Țesut conjunctiv

Țesutul conjunctiv ( lat. Textus conexivus ) este unul dintre cele patru tipuri de bază de țesut care alcătuiesc corpul celor mai abundente animale din organism. Acest set de țesuturi a fost întotdeauna definit în mod necorespunzător ca „conectiv”, deoarece este preferabil să se utilizeze definiția „țesuturi trofo-mecanice”: fiecare subspecie a categoriei îndeplinește o funcție trofică și / sau mecanică.
Descriere generala
Este definit ca diferite tipuri de țesuturi care au în comun funcția de a asigura conexiunea, susținerea, protecția, hrana, apărarea imună, transportul fluidelor, rezerva de energie a altor țesuturi ale diferitelor organe și care derivă din țesutul conjunctiv țesut, mezenchimul (care provine în principal din mezoderm ).
Din punct de vedere histologic , țesutul conjunctiv poate fi împărțit în diferite subtipuri, în funcție de prerogativele lor morfologice și funcționale, toate caracterizate prin faptul că sunt alcătuite din celule care nu sunt atașate una de alta, dar dispersate într-o substanță intercelulară mai mult sau mai puțin abundentă sau extracelular matricial format dintr-o componentă amorfă și o componentă fibroasă.
Celulele țesutului conjunctiv
Țesutul conjunctiv are o mare varietate de celule, desemnate să desfășoare diferite activități și în raport cu natura țesutului căruia îi aparțin și poziția pe care și-o asumă în corp. În general, este posibil să se facă o distincție între celulele responsabile de formarea și menținerea matricei ( fibroblaste , condroblaste , osteoblaste, cementoblaste, odontoblaste), celule responsabile de apărarea organismului ( macrofage , mastocite , leucocite ) și celulele responsabile de funcții speciale, cum ar fi adipocitele țesutului adipos, care acumulează grăsimi ca rezervă energetică a corpului. De asemenea, este posibil să le distingem pe baza ciclului lor de viață în celule fixe (macrofage fixe, fibroblaste, adipocite), care își desfășoară întreaga viață în țesutul conjunctiv și celule migratoare (granulocite neutrofile, limfocite, macrofage) care în schimb ajung la țesutul conjunctiv din circulația sângelui. Unele dintre ele, cum ar fi limfocitele, pot trece liber din fluxul sanguin către conjunctiv, altele, cum ar fi granulocitele neutrofile, odată ce s-au deplasat prin diapedeză în conjunctiv nu mai pot reveni în sânge.
Fibroblaste

Fibroblastele sunt cele mai numeroase celule din țesutul conjunctiv propriu-zis . Funcția lor este de a produce fibrele și componentele macromoleculare ale matricei extracelulare, care este de departe cel mai abundent element al țesutului și de care depind funcțiile de susținere ale țesutului conjunctiv. Fibroblastele au în general aspectul fusiform, deși există soiuri care au și morfologii diferite, cum ar fi un aspect înstelat sau întins. În general, se găsesc dispersate în matricea creată de ele însele și, în multe cazuri, sunt dispuse de-a lungul fibrelor.
La microscopul electronic este posibil să se vadă, în zona perinucleară, aparatul Golgi și cei doi centrioli, mitocondriile sunt în general lungi și subțiri, dar în cimentoblaste și odontoblaste au și ele o formă rotunjită. Reticulul endoplasmatic are cisterne aplatizate, iar dezvoltarea sa depinde de starea funcțională a celulei, toate filamentele citoscheletului sunt foarte dezvoltate, în special microfilamentele de actină concentrate în zona corticală. Există numeroase structuri de aderență, cum ar fi podozomi și aderențe focale.
Când își încetează activitatea biosintetică , fibroblastele se transformă în fibrocite, care au citoplasmă slab acidofilă comparativ cu fibroblastele biosintetic active care au cele bazofile. Prin urmare, fibroblastele și fibrocitele reprezintă cele două momente funcționale ale aceleiași celule. Celulele cu funcții similare sunt prezente în diferitele subtipuri de țesut conjunctiv, deși în unele cazuri au particularități funcționale.
Corespunzătoare funcției fibroblastelor din alte tipuri de țesut conjunctiv sunt:
- condroblastele produc matricea țesutului cartilajului.
- osteoblastele produc matricea țesutului osos, caracterizată prin faptul că sunt calcificate .
- cementoblastele și odontoblastele produc matricea în dinți .
Macrofage
Macrofagele sunt, prin difuzie, a doua celulă cea mai numeroasă a țesutului conjunctiv propriu-zis. Macrofagele sunt de tip fix, prezente în țesutul conjunctiv în condiții normale, sau de tip migrator, care se găsește în cazul deteriorării țesuturilor ca proces inflamator. În realitate este același tip de celulă într-o formă diferită, deci este de preferat să se adopte distincția în macrofagele neactivate și macrofagele activate. La microscopul electronic, macrofagele apar ca celule rotunjite, în formă de fus sau stelate, cu un diametru de 10-30 µm, prevăzute cu proeminențe citoplasmatice de tip vilos.
În citoplasmă aparatul Golgi și reticulul endoplasmatic aspru sunt foarte dezvoltate, există și numeroși lizozomi și fagomi, un citoschelet dezvoltat cu filamente intermediare de vimentină cu grosimea de 10 nm și microfilamente asemănătoare actinei cu grosimea de 6 nm, fundamental în ceea ce constituie schela „vilozităților” macrofagului. Nucleul este unic. Deoarece la nivel citoplasmatic fibroblastele și macrofagele fixe sunt destul de similare, pentru a le distinge este necesar să se testeze capacitatea fagocitară marcată a macrofagului prin granulopexie, adică ingestia de către celulă a unui colorant vital. Deplasarea vie a macrofagelor, odată activate, este determinată de ondularea membranei lor plasmatice și este de tip amiboid. Direcția de deplasare este determinată de chimiotaxie.
Capacitatea fundamentală a macrofagelor este cu siguranță fagocitoza , un mecanism pe care îl folosesc cu o funcție defensivă. De fapt, acestea sunt responsabile de absorbția și eliminarea elementelor externe, cum ar fi virusurile , bacteriile , celulele canceroase, celulele sanguine în vârstă, moleculele dăunătoare organismului. Macrofagul este stimulat de numeroși factori chimici care se leagă de corpul străin, cum ar fi anticorpii IgG și IgM, îi recunoaște, îi activează și inițiază o serie de acțiuni care vizează distrugerea și coordonarea răspunsului imun. Pentru a-l fagocita, emite pseudopodii (proeminențe ale membranei plasmatice) care înconjoară corpul străin, încorporându-l în citoplasma macrofagului cu un sistem „balama”. Aici este digerat de hidrolazele acide conținute în lizozomi și de enzime precum lizozima, care scindează membrana plasmatică a multor bacterii și mieloperoxidaza.
Dacă corpul străin este prea mare pentru un singur macrofag, aceste celule se pot agrega în complexe polinucleare (până la o sută de nuclee), numite celule gigant ale corpului străin. Concomitent cu acțiunea fagocitară, macrofagul secretă oxid nitric (NO) și prostaglandine, care induc vasodilatație, interleukina-1 (IL-1) care atrage limfocite și granulocite neutrofile, citokine, care activează proliferarea celulelor înconjurătoare, cresc capacitatea lor fagocitică și le atrage la locul inflamației, eritropoietina , care stimulează maturarea precursorilor eritrocitelor din măduva osoasă, LCR (factori de stimulare a coloniei) care acționează asupra maturării multor alte celule hematopoietice. Macrofagele sunt, de asemenea, celule APC (celule care prezintă antigen), deoarece au antigene parțial procesate de bacterii fagocitate pe membrana lor, permițându-le să fie recunoscute de către limfocite. Cu toate acestea, această prezentare este nespecifică spre deosebire de cea a limfocitelor B.
Limfocite
Limfocitele sunt celule aparținând sistemului imunitar și, deși în mod formal celule conjunctive, se găsesc în principal libere în sânge. Acestea sunt împărțite în două clase principale: limfocite B și limfocite T :
- Limfocitele B sunt capabile să recunoască antigenul prezentat de macrofage și, ca răspuns, se maturizează în celule plasmatice , producând anticorpi care intervin pentru a elimina corpii străini.
- Limfocitele T, pe lângă cooperarea cu limfocitele B și cu proteinele complexului major de histocompatibilitate pentru a permite recunoașterea antigenelor, sunt responsabile și de răspunsul de sine , adică de eliminarea celulelor aparținând organismului însuși, modificate de infecția unui virus sau cancerigen .
Mastocitele
Mastocitele sunt celule cu un diametru de 20-30 µm, au o formă rotunjită sau fus și sunt mobile. Acestea sunt echipate cu proeminențe subtile ale membranei plasmatice, echipamente discrete mitocondriale, reticul endoplasmatic și un mic aparat Golgi. Nucleul este în formă de rinichi și are cromatină dispersată.
Cea mai importantă caracteristică morfologică pentru a le deosebi este prezența, în citoplasmă, a numeroaselor granule rotunjite și electrondense, omogene la om, solubile în apă, care sunt colorate metacromatic cu coloranți de bază precum albastru de toluidină sau cu coloranți pentru sulfat de glicozaminoglicanii, cum ar fi ca albastru Alcian. Granulele sunt acoperite cu membrană și conțin heparină și histamină. Heparina, un glicozaminoglican care conturează colorarea acestor granule, este un anticoagulant, în timp ce histamina este un vasodilatator și crește permeabilitatea capilarelor sanguine. Eliberarea acestor granule are loc în numeroase reacții imunologice, în special în cele de hipersensibilitate imediată, adică atunci când un organism intră în contact cu un antigen pentru care a fost deja sensibilizat anterior. Este suficient ca doi receptori mastocitari (asociați cu IgE în timpul primei expuneri) să intre în contact cu antigenul pentru a determina degradarea acestuia.
Degranularea constă în mișcarea granulelor către membrana plasmatică, în fuziunea membranei granulelor cu aceasta și în eliberarea conținutului granulelor în spațiul extracelular. În acest caz, se spune că are loc o degranulare asincronă. Cu toate acestea, în anumite cazuri este posibil ca răspunsul imun să se extindă asupra unor organe sau sisteme întregi și că degranularea este anafilactică. În acest caz, granulele se îmbină între ele și sunt expulzate violent din mastocit provocând șoc anafilactic. O mastocit este capabilă să-și reconstituie propriul conținut granular în 1-2 zile de la degranulare. De asemenea, sunt capabili să secrete substanțe precum interleukinele 4,5,6 (IL-4, IL-5, IL-6), citokine și factori chimiotactici . Activarea mastocitelor implică eliberarea leucotrienelor, care sunt sintetizate pornind de la acidul arahidonic conținut în unele granule lipidice mici din citoplasma celulei. Leucotrienele induc contracția mușchiului neted al căilor respiratorii și sunt implicate în atacuri de astm .
Adipocite
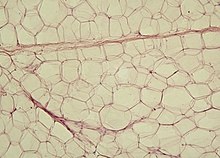
Adipocitele sunt celule fixe de țesut conjunctiv utilizate pentru colectarea, întreținerea și secreția lipidelor . Au un diametru foarte variabil, care poate depăși 100 µm, în formă rotunjită datorită picăturii lipidice unice (pentru adipocite uniloculare) care ocupă aproape întreaga citoplasmă, apăsând nucleul pe membrana plasmatică. Se găsesc în toate tipurile de țesut conjunctiv, de-a lungul vaselor de sânge și constituie tipul celular predominant în țesutul adipos. Acestea îndeplinesc o funcție de rezervă de energie, contribuind la încălzirea corpului, precum și producând hormoni (hormoni steroizi) și factori de creștere. Acestea pot fi colorate cu coloranți solubili în grăsimi precum Sudan Black, Sudan III sau Orange G. Adipocitele există în trei soiuri: adipocite uniloculare , adipocite multiloculare și adipocite pauciloculare .
- adipocitele univacuolar (canera) au un singur mare vacuolă , care conțin lipide, care umple aproape întreaga celulă. Nucleul și citoplasma celulară sunt deci descentralizate și aplatizate de-a lungul marginilor membranei plasmatice. Ele formează țesutul adipos alb.
- adipocitele multivacuolare ( Multiloculare), pe de altă parte, nu posedă vacuolul central, dar au lipidele colectate în numeroase picături mici dispersate în citoplasmă. În aceste celule nucleul se află într-o poziție centrală. Formează țesut adipos maro.
Matrice extracelulara
Toate celulele diferitelor tipuri de țesut conjunctiv se găsesc dispersate într-o substanță gelatinoasă, lichidă sau solidă numită matrice sau matrice extracelulară . Matricea extracelulară constă dintr-o porțiune fibroasă, compusă din proteine , incluse într-o soluție apoasă de proteine, glicoproteine și proteoglicani. Proteinele în cauză sunt: colagen , elastină , laminină , fibronectină , condronectină și osteonectină \ SPARC.
Prin urmare, matricea extracelulară este divizibilă în:
- o matrice de material amorf numită substanță fundamentală ;
- o componentă fibrilară.
Din punct de vedere histologic, la microscop componenta amorfă este îndepărtată în timpul proceselor de preparare: lângă componenta fibroasă există goluri albe care in vivo sunt ocupate de substanța amorfă.
Fibrele
Fibrele țesutului conjunctiv sunt scufundate în substanța amorfă și conferă stabilitate structurală matricei. Fibrele sunt împărțite în trei tipuri de bază, în funcție de compoziția și structura lor:
- fibre de colagen
- fibre reticulare
- fibre elastice
Fibrele de colagen și fibrele reticulare sunt ambele formate din molecule de procolagen , dar diferă unele de altele prin organizarea spațială a acestor molecule; fibrele elastice sunt în schimb alcătuite din două lanțuri proteice de natură diferită: fibrilină și elastină .
Fibrele de colagen
Fibrele de colagen sunt cel mai reprezentat tip de fibre în corpul uman și în țesuturile conjunctive, ele reprezentând singura cea mai abundentă componentă non-minerală după apă, constituind până la 6% din greutatea corporală. Ele apar ca fibre lungi albe ondulate, care se ramifică în mai multe direcții (în cazul unui țesut conjunctiv dens dens sau neregulat) sau într-o singură direcție (țesut conjunctiv dens dens), au o grosime cuprinsă între 1 și 12 µm. Fiecare fibră de colagen este alcătuită din zeci de fibrile mai fine, cu un diametru de 0,2-0,3 µm, care determină striația longitudinală, cufundată într-o substanță amorfă.
Fiecare fibrilă de colagen este la rândul său alcătuită din microfibrile care sunt asociate longitudinal între ele, determinând birefringența lor. Microfibrilele, examinate la microscopul electronic, apar dungate transversal spre axa lor principală, în special dungile se repetă la fiecare 70 nm când sunt proaspete sau la fiecare 64 nm când sunt uscate, prin urmare se spune că au o periodicitate axială de 64-70 nm. Se disting două tipuri de dungi transversale, una mai electrondensă și cealaltă mai puțin electrondensă. Deoarece moleculele de tropocolagen sunt asociate unele cu altele defazate, suprapunându-se pentru un sfert din lungimea lor, cele două tipuri de dungi pot fi explicate afirmând că benzile mai puțin electrondense sunt formate din capetele moleculelor de tropocolagen și de capetele cozile, în timp ce benzile mai electrondense se formează între cozile moleculelor de tropocolagen și intervalele dintre o moleculă și următoarea.
Fibrele de colagen sunt foarte rezistente la tracțiune, flexibile, dar practic inextensibile. În soluția acidă diluată, acestea tind să se umfle, în timp ce sunt dizolvate în soluții care conțin acizi sau baze puternice, precum și sunt digerate în mod specific de enzima colagenază. Denaturarea colagenului, care poate apărea prin fierbere, duce la transformarea fibrelor într-o substanță gelatinoasă. Colagenul este sintetizat în principal de fibroblaste, condroblaste și osteoblaste, dar poate fi produs și de celule epiteliale, așa cum este cazul colagenului de tip IV, care formează lamina bazală. Fibrele de colagen sunt evidențiate în microscopie ușoară prin coloranți acizi precum albastrul de anilină în tehnica de colorare Azan-Mallory, iau eozină și sunt PAS-negative sau ușor PAS-pozitive datorită lanțurilor laterale scurte de carbohidrați constând din legat de galactoză sau glucozil-galactoză la moleculele de hidroxilizină. Există 25 de tipuri diferite de lanțuri α care se asociază între ele în triplete (o moleculă de tropocolagen constă din trei helice α) pentru a forma 29 de tipuri diferite de colagen. Cele 29 de tipuri de colagen pot fi împărțite în trei clase, care sunt:
- Colagenii fibrilari: sunt cele mai frecvente fibre de colagen, singure alcătuiesc aproape tot colagenul din corpul uman, colagenul de tip I, II, III și V. le aparține. Colagenul de tip I reprezintă 90% din colagenul din corp, constituie oasele, tendoanele, fibrele de colagen ale dermei și dentinei. Colagenul de tip II este răspândit în cartilaj și în umorul vitros . Colagenul de tip III este răspândit în derm, mușchi și peretele vaselor de sânge. Colagenul de tip V este răspândit în țesutul interstițial.
- Colagenul asociat cu fibrilele : sunt fibre de colagen care nu apar niciodată singure, dar se asociază întotdeauna cu colagenii fibrilari din fibrilele lor sau formează legături între fibrile și matricea înconjurătoare. Colagenii de tip IX și XII îi aparțin, primul este asociat cu colagenul de tip II din cartilaj, al doilea este asociat cu tipul I și III în derm și tendoane.
- Colageni laminari sau reticulari : sunt fibre de colagen care nu se organizează în mănunchiuri groase ci în ochiuri reticulate, localizate adesea în spațiile pericelulare sau în membrana bazală. Include colagenii de tip IV, care constituie o mare parte a membranei bazale, VIII care este asociat cu endoteliul și X prezente în cartilajele oaselor conjugate.
Fibrele reticulare
Fibrele reticulare, formate din lanțuri de colagen de tip III, sunt răspândite în țesutul conjunctiv slab, mușchii, endoneuriul, țesutul adipos, organele limfoide și peretele vaselor de sânge. Ele sunt, de asemenea, alcătuite din fibrile și microfibrile cu periodicitate axială de 64-70 nm, dar fibrilele sunt mai subțiri (grosimea medie de 50 nm) și, prin urmare, la fel sunt și fibrele reticulare (grosime variabilă 0,5-2 µm). Fibrele reticulare nu se asociază între ele pentru a forma mănunchiuri, ci constituie bătături subțiri și rețele, care rulează pe două planuri sau în sens tridimensional, cu spații mari între ochiurile ocupate de o matrice amorfă. Nu posedă striația longitudinală a fibrelor de colagen, dar au un grad mai ridicat de glicozilare a hidroxilizinei și din acest motiv sunt PAS-pozitive, în plus sunt ușor vopsite cu metoda de impregnare a argintului și din acest motiv sunt definite și ca argirofile fibre.
Fibrele elastice

Fibrele elastice sau fibrele galbene sunt o componentă a țesuturilor conjunctive din unele structuri ale corpului. Acestea sunt fundamentale, de exemplu, în funcționalitatea arterelor, în special pentru aorta. Fibrele elastice se găsesc în matricea extracelulară împreună cu fibrele de colagen și sunt formate din proteine precum elastina și fibrilina [1] . Țesutul elastic se încadrează în categoria țesutului conjunctiv propriu-zis. Acestea sunt colorate prin orceină, ceea ce le conferă o culoare maro caracteristică sau prin metoda fuchsin-resorcinol a lui Weigert.
Substanță amorfă
Substanța amorfă (sau substanța fundamentală / sau anista ) constituie un gel compact în care fibrele sunt scufundate. Acesta constă în principal din macromolecule de glucoza din origine numite glicozaminoglicani (GAG) și asociații ale acesteia din urmă cu proteine, numite proteoglicani .
- Glicozaminoglicanii sunt componentele cele mai importante și abundente ale matricei amorfe. Aceștia sunt polimeri lungi, cu masa atomică variind de la câteva mii la milioane de Da, constând din lanțuri de dizaharide repetate de zeci de ori, formate la rândul lor de un acid uronic (D-glucuronat, L-iduronat) legat de un amino-zahăr ( N-acetil-D-glucozamină , N-acetil-D-galactozamină ). Glicozaminoglicanii pot fi sulfurați (ketaran-sulfat, condroitin-sulfat, heparan-sulfat, dermatan-sulfat, heparină) sau nesulfurat (acid hialuronic). Cel mai important glicozaminoglican este acidul hialuronic , care formează și lanțul central al agregatelor proteoglicanice. Glicozaminoglicanii sunt capabili să lege cantități semnificative de apă.
- Proteoglicanii constau în numeroși glicozaminoglicanii asociați transversal unei proteine care acționează ca un lanț central, în această stare se găsesc majoritatea glicozaminoglicanilor matricei, cu excepția acidului hialuronic care, datorită vâscozității sale ridicate, nu se leagă, ajutând pentru a forma, printre altele, lichid sinovial. Greutatea moleculară a unui proteoglican variază de la 1 la 10 milioane daltoni, din care 80-95% este compus din glicozaminoglicanii și 5-20% din proteine. Acestea sunt sintetizate în aparatul Golgi care leagă o tetrazaharidă specifică (acidul xiloză-galactoză-galactoză-glucuronic) la reziduurile de serină ale proteinei centrale și apoi adaugă câte o monozaharidă la capătul liber al tetrazaharidei. Unii dintre cei mai importanți proteoglicani sunt aggrecanul, prezent în matricea cartilajului, sindecan, versican, neurocan, decorin și β-glican. Mai mult, proteoglicanii se pot uni în jurul unei molecule de acid hialuronic central, formând structuri de ordin superior numite agregate protejcanice (sau complexe), care se numără printre cele mai mari molecule organice existente în natură, cântărind zeci de milioane de Da pentru câțiva μm. În lungime, dimensiuni comparabile la cele ale unei bacterii. Proteoglicanii, datorită structurii, vâscozității și permeabilității lor, constituie filtre moleculare excelente, care pot difuza unele substanțe cu greutate moleculară mică, pot prinde altele mai voluminoase, pot preveni atacul celulelor sanguine datorită sarcinii lor negative, pot acționa prin receptorii de pe membrana plasmatică sau mai frecvent în glicocalix .
- Glicoproteine , în cantități mai mici decât cele două categorii anterioare, printre care se evidențiază fibronectina , care, datorită interacțiunii stabilizatoare a glicozaminoglicanilor sulfați, se leagă de fibrele de colagen.
Datorită densității reduse a macromoleculelor care o constituie, substanța amorfă este transparentă și invizibilă la un microscop proaspăt. Este ușor PAS-pozitiv pentru conținutul său de glicoproteină (este intens PAS-pozitiv în cartilaj, membrane bazale și os în care concentrația de glicoproteină este mai mare), dar poate fi colorată cu metoda albastru Alcian și cu coloranți de bază. Cum ar fi anilina , care dau naștere la fenomene de metacromozie. Metacromia se datorează prezenței glicozaminoglicanilor acizi ai matricei și este mai mare cu cât aceștia sunt mai sulfurați (condrotin-sulfat, ketaran-sulfat, heparan-sulfat). Substanța amorfă conține cantități mari de apă, care cu toate acestea cu greu apar ca lichid sau țesut interstițial liber, dar sunt legate de moleculele matricei, determinând hidratarea lor. Apa legată de matrice, în care se dizolvă gaze și alte substanțe, difuzează pornind de la capilarele sanguine și funcționează ca un mijloc de dispersie și schimb între circulația sângelui și țesutul conjunctiv, permițându-le să fie hrănite. Prin urmare, se spune că substanța amorfă este elementul cu funcție trofică a țesutului conjunctiv. Cantități mari de lichid interstițial liber pot fi găsite în matricea amorfă în caz de inflamație. În plus față de funcția sa trofică, dispunerea moleculelor matricei influențează orientarea fibrelor pe care le conține și, prin țesutul său complex, împiedică răspândirea microorganismelor și a agenților patogeni.
Tipuri de țesut conjunctiv

Țesut conjunctiv
Există diferite tipuri de țesut conjunctiv, clasificate după criterii morfologice și funcționale. Cel mai comun țesut conjunctiv, denumit în general folosind acest termen, se numește țesut conjunctiv propriu-zis (adesea prescurtat ca țesut conjunctiv pd). Îndeplinește funcții de susținere și protecție, constituie baza pe care se sprijină diferitele epitelii și contribuie la apărarea organismului împotriva șocurilor și traumelor externe.
Țesutul conjunctiv propriu-zis este împărțit în:
Țesut conjunctiv dens
Se distinge prin abundența componentei fibroase colectate în fascicule, în comparație cu substanța amorfă și componenta celulară. Pentru tipul de fibre care îl compun, acesta poate fi împărțit în continuare în fibre (fibre de colagen tip I) sau elastice (fibre elastice), iar pentru aranjarea fibrelor se poate distinge în mod regulat , dacă își asumă un curs ordonat sau neregulate dacă nu își asumă un aranjament ordonat. Funcția țesutului conjunctiv dens este în principal mecanică, orientarea și calitatea fibrelor sale determină, de fapt, diferitele sale proprietăți, cum ar fi rezistența la întindere sau deformabilitatea.
- Țesut conjunctiv dens neregulat : este un țesut conjunctiv caracterizat prin numeroase fibre de colagen care se agregează în fascicule foarte dense, uneori însoțite de rețele de țesut elastic. Celulele sunt puține, există mai ales fibroblaste și macrofage rare, substanța amorfă este redusă. Se găsește în derm, în capsula fibroasă a organelor precum splina, ficatul, testiculul, ganglionii limfatici, formează teaca celor mai importanți tendoane și nervi și periostul.
- Țesut conjunctiv dens regulat : este un țesut conjunctiv caracterizat prin fibre de colagen dens ambalate, toate orientate în aceeași direcție, în conformitate cu cea a tracțiunii pe care țesutul trebuie să o suporte. Substanța amorfă este rară, celulele sunt foarte puține, care sunt aproape exclusiv fibroblaste dispuse în interstițiile subțiri ale fibrelor de colagen. Ca și în cazul neregulilor densi, fibrele de colagen pot fi asociate cu rețelele de țesut elastic. Formează tendoane, ligamente, aponevroză, stromă corneeană. Aici, fibroblastele specializate numite tendinocite sunt inserate între fasciculele paralele de fibre de colagen, a căror formă specifică de stea citoplasmatică poate fi apreciată într-o vedere transversală, dar ale căror extensii sunt confundate cu fibrele de colagen dacă sunt observate în urma unei tăieturi longitudinale. Substanța tendonului este înconjurată de un conjunctiv slab ordonat numit epitendineu. În tendoane și ligamente fibrele ating cel mai ordonat aranjament și toate sunt orientate în aceeași direcție cu fasciculele legate de țesut conjunctiv liber, în aponevroze fibrele sunt aranjate în straturi ordonate în mai multe direcții, în stroma corneei în schimb aceste straturi sunt orientate perpendicular unul pe altul.
- Tessuto connettivo denso elastico : è un connettivo caratterizzato dalla prevalenza delle fibre elastiche sulle fibre collagene, vi sono fibroblasti interposti tra i fasci di fibre elastiche, a loro volta avvolti da fibre reticolari. Forma i legamenti gialli delle vertebre, le corde vocali, le lamine fenestrate delle arterie maggiori.
Tessuto connettivo lasso
È il tessuto connettivo propriamente detto più diffuso. Si distingue per l'abbondanza della sostanza amorfa rispetto alla componente fibrosa e su quella cellulare e per il maggior numero di nuclei cellulari rispetto al connettivo denso. Per il tipo di fibre che lo compongono può essere ulteriormente classificato come:
- fibroso (fibre collagene tipo I),
- reticolare (fibre collagene di tipo III),
- elastico (fibre elastiche).
Il tessuto connettivo lasso reticolare è particolarmente diffuso negli organi emopoietici e linfoidi, nella muscolatura lisce e in alcune ghiandole; tra le sue fibre sono presenti numerosi macrofagi e fibroblasti. Uno speciale tipo di tessuto connettivo lasso è il tessuto mucoso , diffuso nell'embrione e in particolare costituente della "gelatina di Wharton", ovvero la sostanza amorfa del cordone ombelicale. Tale tessuto si definisce mucoso a causa della sua consistenza, dovuta all'abbondante quantità di acido ialuronico. Possiede poche fibre collagene o reticolari, scarsi macrofagi ma numerosi fibroblasti stellati. Se colorato, presenta intensa basofilia. Il tessuto connettivo lasso forma la tonaca propria e la tonaca sottomucosa delle mucose, avvolge molti organi e si inoltra in essi con setti che ne suddividono il parenchima in lobi e lobuli, costituisce inoltre lo stroma, la tonaca intima e la tonaca avventizia delle arterie, la tonaca media e avventizia delle vene assieme al tessuto muscolare liscio. Connette gli organi e ne riempie gli spazi liberi, circonda muscoli (epimisio, perimisio) e nervi (endonevrio, perinevrio).
Tessuto adiposo
Il tessuto adiposo , che più propriamente andrebbe chiamato organo adiposo , è un particolare tipo di tessuto connettivo. Ha un colorito giallo ed una consistenza molliccia, ed è costituito da cellule adipose, dette adipociti , che possono essere singole o riunite in gruppi nel contesto del tessuto connettivo fibrillare lasso. Se le cellule adipose sono molte, e per questo sono organizzate in lobuli, allora costituiscono il tessuto adiposo che è una varietà di tessuto connettivo lasso. Questo tessuto è presente in molte parti del corpo e, in particolare, sotto alla pelle , venendo a costituire il pannicolo adiposo (lat. panniculus diminutivo di pannus , cioè panno) cioè striscia o strato di tessuto di grasso sottocutaneo particolarmente abbondante.
Per il 50% è accumulato nel tessuto connettivo sottocutaneo dove svolge sia un'azione di copertura, che un'azione meccanica che un'azione coibente. Il 45% lo ritroviamo nella cavità addominale dove forma il tessuto adiposo interno. Il 5% lo ritroviamo nel tessuto muscolare come grasso di infiltrazione che ha la funzione di agevolare e facilitare la funzione del tessuto muscolare. Questo sottotipo di tessuto è costituito dalle cellule adipose multiloculari (al contrario dei normali adipociti non hanno un'unica goccia lipidica ma tante piccole gocce che aumentano la superficie di combustibile esposta al citosol e lo rendono quindi più disponibile per il metabolismo cellulare), è molto scarso nell'uomo adulto e appare brunastro se osservato al microscopio ottico, sia per la presenza massiccia di mitocondri che per l'elevata vascolarizzazione.
Il tessuto adiposo bruno ha esclusivamente la funzione di produrre calore perché i mitocondri delle cellule adipose multiloculari hanno meno ATP sintetasi, l'enzima che catalizza la sintesi dell'ATP, a partire dall'ADP, da fosforo inorganico e dall'energia derivante dalla respirazione cellulare. Posseggono invece una proteina canale (la termogenina) la quale dissipa il gradiente elettrochimico degli ioni idrogeno che la catena di trasporto degli elettroni normalmente produce a cavallo tra la membrana interna e lo spazio intermembrana. Questa peculiarità fa sì che l'energia prodotta dalla scissione dei trigliceridi non venga utilizzata per la produzione di ATP e venga trasformata in calore.
Il grasso bruno è ben rappresentato nei neonati di molte specie (nella specie umana soprattutto a livello della nuca, del collo e delle scapole). Negli adulti è abbondante invece quasi esclusivamente nelle specie che vanno in letargo, mentre negli adulti di altre specie, compresa quella umana, esso è scarsamente presente (l'esistenza di due diversi tipi di lipoma, cioè di neoplasie del tessuto adiposo, mostra però la permanenza di due diversi tipi di tessuto adiposo anche nell'individuo adulto). (of)
Tessuto cartilagineo
Il tessuto cartilagineo è un particolare tipo di tessuto connettivo. È costituito da fibre connettivali immerse in una sostanza amorfa molto consistente chiamata condrina e di cellule contenute in cavità lenticolari. Le cellule sono disposte in gruppi isogeni e vengono chiamate condrociti. Questo tipo di tessuto si divide in: ialino, elastico e fibroso. La Cartilagine Ialina è il tipo di Cartilagine di gran lunga più diffuso nel corpo, essa è ricca di fibre collagene immerse in una matrice "gommosa" dall'aspetto vitreo (ialino significa appunto "vitreo"). La Cartilagine Ialina ricopre le estremità, forma parte del naso, lo scheletro della laringe e dei bronchi, e congiunge le costole allo sterno. Mentre la Cartilagine Elastica è una varietà del tessuto cartilagineo ed è quella che forma il padiglione auricolare.
Tessuto osseo
Il tessuto osseo è un tipo particolare di tessuto connettivo, che svolge funzione di sostegno strutturale dell'intero organismo. La sua caratteristica principale è quella di possedere una matrice extracellulare calcificata , che fornisce al tessuto stesso notevoli doti di compattezza e di resistenza. La matrice contiene inoltre fibre, specialmente collagene, che conferiscono al tessuto un certo grado di flessibilità, e ovviamente da cellule denominate osteoblasti . In base all'organizzazione della matrice, il tessuto osseo può essere diviso in due sottotipi: tessuto osseo lamellare e tessuto osseo non lamellare .
- il tessuto osseo non lamellare, è presente nei volatili, mentre nei mammiferi rappresenta la versione immatura del tessuto osseo, ed è presente solo durante lo sviluppo dell'organismo, per essere poi rimpiazzato dal tessuto lamellare durante la crescita. In questo tipo di tessuto la matrice calcificata non è organizzata in strutture definite, ma si presenta disordinata ed irregolare
- il tessuto osseo lamellare è presente invece nell'organismo adulto, e caratterizzato dall'alto grado di organizzazione dei componenti della matrice, che sono disposte in strati, definiti appunto lamelle , altamente ordinati. Può a sua volta essere suddiviso in due tipi, a seconda del tipo di organizzazione delle lamelle: tessuto osseo spugnoso e tessuto osseo compatto.
- nel tessuto osseo spugnoso, le lamelle vanno a costituire strutture ramificate definite spicole ; per questo motivo, all'esame ottico appare come una massa spugnosa ricca di cavità intercomunicanti
- nel tessuto osseo compatto invece, le lamelle si organizzano a formare strutture concentriche, definite osteoni , addossate le une alle altre a lasciare un'unica lacuna centrale.
Tessuto ematico o sangue
Il sangue è un tessuto fluido contenuto nei vasi sanguigni dei Vertebrati , dalla composizione complessa, può essere considerato come una varietà di tessuto connettivo. È formato da una parte liquida detta siero e da una parte corpuscolare, costituita da cellule o frammenti di cellule. Ha una funzione trofica (cioè portano sostanze nutritive, ossigeno, ormoni, ecc.)
Linfa
La linfa è un altro tessuto fluido, che circola nel sistema linfatico. Si distingue dal sangue sia per la composizione molecolare del plasma, sia per il contenuto cellulare: nella linfa sono infatti del tutto assenti i globuli rossi e sono preponderanti i linfociti.
Note
- ^ Valerio Monesi, Istologia , sesta edizione, Piccin, pp. 206-207.
Bibliografia
- S. Adamo, P. Carinci, M. Molinaro, G. Siracusa, M. Stefanini, E. Ziparo (a cura di), Istologia di V. Monesi . Piccin Editore, 5ª edizione, ISBN 88-299-1639-0
- Don W. Fawcett, Bloom & Fawcett Trattato di Istologia . McGraw-Hill, 12ª edizione, ISBN 88-386-2050-4
- B. Young, JW Heath, Atlante di istologia e anatomia microscopica del Wheater . Casa Editrice Ambrosiana, 3ª edizione, ISBN 88-408-1171-0
Voci correlate
- Istologia
- Linfa (zoologia)
- Sangue
- Tessuto (biologia)
- Tessuto epiteliale
- Tessuto muscolare
- Tessuto nervoso
- Tessuto adiposo
- Tessuto cartilagineo
- Tessuto ghiandolare
- Tessuto osseo
Collegamenti esterni
- ( EN )Tessuto connettivo , su Enciclopedia Britannica , Encyclopædia Britannica, Inc.
| Controllo di autorità | Thesaurus BNCF 11717 · LCCN ( EN ) sh85031183 · GND ( DE ) 4006725-7 · BNF ( FR ) cb119348626 (data) · NDL ( EN , JA ) 00565679 |
|---|