extind de la hilul cerebelului până la trunchiul cerebral al podului și sunt cele mai voluminoase. Mai delimitat caudal, împreună cu fața frontală a flocului și pedunculii cerebeloși inferiori, colțul cerebral, o zonă din care ies nervii cranieni VII, intermediarii VIII și înfășurați de o teacă meningeală comună.
- Pedunculii cerebeloși superiori sau brahia conjunctivă care se extind de la hilul cerebelului la tegmentul creierului mediu , sunt de formă turtită și conectați între ei prin valva Vieussens.
Subdiviziune lobara
Atât viermele, care emisferele sunt împărțite de cinci brazde adânci, fisuri sau fisuri în lobii și lobulii principali. Fisurile secundare împart apoi fiecare lobul în lamine.
Lobi

Cerebelul este împărțit de fisurile posterolaterale și primare în 3 lobi, fiecare constând dintr-o porțiune centrală în vierme și două porțiuni laterale în emisfere:
- lobul anterior este constituit din lingula, din lobulul central și din culmen și aripile lor și anume aripa lingulei, aripa lobulului central și lobul quandrangolare din față. Pe fața superioară este delimitată postero-inferior de fisura primară care este purtată antero-lateral întâlnind fisura orizontală. Pe fața inferioară este delimitată de marginea superioară a pedunculilor cerebeloși mijlocii.
- lobul posterior este constituit din lobulul simplu, din folium vermis, din tubercul, din piramida, dall'uvula și aripile lor sau lobul pătrangular posterior, lobulii semilunari superiori și inferiori, lobul biventr și amigdalele cerebeloase. Pe fața superioară este delimitată anterior de sulcul primar și de fisura orizontală, pe fața inferioară de fisura posterolaterală care o separă de lobul flocculonodular. Amigdalele sunt două structuri caracteristice în formă triunghiulară, cu vârful orientat spre inferior-medial care constituie aripile uvulei; au tendința de a acoperi ușor viermele cerebelos pe aspectul inferior al cerebelului.
- lobul flocculonodulare este constituit din nodul și aripa sa, flocul. Se află între vălul medular inferior și fisura posterolaterală.
Lobule
Celelalte fisuri împart apoi lobul anterior și posterior în lobuli suplimentari. Clasificarea Reil [17] prevede o împărțire în lobuli ai vermisului și lobulilor emisferelor. Pentru fiecare lobul al viermelui, există doi lobuli ai emisferelor, unul pe fiecare parte.
Lobulii viermelui
Fața superioară
- Lobulul lingulei viermelui
- Lobulul central
- Munticulus cu culmen si inclinati
- Folium vermis
Fața inferioară
- Tubercul vermis în față și în spate
- Piramidă
- Uvula sau uvula
- Nodul vermian
Lobulii emisferelor
Fața superioară
- Frenule ale lingulei
- Aripile lobulului central
- Lobi patrulateri superiori și inferiori
- Lobi semilunari superiori
Fața inferioară
- Lobulii semilunari inferiori și lobulul gracilis
- Lobuli biventrici
- Amigdalele
- Flocculi
Crăpături

Cerebelul are numeroase fisuri (numite și fisuri sau brazde), împărțite în principale (care dau viață lobilor) și secundare (responsabile de formarea lobulilor). Pe fața superioară fisurile devin progresiv mai lungi, de la fisura precentrală la fisura orizontală, apoi pe fața inferioară scad la fisura posterolaterală. Pentru a evita repetarea, fiecare fantă împarte atât o porțiune de vierme de următoarea, cât și aripa corespunzătoare asociată cu viermele de următoarea.
Există doar două fisuri principale:
- fisura primară: este cea mai adâncă, formează o curbă antero-laterală și împarte lobul anterior și lobul posterior de culmen de lobul simplu
- fisură postero-laterală: împarte lobul posterior de lobul flocculonodulare și uvula de nodul
Fisurile secundare sunt:
- Fisura precentrală este cea mai scurtă din fața superioară a cerebelului, împarte lingula de lobulul central.
- Fisura preculminare împarte lobulul central de culmen.
- Sulcul postero-superior împarte lobulul simplu de folium vermis.
- Fisura orizontală este cea mai lungă dintre toate, se întinde de la pedunculul cerebelos mijlociu la valecula și împarte folium vermis de tubercul.
- Fisura lunogracilă începe pe suprafața tuberculului.
- Fisura prebiventrale împarte tuberculul de piramidă.
- Fisura intrabiventrală rulează pe suprafața piramidei.
- Fisura secundară împarte piramida dall'uvula.
Structura și configurația internă
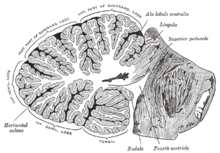

Cerebelul are un strat de substanță cenușie superficială, numit cortex cerebelos, de formă neregulată datorită numeroaselor brazde la tendință predominant transversă. La adâncurile cortexului se află substanța albă care formează, în centru, corpul medular din care se ramifică ramuri de substanță care urmează pliurile griului superficial și formează „arbor vitae”. În adâncimea corpului medular sunt localizate patru nuclee (numite nuclei intrinseci): nucleul acoperișului (sau fastigium), nucleul globos, nucleul emboliform și nucleul dentat.
Cortexul cerebelos
Cortexul cerebelos are o suprafață egală cu jumătate din creier , dar conține un număr de neuroni crescut, deși greutatea sa este de zece ori mai mică [18] . Cu toate acestea, tridimensional, cortexul cerebelos are o structură microscopică uniformă în toate părțile sale, spre deosebire de cortexul cerebral mai complex și mai variabil [18] . Celulele cortexului cerebelos sunt orientate caracteristic într-un mod riguros geometric în raport cu suprafața și axele lamelei lor.
De asemenea, are vase de sânge , celule ale neurogliei și 6 tipuri neuronale [19] :
- granule,
- celule coș,
- celule stea,
- Celulele Golgi,
- celule fus,
- Celulele Purkinje.
Straturi cerebelare

Cortexul cerebelos este format din trei straturi care, de la cel mai superficial la cel mai profund, sunt:
- stratul molecular,
- stratul celular Purkinje,
- stratul de granule.
Stratul molecular
Stratul molecular, adesea de 0,3-0,4mm [18] [20] , este constituit din arborizarea dendritică groasă a celulelor Purkinje și Golgi, din fibre paralele și din fibre urcătoare , în timp ce se dovedește a fi săracă în corpurile celulare. : constau numai dintr- un coș de celule , celulele stelate și celulele fusului.
Arborizarea dendritică a celulelor Purkinje se extinde spre regiunea corticală și este aplatizată datorită dezvoltării într-o direcție transversală față de lungimea lamelelor cerebrale, spre deosebire de celulele Golgi care își extind ramurile atât în direcție transversală cât și longitudinală, mergând să invadeze și o parte din teritoriul ocupat de celulele Purkinje.
Fibrele paralele apar din bifurcația în T a numeroșilor axoni ai granulelor care, odată urcați în stratul molecular, diverg, dând viață fibrelor în sine. Fibrele cățărătoare, pe de altă parte, par a fi terminațiile fibrelor măslin-cerebrale. Ambele fibre se întâlnesc și derivă sinapse cu celule și dendrite prezente în stratul molecular.
În cele din urmă, extensiile celulelor epiteliale ale gliei (ale lui Bergmann) se aranjează radial și, la rândul lor, emit extensii care învelesc toate elementele neuronale prezente, cu excepția sinapselor. Unirea expansiunilor conice ale acestor celule, la nivelul suprafeței cerebelului, constituie o membrană limitativă externă [21] .
Stratul celular Purkinje
Stratul celular Purkinje conține corpurile celulare Purkinje, o parte din arborele lor dendritic și celulele Golgi. Celulele sunt aranjate într-un singur rând.
Strat de granule
Stratul granulelor, mai profund, are o grosime cuprinsă între 0,1 mm și 0,5 mm în fantele de pe partea superioară a lamelelor [20] [21] , conține aproximativ 2,7 milioane pe milimetru cub de neuroni granulari [21] . În acest strat conține corpurile celulare, dendritele bazale și celulele Golgi axonale ; corpurile celulare ale celulelor granulelor mici și partea inițială a axonului lor; ramurile terminale ale fibrelor muscoide (fibre aferente) și o parte a fibrelor cățărătoare care tranzitează către stratul molecular. Din granule mici, provin axonii care se formează fibrele paralele care rulează în stratul molecular.
Celulele cortexului cerebelos
Cerebelul are, pe lângă celulele neurogliei, șase tipuri de celule neuronale [19] : celule Purkinje, celule Golgi, celule granulare, în celule coș, celule stelate și celule fus. Acestea din urmă sunt puține la număr, de tip asociativ cu axon scurt și prezente în întregime în stratul molecular.
Toți sunt neuroni inhibitori care eliberează acid γ-aminobutiric (GABA) în afară de celulele granulare care sunt neuroni excitatori care eliberează L-glutamat .
Celulele Purkinje
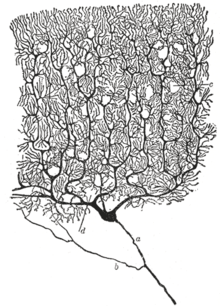
Celulele Purkinje sunt neuroni mari al căror corp alungit este situat în stratul omonim. Acestea sunt aranjate la intervale destul de regulate în interiorul său și au un arbore dendritic dens arborizat care provine de la una sau două dendrite primare și duce până la stratul molecular, unde contractă mii de sinapse prin intermediul numeroaselor spini dendritici cu fibrele paralele ale celule granulare. Arborele dendritic al celulelor Purkinje este limitat în mod caracteristic la o lamă subțire de țesut perpendicular pe axa principală a lamelei. Dendritele celulare Purkinje contractează sinapse prin intermediul unor spini dendritici stubby cu fibre cățărătoare. Ei primesc sinapse inhibitorii de la celulele stelate, celulele coșului și ramurile recurente ale axonilor din alte celule Purkinje. Citoplasma celulelor Purkinje are un reticul endoplasmatic ridat foarte superficial în comparație cu membrana plasmatică și foarte dezvoltat, asociat cu multe mitocondrii și ribozomi. Axonii celulelor Purkinje provin din partea lor bazală și se deplasează spre stratul de granule și apoi penetrează substanța albă. Acestea contractă sinapse assoaxonice cu celule coș. Fiecare axon emite ramuri recurente care formează sinapse cu celule coș (axosomatic), celule Golgi (axosomatic sau axonal) și alte celule Purkinje (axosomatic sau axonal). Restul axonului se acoperă cu teacă de mielină în substanța albă și apoi se ramifică difuz spre nucleii cerebelului sau nucleii vestibulari.
În general, sunt neuroni inhibitori GABAergici a căror funcție este de a modula activitatea nucleilor intrinseci ai cerebelului.
Celulele Golgi
Celulele Golgi au menționat că celulele stelate interioare [20] sau granulele mari [20] sunt neuroni din pirenoforo mare, care se află în zona de suprafață a granulelor stratului, aproape de corpurile celulare ale celulelor Purkinje. Ei posedă un copac dendritic cu o arborizare largă care se extinde semnificativ, spre deosebire de celulele Purkinje, atât în planul longitudinal cât și transversal cu proporții similare. Arborele dendritic se deplasează până la stratul molecular. Arborizarea axonală a celulelor Golgi se extinde în stratul de granule și formează complexe sinaptice numite glomeruli cerebeloși cu fibre de mușchi. Fibrele paralele contractă sinapse cu arborele lor dendritic, dar pot primi și ramuri recurente din celulele Purkinje, fibre urcătoare sau fibre muscoide. Unele dintre celulele Golgi prezintă un'arborizzazione dendritică și în stratul granular, unde devin parte a glomerulilor cerebeloși și primesc sinapse de excitare a fibrelor mușchite. Extensia arborelui dendritic și arborizarea axonală ocupă un volum similar de țesut.
Celulele granulelor
Celulele granulare sunt cele mai numeroase celule ale cerebelului: conform unei estimări, par a fi în număr de 46 miliarde și într-un raport de 3000 cu celulele Purkinje [21] . Sunt neuroni mici, cu corpuri rotunjite din nucleele cu diametrul de 5-8 uM [20] și o citoplasmă bogată în ribozomi și mitocondrii. Se găsesc în stratul de granule care le datorează numele lor. Fiecare emite trei până la cinci dendrite care se termină cu o gheară caracteristică și intră în contact cu grupul de terminații de fibre mușchite care formează glomerulii cerebeloși. Celulele granulare au un singur axon care traversează cortexul cerebral până la stratul molecular unde se ramifică la T, formând fibre paralele. Fibrele paralele se desfășoară paralel cu axa majoră a lamelei cerebrale în care sunt situate pentru câțiva milimetri și sunt împânzite cu structuri globulare (sau umflături) care reprezintă sinapsele cu spini dendritici ai celulelor Purkinje, cu celulele stelate externe, cu celulele coșului și cu cele ale celulelor Golgi. Fiecare arborizare dendtrică poate fi traversată de aproximativ 250.000 de fibre paralele, deși nu se spune că fiecare fibră paralelă se sinapsează pentru a se contracta cu toate dendritele care se întâlnesc [22] . Spre deosebire de alte populații, celulele granulare sunt singurii neuroni excitatori și eliberează L-glutamat în sinapsele lor.
Celule coș
Celulele canistrului posedă un pirenoforo care se află în partea cea mai adâncă a stratului molecular. Sunt interneuroni inhibitori care posedă o arborizare dendritică moderat ramificată pe care unele fibre paralele formează sinapse, este perpendiculară pe axa majoră a lamelei, o geometrie care urmează și dendritele celulelor Purkinje. Celulele coșului primesc, de asemenea, sinapse axosomatice din ramurile recurente ale axonilor celulari Purkinje, fibre urcătoare și fibre muscoide. Axonii celulelor coșului se caracterizează printr-o relație invers proporțională între diametrul lor și distanța față de pirenofor; se găsesc adânc în stratul molecular, unele dintre ramurile lor se desfășoară perpendicular și altele paralele cu axa lamelei în care se află și traversează arborizările a cel puțin o duzină de celule Purkinkje care ating o lungime de câțiva milimetri. Au fost denumite astfel deoarece ramurile colaterale ale axonilor lor flancează dendritele celulelor Purkinje și formează apoi o rețea de fibre de coș în jurul corpurilor și axonului acestor celule.
Celulele stea
Celulele stelate sau celulele stelate externe sunt neuroni mici de formă stelată care se găsesc în stratul molecular al cortexului cerebelos. Ei posedă un'arborizzazione dendritică slab ramificată care se dezvoltă într-un plan perpendicular pe axa lamelelor, în timp ce axonii lor vor merge paralel [20] axa principală a lamelei în care se află. Axonii formează sinapse cu arborele dendritic al celulelor Purkinje. Dendritele celulelor stelate primesc terminații sinaptice de excitare din fibrele paralele ale celulelor granulare.
Celule Lugaro [23]
Sunt neuroni în formă de trabuc situați la același nivel cu celulele Purkinje. Acești neuroni glicinergici inervează celulele stelate și celulele coșului și formează un axon lung orientat transversal care se termină pe celulele Purkinje. Aceștia primesc principalele aferențe dintr-un sistem serotoninergic extracerebelos.
Celule perie unipolare [23]
Acestea sunt celule neuronale de excitare, prezente în principal în regiunile vestibulare ale cerebelului, unde acționează ca un fel de "rapel" pentru aferențele din fibrele muscoide. Fibrele muscoide se termină în sinapse foarte mari la baza („peria”) acestor celule. Axonii lor se termină ca fibre muscoide pe granule.
Fibre aferente

Cortexul cerebelos cuprinde două tipuri de fibre aferente excitatorii, fibrele cățărătoare și fibrele muscoide, distincte în funcție de modul în care se termină în cortexul cerebelos.
În plus față de mușchi și fibre cățărătoare, cortexul cerebelului conține un plex bogat de fibre aminergice din diferite districte ale sistemului nervos central care eliberează neurotransmițători precum norepinefrina, serotonina și dopamina într-un mod paracrin, mai degrabă decât prin sinapse, ceea ce înseamnă că efectele lor se exercită asupra grupurilor de celule și nu asupra unei singure celule. Fibrele serotoninergice provin din formarea reticulară bulbară, cele din locus coeruleus noradrenergic și sunt inhibitoare pentru celulele Purkinje și, în cele din urmă, cele dopaminergice provin din tegmentul ventral.
Fibre cataratoare
Fibrele cățărătoare sunt fibre mielinizate excitante care provin din nucleul olivar inferior și ajung la cerebel prin intermediul fasciculului olivocerebelar. Relația dintre celulele Purkinje și neuronii nucleului olivar inferior este de aproximativ 10: 1, iar apoi fiecare fibră olivocerebelară dă lumină a aproximativ 10 fibre urcătoare [22] . Fibrele se ridică prin substanța albă unde aruncă numeroase colaterale pentru nucleele intrinseci. Sosire în coaja se distribuie către celulele coșurilor și se stelează pentru a se termina apoi în stratul molecular unde se înfășoară în dendritele celulelor Purkinje prin contractarea cu ele a unui număr mare de sinapse (de asemenea, câteva mii [24] ).
Fibrele muscoide
Fibrele cu mușchi sunt fibre mielinizate care provin din fascicule spinocerebelare, trigeminocerebelari, reticolocerebelari și pontocerebelari. Substanța albă emite numeroase ramuri laterale către nucleele intrinseci și odată ajunși în stratul de granule pierde învelișul de mielină pentru a se extinde în clustere sau rozete. Aceste terminații varicoase, împreună cu dendritele granulelor și axonii celulelor Golgi, devin parte a glomerulilor cerebeloși în care șaibele au o locație centrală. Fibrele muscoide nu ajung la stratul molecular, ci se opresc la cel al granulelor, delegând axonilor granulelor transmiterea semnalului până la stratul cel mai exterior. Cu dendritele granulelor formează sinapse glutamatergice, iar receptorii postsinaptici sunt predominant NMDA.
Substanță albă
La sostanza bianca cerebellare, posta profonda alla corteccia, è composta dal corpo midollare e dall' arbor vitae , cioè i tralci di sostanza bianca che partono dal corpo midollare per seguire in profondità le lamine e lamelle rivestite poi dalla corteccia.
Corpo midollare
Il corpo midollare è un massa compatta di sostanza bianca che contiene fibre afferenti per la corteccia, fibre efferenti dalla corteccia (destinate ai nuclei profondi e ai nuclei del tronco encefalico) e fibre di associazione. Il corpo è suddiviso in due lamine che si separano ad angolo acuto delimitando il recesso superiore del IV ventricolo:
- lamina superiore : prosegue nel velo midollare superiore, una formazione di sostanza bianca che forma il tetto della parte pontina del IV ventricolo;
- lamina inferiore : continua nel velo midollare inferiore che contiene tessuto nervoso atrofico e risulta costituito da un ependima raddoppiato dalla membrana tectoria, costituisce il tetto della parte bulbare del IV ventricolo .
Il tronco encefalico è connesso al corpo midollare tramite i tre peduncoli cerebellari. La disposizione ordinata delle fibre nei peduncoli si perde all'interno del corpo stesso, per riformarsi poi a livello delle lamine e delle lamelle.
Nuclei intrinseci

I nuclei intrinseci del cervelletto sono situati nella parte profonda del corpo midollare. Si è deciso di trattarli in un paragrafo a parte per l'importanza che rivestono nelle connessioni cerebellari. Sono quattro paia e, in ordine mediolaterale, sono: nucleo del tetto , nucleo globoso , nucleo emboliforme e nucleo dentato .
Ai nuclei afferiscono sia gli assoni delle cellule di Purkinje sia i collaterali rilasciati dalle fibre rampicanti e muscoidi prima di impegnarsi nella corteccia. Le cellule di Purkinje rilasciano nei nuclei degli impulsi sempre a significato inibitorio con la generazione di una scarica tonica, mentre i collaterali delle altre fibre svolgono una funzione eccitatoria attraverso un'attività elettrica irregolare (burst).
Nucleo del tetto (o del fastigio)
Il nucleo del tetto si trova nella sostanza bianca al di sopra del IV ventricolo ed è separato dallo strato ependimale da un fascio di fibre mieliniche. Dista solo 12mm [24] dal nucleo controlaterale e di fatto è il nucleo più mediale. Presenta una forma ovoidale e le cellule multipolari sono più grandi nei due terzi craniali rispetto alla parte più inferiore. Appartiene funzionalmente al vestibolocerebello.
Nucleo globoso
Il nucleo globoso comprende 2-3 gruppi di cellule multipolari simili al nucleo del tetto. È di dimensioni inferiori rispetto al precedente. Si trova in prossimità dell'ilo del nucleo dentato dal quale è difficilmente individuabile. Insieme al nucleo emboliforme forma quello che viene a volte definito il nucleo interposito . Appartiene funzionalmente allo spinocerebello.
Nucleo emboliforme
Il nucleo emboliforme è situato lateralmente è un po' più caudalmente al nucleo globoso e presenta 2-3 gruppi di cellule di varie dimensioni. Si trova in prossimità dell'ilo del nucleo dentato dal quale è difficilmente individuabile. Insieme al nucleo globoso forma quello che viene a volte definito il nucleo interposito . Appartiene funzionalmente allo spinocerebello.
Nucleo dentato
Il nucleo dentato è il più voluminoso ed è disposto vicino al verme tra il paraflocculo e il peduncolo cerebellare inferiore. È chiamato anche oliva cerebellare per la particolare disposizione simile al nucleo olivare inferiore: ha la forma di una lamina grigia pieghettata con l'apertura volta in direzione dorsale e mediale. Presenta cellule multipolari di varie dimensioni dove, le più grandi e filogeneticamente più antiche prendono una posizione dorsomediale, mentre le più piccole e recenti una ventrolaterale. Appartiene funzionalmente al pontocerebello.
Suddivisione e classificazione dei territori
In base all'origine filogenetico o alla funzione, è possibile suddividere il cervelletto in diversi territori che possono comprendere o meno i diversi lobi e lobuli. Le connessioni del cervelletto e quindi le suddivisioni funzionali sono strettamente collegate alla filogenesi. Nell'evoluzione, compare prima il lobo flocculonodulare che si sviluppa insieme ai nuclei vestibolari spiegando i collegamenti fra archicerebellum e cerebello vestibolare e il motivo per cui le fibre, anziché passare per i nuclei intrinseci nel corpo midollare, si portano direttamente al bulbo.
Suddivisione filogenetica
In base all'origine filogenetica è possibile suddividere il cervelletto in tre porzioni, dalla più antica alla più recente:
- archicerebellum : comprende il nodulo ei flocculi e corrisponde quindi al lobo flocculonodulare, delimitato dalla Scissura Postero-laterale;
- paleocerebellum : compare nella filogenesi insieme alla stazione eretta e comprende le formazioni situate anteriormente alla Scissura Primaria, che nel complesso corrispondono al lobo anteriore, con l'aggiunta della piramide , dell' uvula -o ugola- e delle tonsille ad essa correlate;
- neocerebellum : si è sviluppato in collegamento con la corteccia telencefalica. Il neocerebellum è il territorio più esteso, comprendendo gli emisferi cerebellari.
Suddivisione funzionale
La suddivisione funzionale del cervelletto è stata fatta prendendo in considerazione i territori di afferenza e di efferenza definendo quindi: un vestibolocerebello , uno spinocerebello e un pontocerebello .
Vestibolocerebello (archicerebello)
Il vestibolocerebello o cerebello vestibolare : corrisponde al lobo flocculonodulare ed è quindi completamente sovrapponibile all' archicerebellum . È connesso ai nuclei vestibolari del bulbo e riceve afferenze vestibolari dirette dai canali semicircolari, dagli organi otolitici, dalle paraolive, dalla formazione reticolare e dai nuclei basilari del ponte. Le principali efferenze sono dirette, quindi, ai nuclei vestibolari e costituiscono la prima tappa della via cerebellovestibolospinale . Per queste connessioni il vestibolocerebello è coinvolto nel mantenimento e ripristino dell'equilibrio per la stazione eretta , la deambulazione e di coordinazione del movimento di testa e occhi.
Spinocerebello (paleocerebello)
Lo spinocerebello o cerebello spinale : è posizionato nella parte centrale dei lobi anteriore e posteriore, trovandosi quindi in tutto il verme esclusa la sua zona centrale. Le afferenze sono costituite da tratti della via spinocerebellare , prima di arrivare nel cervelletto passano attraverso il midollo spinale, il tronco encefalico e il nucleo spinale del trigemino. Trasportano informazioni esterocettive e propriocettive che formano due distinte rappresentazioni della periferia nel lobo anteriore e posteriore. Le efferenze sono dirette alla formazione reticolare e ai nuclei vestibolari laterale e inferiore attraverso il nucleo del tetto, mentre attraverso i nuclei emboliformi e globoso raggiungono il nucleo rosso controlaterale e poi il nucleo centrale ventrale del talamo . Da qui poi i fasci raggiungeranno i motoneuroni del midollo spinale e la corteccia motoria. Lo spinocerebello è quindi connesso a due distinte vie: cerebellovestibolospinale e cerebellorubro(reticolo)spinale con le quali agisce sia sui muscoli estensori che flessori.
Pontocerebello (neocerebello)
Il pontocerebello o cerebrocerebello o cerebello cerebrale : è rappresentato dagli emisferi cerebellari e dalla parte centrale del verme. Le fibre afferenti, tramite i nuclei basilari del ponte , arrivano dalla corteccia sensitiva e motoria, dal nucleo olivare superiore, dal midollo spinale e dalla lamina tecti . Le fibre efferenti, attraverso il nucleo dentato, sono dirette verso il nucleo rosso controlaterale da cui poi si dirigono al talamo e alla corteccia motoria e premotoria.
Derivazione embriologica

Il cervelletto si sviluppa dal labbro rombico, un ispessimento della parte dorsale delle lamine alari del metencefalo , che forma il margine craniale del IV ventricolo [25] [26] . Due emisferi rudimentali si formano come rigonfiamenti rotondeggianti proiettati nel ventricolo stesso separati da una parte del tetto metencefalico che andrà a formare il verme. Successivamente si assiste all'estroversione del cervelletto: una riduzione della porzione intraventricolare accompagnata dall'aumento della prominenza extraventricolare. Tale sviluppo è dovuto alla migrazione verso l'esterno delle cellule presenti nel labbro rombico per formare uno strato sotto la glia limitante sopra la superficie del cervelletto in evoluzione, lo strato germinativo esterno [25] .
Si forma poi la prima fessura, che corrisponde alla parte più laterale della fessura posterolaterale. Alla fine del terzo mese compare un solco trasversale sulla superficie superiore del cervelletto primitivo formando la prima fessura che divide il verme ed entrambi gli emisferi [25] .
Avviene quindi l'espansione degli emisferi che, accrescendosi in misura maggiore al verme, lo vanno a ricoprire segregandolo nel solco profondo della vallecola. Nascono poi numerosi solchi trasversi [25] .
Alcuni neuroblasti della zona intermedia delle lamine alari migrano nella zona marginale e si differenziano nei neuroni della corteccia cerebellare. Altri neuroblasti, invece, danno origine ai nuclei intrinseci [26] .
Trasmissione degli impulsi
Le fibre afferenti parallele e rampicanti veicolano sempre un segnale di tipo eccitatorio, mentre le cellule stellate e dei canestri contraggono sinapsi inibitorie. Le fibre muscoidi che terminano nella corteccia formano i glomeruli cerebellari dove l'impulso che trasmettono alle cellule dei granuli viene modulato dalle sinapsi inibitorie degli assoni delle cellule del Golgi. Le fibre efferenti della corteccia, costituite dagli assoni delle cellule di Purkinje, sono sempre inibitorie.
Afferenze
Le fibre afferenti al cervelletto, qualsiasi sia la loro origine e la via a cui appartengono, sono solo di due tipi: muscoidi e rampicanti. Le fibre entrano nel cervelletto attraverso i peduncoli cerebellari e, passando nella sostanza bianca, gettano collaterali nei nuclei intrinseci per terminare poi nella corteccia in modo del tutto diverso.
Le fibre muscoidi si fermano nello strato dei granuli e terminano formando i glomeruli cerebellari dove contraggono sinapsi di tipo eccitatorio con le cellule dei granuli. In tale sede terminano anche gli assoni delle cellule del Golgi. Da qui, l'impulso segue gli assoni delle cellule dei granuli che ascende in modo rettilineo nello strato molecolare finché, una volta raggiunto, non si biforcano a T dando vita alle fibre parallele che decorrono parallelamente all'asse maggiore delle lamelle (perpendicolarmente agli alberi dendritici delle cellule di Purkinje) e contraggono sinapsi eccitatorie con tutti i dendriti che incontrano (sia sulle cellule di Purkinje, che in quelle a canestro o stellate).
Le fibre rampicanti, invece, si dirigono direttamente nello strato molecolare della corteccia e stabiliscono sinapsi eccitatorie con i dentriti delle cellule di Purkinje.
Circuiti della corteccia

Le cellule di Golgi, presenti nello strato granulare, ricevono anch'esse sinapsi eccitatorie dalle fibre muscoidi e parallele che traducono poi in un segnale inibitorio per i granuli.
Le cellule stellate e dei canestri, presenti nello strato molecolare, presentano una s parallela a quella delle cellule di Purkinje. L' assone delle cellule dei canestri, in particolare, si dirige lungo un piano perpendicolare all'asse della lamella portandosi a due cellule di Purkinje laterali a quella su cui si ramifica. Le fibre parallele che arrivano sulle cellule dei canestri, portano un segnale di tipo eccitatorio che si ripercuote in un segnale inibitorio verso cellule di Purkinje poste in file più caudali e craniali, producendo un'inibizione laterale che aumenta il contrasto tra le file di tali cellule.
Efferenze
Gli impulsi corticifughi sono di tipo inibitorio e sono tutti dati dall'integrazione o dalla somma degli impulsi scaricati sulle cellule di Purkinje. Gli assoni delle cellule di Purkinje, lasciano la sostanza grigia per terminare nei nuclei intrinseci della sostanza bianca dove svolgono un'ulteriore attività inibitoria che contrasta l'azione eccitatoria dei collaterali delle fibre muscoidi e rampicanti. Un ulteriore controllo sull'impulso avviene anche sulle e dalle cellule di Purkinje stesse: il ramo collaterale ricorrente, che si stacca dal loro assone poco dopo la sua nascita, si dirige verso i denditri con lo scopo di ridurre l'attività efferente con la sua funzione inibitoria sulla cellula stessa.
Infine, le fibre efferenti del cervelletto nascono dai nuclei intrinseci dove avvengono le modulazioni inibitorie finali dopo il passaggio dell'impulso nella corteccia.
Glomerulo cerebellare
Le fibre muscoidi, terminando espandendosi in rosette sui dendriti delle cellule dei granuli, formano i glomeruli cerebellari . Tali glomeruli sono costituiti dall'espansione terminale di una fibra muscoide, 4-5 dendriti delle cellule dei granuli e dall'assone di cellule del Golgi. Nei glomeruli avviene la modulazione della trasmissione degli impulsi trasportati dalle fibre muscoidi ai granuli che poi passano alle cellule di Purkinje. L'impulso di tipo eccitatorio viene, infatti, controllato in senso inibitorio dall'assone della cellule del Golgi che, funzionando come una griglia, seleziona l'entità delle afferenze che devono raggiungere la corteccia.
Connessioni afferenti
Le afferenze al cervelletto sono rappresentate dalle fibre muscoidi che terminano ed attivano le cellule dei granuli, le cellule del Golgi ei nuclei cerebellari e dalle fibre rampicanti provenienti dall'oliva inferiore che invece formano sinapsi con i nuclei cerebellari, con le cellule a canestro, con le cellule del Golgi e controlateralmente sulle cellule di Purkinje. Le fibre che si portano al cervelletto sono spinocerebellari, trigeminocerebellari, vestibolocerebellari, reticolocerebellari, pontocerebellari e olivocerebellari.
Fibre spinocerebellari
Le fibre spinocerebellari sono divise in posteriori, anteriori e rostrali [27] . Sono tutte vie eccitatorie e prima di entrare nella corteccia, emettono collaterali per i nuclei del tetto, globoso ed emboliforme.
Fascio spinocerebellare posteriore
Il fascio spinocerebellare posteriore origina dalla colonna di Clarke, alla base del corno posteriore dei neuromeri toracici, risale omolateralmente, entra attraverso il peduncolo cerebellare inferiore e termina poi omolateralmente nel verme e nelle regioni adiacenti del lobo anteriore e nella piramide e nelle regioni adiacenti del lobo posteriore [28] . Viste le afferenze della colonna di Clarke [29] questo fascio trasporta le informazioni propriocettive ed esterocettive provenienti dagli arti inferiori omolaterali. Una caratteristica di questo fascio è quella di avere gli assoni con il diametro maggiore di tutto il sistema nervoso centrale che, con i suoi 20 µm, permette una maggiore velocità di conduzione [28] .
Fascio spinocerebellare anteriore
Il fascio spinocerebellare anteriore origina a livello lombosacrale nella sostanza grigia intermedia del midollo spinale, decussa subito dopo l'origine e risale sino al mesencefalo per poi scendere ed entrare nel cervelletto attraverso il peduncolo cerebellare superiore. Nella sostanza bianca del cervelletto le fibre di questo fascio decussano una seconda volta, staccano terminazioni per i nuclei cerebellari e poi si portano alla corteccia cerebellare. Grazie ai due incroci, la distribuzione delle fibre ai nuclei e alla corteccia ricalca quella delle fibre del fascio posteriore [28] . Le sue afferenze sono costituite da informazioni relative agli archi riflessi degli arti inferiori e della parte inferiore del tronco.
Fascio spinocerebellare rostrale
Il fascio spinocerebellare rostrale origina dalla zona intermedia dei corni posteriori della sostanza grigia cervicale nel midollo spinale, risale omolateralmente lungo il midollo e il tronco encefalico per poi entrare nel cervelletto attraverso il peduncolo cerebellare inferiore dove si distribuisce a parti della corteccia simili a quelle del fascio cuneocerebellare. Trasporta informazioni relative agli archi riflessi degli arti superiori e del tronco superiore.
Fascio cuneocerebellare
Il fascio cuneocerebellare origina dal nucleo cuneato e dal nucleo cuneato accessorio, segue il fascicolo cuneato risalendo omolateralmente per poi entrare nel cervelletto. Qui termina nella parte posteriore del lobo anteriore, con le fibre propriocettive che si distribuiscono all'apice delle lamelle e le esterocettive alla loro base. Le regioni in cui termina sono il lobulo centrale, il culmen, il lobulo semplice, l'ala del lobo centrale, il lobulo quadrangolare anteriore e posteriore, il tuber, la piramide, l'uvula, il lobulo semilunare inferiore e parte del lobulo biventre. Trasporta informazioni relative a muscoli, articolazioni e neuroni sensitivi cutanei dell'arto superiore.
Fibre trigeminocerebellari
Le fibre trigeminocerebellari originano dai nuclei del trigemino nel tronco encefalico e si portano omolateralmente al cervelletto presso il lobulo semplice e il lobulo gracile, ma anche all'oliva inferiore omolaterale, che poi a sua volta proietta alla corteccia del cervelletto. Trasportano informazioni propriocettive e interocettive della testa.
Fibre olivocerebellari
Le fibre rampicanti che entrano nel cervelletto originano solamente da nuclei appartenenti al complesso olivare inferiore: da qui nascono per terminare controlateralmente su zone (costituite da strisce longitudinali) di cellule di Purkinje nella corteccia cerebellare. I collaterali delle fibre rampicanti si staccano per i nuclei cerebellari o vestibolari che contraggono sinapsi con le cellule di Purkinje raggiunte dalle rampicanti stesse. In tal modo le fibre olivocerebellari possono controllare i segnali in uscita dal cervelletto [28] . In base alla suddivisione del complesso olivare inferiore si hanno due diverse origini per le fibre rampicanti:
- Nucleo olivare principale : da qui nascono le fibre olivari che si dirigono alla corteccia cerebellare emettendo collaterali per i nuclei vestibolare laterale e cerebellari.
- Nucleo olivare accessorio : proiettano al verme e agli emisferi adiacenti.
Fibre vestibolocerebellari
Le fibre vestibolocerebellari primarie originano dal nervo vestibolare, ramo del nervo vestibolococleare , attraversano il corpo iuxtarestiforme e il nucleo vestibolare superiore per poi distribuirsi omolateralmente allo strato dei granuli della parte inferiore dell'uvula, del nodulo, dalla parte ventrale del lobo anteriore e del fondo delle fessure profonde del verme nella corteccia cerebellare [30] .
Le fibre vestibolocerebellari secondarie originano dal nucleo vestibolare superiore, dal mediale e dall'inferiore, si portano bilateralmente nella corteccia cerebellare presso le stesse regioni delle fibre vestibolocerebellari primarie e in più anche al flocculo e all'adiacente paraflocculo ventrale (il paraflocculo accessorio del cervelletto umano), che non ricevono proiezioni vestibolocerebellari primarie.
Fibre reticolocerebellari
Le fibre reticolocerebellari originano dal nucleo reticolare laterale nel bulbo e dai nuclei reticolari paramediani e tegmentali del ponte.
- Nucleo reticolare laterale : proietta ad entrambi i lati del verme e degli emisferi. Le fibre provenienti dalla parte dorsale del nucleo si portano omolateralmente al corrispondente emisfero cerebellare, quelle della parte ventrale del nucleo si distribuiscono bilateralmente al verme; alcuni rami si portano al nucleo del fastigio, al nucleo emboliforme e ad una parte del nucleo globoso.
- Nucleo reticolare paramediano : proietta diffusamente a tutto il cervelletto.
- Nucleo reticolare tegmentale : proietta al cervelletto tramite il peduncolo cerebellare medio e le sue fibre terminano nel lobo anteriore, lobulo semplice, folium vermis, tuber, al nodulo e al flocculo; rami proiettano al nucleo del fastigio, al nucleo globoso e al nucleo dentato.
Fibre pontocerebellari
Le fibre pontocerebellari originano dai nuclei pontini ed entrano nel cervelletto attraverso il peduncolo cerebellare medio di cui costituiscono quasi tutte le afferenze.
- Nuclei pontini dorsali : ricevono afferenze dalle aree visive della corteccia e trasportano informazioni sul controllo del movimento proiettandole al lobulo semplice, lobulo quadrangolare anteriore, folium vermis e tuber.
- Nuclei reticolari pontini : proiettano a tutte le aree del cervelletto fatta eccezione per il nodulo e la lingula, alcuni collaterali giungono al nucleo dentato.
Connessioni efferenti
È possibile dividere le connessioni efferenti del cervelletto nelle proiezioni inibitorie della corteccia (e nella fattispecie delle cellule di Purkinje) dirette ai nuclei cerebellari e vestibolari e nelle proiezioni efferenti dirette dai nuclei cerebellari ai centri motori di tronco encefalico e corteccia cerebrale motoria.
Le principali efferenze del cervelletto sono le fibre corticonucleari, corticovestibolari, cerebellovestibolari, cerebelloreticolari, cerebellorubre e cerebellotalamiche.
Efferenze dalla corteccia
Il verme cerebellare proietta principalmente ai nuclei vestibolari e al nucleo del fastigio, mentre gli emisferi cerebellari proiettano al nucleo dentato, al nucleo emboliforme, ai nuclei globosi ea varie parti della formazione reticolare.
Zone della corteccia

La corteccia cerebellare è suddivisia in zone in base alla destinazione delle fibre che vi nascono [31] . Questa suddivisione consiste in strisce ( zone ) di cellule di Purkinje non individuabili nelle sezioni istologiche convenzionali. Il verme del lobo anteriore e il lobulo semplice sono divisi in:
- zona A: la striscia più mediale del verme; proietta al polo craniale del nucleo del tetto e riceve le fibre rampicanti dalla regione caudale del nucleo olivare accessorio mediale;
- zona X: proietta ai gruppi di cellule interstiziali, che si trovano tra il nucleo del fastigio e il nucleo interposito posteriore, e riceve le fibre rampicanti dalla regione intermedia del nucleo olivare accessorio mediale;
- zona B: la striscia laterale del verme; proietta al nucleo vestibolare laterale e riceve fibre rampicanti dalla parte caudale del nucleo olivare accessorio dorsale.
Proseguendo verso gli emisferi, nella regione intermedia, sono presenti le zone C 1 , C 3 , Y. Queste zone proiettano al nucleo interposito anteriore e ricevono fibre rampicanti dalla regione craniale del nucleo olivare accessorio dorsale.
Il resto degli emisferi è costituito da:
- zona C 2 : proietta al nucleo interposito posteriore e riceve le fibre afferenti rampicanti dalla regione craniale del nucleo olivare accessorio mediale;
- zona D (D 1 e D 2 ): proietta al nucleo dentato caudale e rostrale e riceve le fibre rampicanti dalle lamine ventrali e dorsali del nucleo olivare principale.
Le principali connessioni delle zone C 2 e D sono con la corteccia cerebrale. Esse sono infatti connesse ad aree corticali motorie, premotorie e parietali con suddivisione somatotopica. [32]
Le altre parti del cervelletto presentano una suddivisione zonale di minore importanza o ancora poco chiara, rimanendo luogo di numerose efferenze. Il flocculo, invece, non presenta zone e le sue proiezioni sono dirette ai nuclei vestibolari superiore, mediale ed inferiore.
Fibre corticonucleari
Le ' fibre corticonucleari originano dalle cellule di Purkinje del verme del lobulo anteriore del cervelletto, in un'area detta zona A, che comprende la porzione mediale del verme e di cui fanno parte tutte le divisioni del verme tranne il nodulo. Queste fibre proiettano al polo craniale del nucleo del fastigio. Il folium vermis e il tuber, due regioni del verme che ricevono afferenze visive, proiettano anch'essi al nucleo del fastigio, ma al suo polo caudale e sono coinvolti nei movimenti saccadici dell'occhio. Le zone C1 e C3, laterali alla zona B e comprendenti cellule di Purkinje degli emisferi cerebellari proiettano entrambe ai nuclei emboliformi, mentre la zona C2, tra essi compresa, proietta al nucleo globoso. La parte restante degli emisferi, la più laterale, proietta al nucleo dentato. Le fibre corticonucleari sono in parte efferenze del paleocervelletto (quelle che originano dalle zone B, C1, C2 e C3) e in parte del neocervelletto (tutto quanto proietta al nucleo dentato).
Fibre corticovestibolari
Le fibre corticovestibolari originano dalle cellule di Purkinje del verme del lobulo anteriore del cervelletto, precisamente da una stretta striscia longitudinale (dunque perpendicolare alle lamelle) appena laterale alla zona A, detta zona B che comprende lobulo centrale, culmen e soprattutto il lobulo semplice. I loro assoni proiettano al nucleo vestibolare laterale omolaterale, nel ponte. La piramide, l'uvula, il nodulo e il flocculo proiettano direttamente ai nuclei vestibolari superiore, mediale ed inferiore. In generale le fibre corticovestibolari sono efferenze dell'archicervelletto.
Efferenze dai nuclei
Le efferenze dei nuclei intrinseci sono dirette ai centri motori del tronco encefalico e, attraverso il talamo, alla corteccia motoria.
Fibre cerebellovestibolari
Le fibre cerebellovestibolari comprendono le connessioni dei nuclei cerebellari con i nuclei vestibolari. Il nucleo del fastigio è collegato reciprocamente con i nuclei vestibolari, la sua efferenza più importante in questo senso è il fascicolo uncinato che decussa all'interno della sostanza bianca cerebellare, passa posteriormente al peduncolo cerebellare superiore e raggiunge i nuclei vestibolari lateralmente. Dal nucleo del fastigio origina tuttavia anche un contingente di fibre non crociate che raggiunge i nuclei vestibolari (inferiore e mediale) attraverso i peduncoli cerebellari inferiori. Dal nucleo vestibolare laterale origina poi il fascio vestibolospinale laterale, mentre dal nucleo vestibolare mediale il fascio vestibolare mediale. Alcune fibre del nucleo del fastigio non sinaptano nei nuclei vestibolari e procedono inferiormente nel midollo spinale costituendo il fascio fastigiospinale. I nuclei vestibolare mediale ed inferiore proiettano fibre al talamo mediante il fascicolo longitudinale mediale, tali fibre sono implicate nel riflesso vestibolo-oculare.
Fibre cerebelloreticolari
Le fibre cerebelloreticolari comprendono le connessioni dei nuclei cerebellari con la formazione reticolare. Alcune fibre cerebellovestibolari originatesi dal polo craniale nucleo del fastigio attraversano i nuclei vestibolari per poi distribuirsi alla formazione reticolare mediale e sui grandi neuroni del nucleo reticolare mediale. La formazione reticolare a sua volta è connessa al midollo spinale mediante il fascio reticolospinale mediale e il fascio reticolospinale laterale. Il polo caudale del nucleo del fastigio origina fibre che decussano e poi si portano alla formazione reticolare mediana del ponte, esse controllano i movimenti orizzontali dello sguardo.
Fibre cerebellorubre
Le fibre cerebellorubre comprendono le connessioni dei nuclei cerebellari con il nucleo rosso del mesencefalo. Il nucleo emboliforme proietta alla porzione magnocellulare nucleo rosso e questo a sua volta dà origine al fascio rubrospinale che tuttavia nell'uomo è poco sviluppato rispetto ad altri animali. Il nucleo dentato proietta alla porzione parvicellulare del nucleo rosso che a sua volta dà origine al fascio tegmentale centrale che si collega con il nucleo olivare principale.
Fibre cerebellotalamiche
Le fibre cerebellotalamiche comprendono le connessioni dei nuclei cerebellari con il talamo . Il nucleo globoso proietta al nucleo ventrale laterale del talamo che a sua volta è connesso con la corteccia motoria, in particolare alle porzioni da cui originano fibre volte al controllo dei muscoli assiali degli arti. Il nucleo dentato proietta al nucleo ventrale laterale del talamo mediante il fascio dentatotalamico, il nucleo è a sua volta connesso con la corteccia premotoria. Infine anche alcune fibre del nucleo del fastigio giungono al talamo, alcune attraversando prima i nuclei vestibolari del ponte altre, soprattutto originatesi dalla sua porzione caudale, in maniera diretta.
Anatomia funzionale
Le informazioni che arrivano dalla periferia portano i dettagli dei movimenti nel corso della loro esecuzione permettendo al cervelletto di controllare il tono muscolare durante i movimenti stessi e quindi di adattare subito lo stato di contrazione muscolare alle variazioni di carico o delle richieste fisiologiche per quanto concerne l'ampiezza, l'efficacia, la traiettoria e la durata del movimento [33] . Nel fare questo permette, quindi, di ridurre al minimo le contrazioni non coordinate dei muscoli antagonisti che causano piccole oscillazioni durante il movimento, il cosiddetto tremore fisiologico (in questo è implicata la porzione posterolaterale del cervelletto [34] ).
Sia i segnali periferici che centrali vengono confrontati in ogni istante e usati per produrre una risposta efficiente per modulare, ed eventualmente correggere, il segnale sia nel tragitto discendente che ascendente. Qualora sussistano delle differenze tra il movimento programmato e quello effettivamente realizzato, il cervelletto è in grado di correggere, con un meccanismo di feedback negativo , il movimento durante il suo realizzarsi.
Il modulo di funzionamento del cervelletto si ripete invariato nonostante le diverse connessioni e funzioni attribuite. La corteccia è costituita ovunque dagli stessi strati che sono raggiunti dalla stesse fibre afferenti e da cui dipartono l'unico tipo di fibre efferenti dirette ai nuclei intrinseci del cervelletto [35] .
Un danno cerebellare, infine, non impedisce l'utilizzo di una determinata funzione, ma ne riduce l'efficienza.
Apprendimento motorio
Queste capacità del cervelletto si applicano sia durante i processi cognitivi (che coinvolgono le aree corticali limbiche e paralimbiche) sia durante la ripetizione di movimenti complessi (apprendimento motorio). La ripetizione nel tempo di determinati schemi motori, modifica i circuiti cerebellari (effetto plastico) accrescendo l'efficienza di controllo nell'atto motorio e quindi anche l'efficacia con cui viene effettuato. Si parla soprattutto di azione che richiedono schemi elevati di coordinazione (suonare il pianoforte) e in cui l'esecuzione dei singoli atti che compongono il movimento sia indipendente dall'intervento cosciente della corteccia cerebrale [35] .
Nel cervelletto si crea quindi un ben preciso schema motorio che viene trasmesso al complesso olivare inferiore tramite proiezioni collaterali del fascio corticospinale. Anche alcune informazioni cerebellari uscenti attraverso il peduncolo cerebellare superiore che raggiungono il nucleo rosso vengono poi ritrasmesse all'oliva inferiore per il confronto con lo schema motorio originale [34] .
Correzione del movimento
In ogni caso sembra che proprio per questo motivo, il sistema delle fibre rampicanti olivocerebellari sia implicato nella correzione degli errori motori durante la replicazione di uno schema già preimpostato e memorizzato [34] . Le fibre rampicanti, infatti, aumentano la loro scarica di potenziali complessi generando un aumento di Ca 2+ intracellulare molto evidente. Questo, sommatosi all'aumento relativo alla stimolazione delle stesse cellule da parte delle fibre parallele, induce in queste ultime un processo di depressione sinaptica a lungo termine ( long term depression ) causato fondamentalmente da un'endocitosi dei recettori AMPA per il glutammato con conseguente diminuzione di scarica delle cellule del Purkinje sui nuclei. Così il movimento tenderà ad essere corretto tramite le vie di controllo del cervelletto sulla corteccia e sulle vie motorie discendenti.
Mantenimento della stazione eretta
A questo riguardo svolge un ruolo essenziale il verme del cervelletto essendo implicato in quella serie di movimenti muscolari inconsci che permettono di mantenere la stazione eretta mentre si compie un'azione e nell'insieme noti come anticipazione motoria [34] . Nel momento in cui si prende un libro da uno scaffale, per esempio, vengono attivati per prima i muscoli flessori plantari della caviglia e non i flessori della spalla , del gomito o delle dita . Questo permette che la parte anteriore del piede spinga indietro gli arti inferiori e il tronco nel momento in cui si afferra il libro permettendo di mantenere la stabilità. Nel momento in cui solleviamo il libro, poi, i muscoli erettori della colonna vertebrale correggono il peso del libro e del braccio impedendo lo sbilanciamento in avanti di testa e tronco. Nello stesso tempo recettori labirintici informano il cervelletto di ogni variazione dell'equilibrio permettendogli di effettuare una risposta coerente con le intenzioni ei movimenti effettuati [34] .
Tempistica e cooperazione
Un esempio dell'importanza della tempistica con cui devono avvenire i movimenti motori e la coordinazione fra i muscoli si ha nella lettura. Per leggere una pagina di un libro è necessario che ad ogni riga gli occhi che scorrono le lettere nelle righe scattino indietro per iniziare la riga successiva [34] . Errori in questa azione può causare la dislessia [34] .
Patologie
Lesioni

Le lesioni al cervelletto sono tutte accomunate da una perdita della coordinazione del movimento o atassia . Lesioni cerebellari specifiche, naturalmente, portano a particolari tipi di atassia che risulta sempre nello stesso lato della lesione [36] .
Lesioni della linea mediana del verme o del lobulo flocculonodulare portano ad un' atassia del tronco, con caratteristica andatura barcollante tipica dell'ubriaco. Questa scoordinazione si accompagna in genere a movimenti anomali degli occhi ( nistagmo ) [36] .
Lesioni laterali al verme causano un' atassia appendicolare, un'incoordinazione degli arti che si traduce nell'incapacità di svolgere movimenti appropriati alla distanza da raggiungere ( dismetria ) ea tremori intenzionali cioè alterazioni della contrazione muscolare durante l'atto motorio [36] .
Lesioni al lobo anteriore portano ad un aumento abnorme del tono muscolare ( ipertonia ). Tale sintomo è anche indicativo di una lesione corticale con attività dei nuclei profondi intatta: in questo caso si assiste ad un'ipertonia dei muscoli estensori che causa un'iperestensione di testa e collo ( opistotono ) [36] .
Anche l' ipotonia , che consiste nella perdita del tono muscolare, è un'alterazione che può essere causata da lesioni al cervelletto. L'ipotonia riguarda tutti i muscoli quando si ha una perdita di funzione totale del cervelletto, oppure solo dei muscoli antigravitari se riguarda solo il nucleo del tetto [36] .
Sindrome dell'angolo pontocerebellare
Tale sindrome è un quadro anatomoclinico che coinvolge gli elementi presenti nell'angolo cerebellare. Tale patologia è solitamente causata da un tumore che coinvolge la guaina del nervo vestibolococleare che causa disturbi all'udito, dell'equilibrio e alterazioni al nervo facciale . Se si espande, può portare alla compromissione del ponte , del cervelletto e di altri nervi encefalici . Altre cause della sindrome possono essere una compressione dovuta ad emorragie o processi espansivi del tronco encefalico [37] .
Note
- ^ William Hodos, Evolution of Cerebellum , in Encyclopedia of Neuroscience , Springer, 2009, pp. 1240-1243, DOI : 10.1007/978-3-540-29678-2_3124 .
- ^ ( EN ) Frederico AC Azevedo, Ludmila RB Carvalho e Lea T. Grinberg, Equal numbers of neuronal and nonneuronal cells make the human brain an isometrically scaled-up primate brain , in Journal of Comparative Neurology , vol. 513, n. 5, 2009, pp. 532-541, DOI : 10.1002/cne.21974 . URL consultato il 18 novembre 2019 .
- ^ Wolf U, Rapoport MJ, Schweizer TA, Evaluating the affective component of the cerebellar cognitive affective syndrome , in J. Neuropsychiatry Clin. Neurosci. , vol. 21, n. 3, 2009, pp. 245-53, DOI : 10.1176/jnp.2009.21.3.245 , PMID 19776302 .
- ^ ( EN ) Scott Marek, Joshua S. Siegel e Evan M. Gordon, Spatial and Temporal Organization of the Individual Human Cerebellum , in Neuron , vol. 100, n. 4, 21 novembre 2018, pp. 977–993.e7, DOI : 10.1016/j.neuron.2018.10.010 . URL consultato il 18 novembre 2019 .
- ^ ( EN ) Jeremy D. Schmahmann, An Emerging Concept: The Cerebellar Contribution to Higher Function , in Archives of Neurology , vol. 48, n. 11, 1º novembre 1991, p. 1178, DOI : 10.1001/archneur.1991.00530230086029 . URL consultato il 18 novembre 2019 .
- ^ Jeremy D. Schmahmann, <174::aid-hbm3>3.3.co;2-w From movement to thought: Anatomic substrates of the cerebellar contribution to cognitive processing , in Human Brain Mapping , vol. 4, n. 3, 1996, pp. 174-198, DOI : 10.1002/(sici)1097-0193(1996)4:3<174::aid-hbm3>3.3.co;2-w . URL consultato il 18 novembre 2019 .
- ^ ( EN ) Orçun Orkan Özcan, Xiaolu Wang e Francesca Binda, Differential Coding Strategies in Glutamatergic and Gabaergic Neurons in the Medial Cerebellar Nucleus , in The Journal of Neuroscience , 6 novembre 2019, pp. 0806-19, DOI : 10.1523/JNEUROSCI.0806-19.2019 . URL consultato il 18 novembre 2019 .
- ^ a b Fine EJ, Ionita CC, Lohr L, The history of the development of the cerebellar examination , in Semin. Neurol. , vol. 22, n. 4, 2002, pp. 375-84, DOI : 10.1055/s-2002-36759 , PMID 12539058 .
- ^ ( EN ) Jeremy D. Schmahmann, Disorders of the Cerebellum: Ataxia, Dysmetria of Thought, and the Cerebellar Cognitive Affective Syndrome , in The Journal of Neuropsychiatry and Clinical Neurosciences , vol. 16, n. 3, 2004-8, pp. 367-378, DOI : 10.1176/jnp.16.3.367 . URL consultato il 18 novembre 2019 .
- ^ ( EN ) Christian Bellebaum e Irene Daum, Cerebellar involvement in executive control , in The Cerebellum , vol. 6, n. 3, 2007, pp. 184-192, DOI : 10.1080/14734220601169707 . URL consultato il 18 novembre 2019 .
- ^ MG Leggio, Phonological grouping is specifically affected in cerebellar patients: a verbal fluency study , in Journal of Neurology, Neurosurgery & Psychiatry , vol. 69, n. 1, 1º luglio 2000, pp. 102-106, DOI : 10.1136/jnnp.69.1.102 . URL consultato il 18 novembre 2019 .
- ^ ( EN ) Peter Mariën, Kim van Dun e Johanna Van Dormael, Cerebellar induced differential polyglot aphasia: A neurolinguistic and fMRI study , in Brain and Language , vol. 175, 2017-12, pp. 18-28, DOI : 10.1016/j.bandl.2017.09.001 . URL consultato il 18 novembre 2019 .
- ^ J. Schmahmann, The cerebellar cognitive affective syndrome , in Brain , vol. 121, n. 4, 1º aprile 1998, pp. 561-579, DOI : 10.1093/brain/121.4.561 . URL consultato il 18 novembre 2019 .
- ^ a b Da Trattato di Anatomia Umana, volume III , p. 89
- ^ Dale Purves, Neuroscience , a cura di Dale Purves, 5th, Sunderland, Mass., Sinauer, 2011, pp. 417-423, ISBN 978-0-87893-695-3 .
- ^ a b Dale Purves Purves, Neuroscience. , 4th, New York, WH Freeman, 2007, pp. 197 –200, ISBN 0-87893-697-1 .
- ^ Da Trattato di Anatomia Umana, volume III , p. 93
- ^ a b c Da Anatomia del Gray , p. 258
- ^ a b In Anatomia del Gray , p. 258 si parla di 5 tipi di neuroni, ma il Trattato di Anatomia Umana, volume III , p. 96 parla anche di una sesta popolazione, le cellule fusiformi.
- ^ a b c d e f Da Trattato di Anatomia Umana, volume III , p. 96
- ^ a b c d Da Anatomia del Gray , p. 260
- ^ a b Da Anatomia del Gray , p. 261
- ^ a b Henry Gray, Henry Gray e Edward Anthony Spitzka, Anatomy, descriptive and applied, by Henry Gray. , Lea & Febiger,, 1913. URL consultato il 30 dicembre 2018 .
- ^ a b Da Trattato di Anatomia Umana, volume III , p. 100
- ^ a b c d Da Anatomia del Gray , p. 336
- ^ a b Da Lo sviluppo prenatale dell'uomo , p. 397
- ^ L'aggettivo rostrale, in anatomia, si riferisce alla parte più craniale di un organo o sezione anatomica. Usato anche in embriologia per descrivere la direzione di sviluppo, ne sono sinonimi craniale e cefalico .
- ^ a b c d Da Anatomia del Gray , p. 262
- ^ La colonna di Clarke riceve tutte le afferenze primarie dei muscoli e delle articolazioni degli arti inferiori, che la raggiungono tramite il fascicolo gracile, e rami collaterali di neuroni sensitivi cutanei.
- ^ Da Anatomia del Gray , p. 264
- ^ Da Anatomia del Gray , p. 265
- ^ Henry Gray, Henry Gray e Edward Anthony Spitzka, Anatomy, descriptive and applied, by Henry Gray. , Lea & Febiger,, 1913. URL consultato il 2 gennaio 2019 .
- ^ Da Trattato di Anatomia Umana, volume III , p. 94
- ^ a b c d e f g Da Anatomia del Gray , p. 267
- ^ a b Da Trattato di Anatomia Umana, volume III , p. 95
- ^ a b c d e Da Trattato di Anatomia Umana, volume III , p. 102
- ^ Da Trattato di Anatomia Umana, volume III , p. 92
Bibliografia
- Anastasi et al., Trattato di Anatomia Umana, volume III , Milano, Edi.Ermes, 2012, ISBN 88-7051-287-8
- Susan Standring, Anatomia del Gray, volume I , Milano, Elsevier, 2009, ISBN 978-88-214-3132-6
- Keith Moore, TVN Persaud, Lo sviluppo prenatale dell'uomo , Napoli, EdiSES, 2009, ISBN 978-88-7959-348-9
Voci correlate
- Atassia
- Bulbo (encefalo)
- Cervello
- Mesencefalo
- Midollo spinale
- Nucleo dentato
- Ponte di Varolio
- Ventricoli cerebrali
- Vie somatosensoriali
Altri progetti
-
 Wikizionario contiene il lemma di dizionario « cervelletto »
Wikizionario contiene il lemma di dizionario « cervelletto » -
 Wikimedia Commons contiene immagini o altri file su cervelletto
Wikimedia Commons contiene immagini o altri file su cervelletto
Collegamenti esterni
- Cervelletto , su Treccani.it – Enciclopedie on line , Istituto dell'Enciclopedia Italiana .
- ( EN ) Cervelletto , su Enciclopedia Britannica , Encyclopædia Britannica, Inc.
- Cervelletto , in Treccani.it – Enciclopedie on line , Istituto dell'Enciclopedia Italiana.
| Controllo di autorità | Thesaurus BNCF 38526 · LCCN ( EN ) sh85022078 · GND ( DE ) 4125073-4 |
|---|