Drosophila melanogaster
| Drosophila | |
|---|---|
 | |
| Clasificare filogenetică | |
| Domeniu | Eukaryota |
| Ordin | Diptera |
| Subordine | Brachicera |
| Infraordon | Muscomorpha |
| Secțiune | Ciclorhafa |
| Secțiune | Schizophora |
| Subsecțiune | Acaliptrate |
| Superfamilie | Ephydroidea |
| Familie | Drosophilidae |
| Subfamilie | Drosophilinae |
| Trib | Drosophilini |
| Subtrib | Drosophilina |
| Infratribu | Drosophiliti |
| Tip | Drosophila |
| Subgen | Sophophora |
| Specii | D. (Sophophora) melanogaster |
| Clasificare clasică | |
| Domeniu | Eukaryota |
| Regatul | Animalia |
| Sub-regat | Eumetazoa |
| Ramură | Bilateria |
| Phylum | Arthropoda |
| Subfilum | Tracheata |
| Superclasă | Hexapoda |
| Clasă | Insecta |
| Subclasă | Pterygota |
| Cohortă | Endopterygota |
| Superordine | Oligoneoptera |
| Secțiune | Panorpoidea |
| Ordin | Diptera |
| Subordine | Brachycera |
| Cohortă | Ciclorhafa |
| Secțiune | Schizophora |
| Subsecțiune | Acaliptrate |
| Familie | Drosophilidae |
| Tip | Drosophila |
| Specii | D. melanogaster |
| Nomenclatura binominala | |
| Drosophila melanogaster Meigen , 1830 | |
| Sinonime | |
Drosophila ampelophila Musculiță de oțet Muscă de oțet | |
Drosophila sau musca fructelor ( Drosophila (Sophophora) melanogaster Meigen , 1830 ) este o insectă din ordinul Dipterelor . Este un organism model pentru cercetarea științifică , deoarece este ușor de manipulat și are un ciclu de viață scurt.
Aspect fizic

Drosophila are ochi compuși roșii de cărămidă. Această culoare provine din prezența a 2 pigmenți: Xantomatin de culoare maro și Drosopterina , roșu [1] . Corpul este galben-maroniu, cu niște inele transversale negre pe abdomen , din aceste inele derivă denumirea științifică melanogaster (din grecescul "melanos", negru și "gaster", burta). Dimorfismul sexual este evident: femelele au 2,5 mm lungime, în timp ce masculii sunt puțin mai scurți, iar partea terminală a corpului lor este mai întunecată. Principala diferență între cele două sexe (ușor de observat la microscop ) este smocul de fire mici de păr care înconjoară anusul și organele genitale ale masculului.
Ciclu de viață
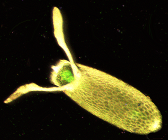
Dacă un organism Drosophila melanogaster este ținut în condiții optime la 25 ° C, acesta poate supraviețui aproximativ 50 de zile de la stadiul oului până la moarte [2] . Ciclul de viață, pe de altă parte, este de aproximativ 10-12 zile; 22-24 dacă Drosophila este plasată la 18 ° C [3] . Femelele pot depune până la 600 de ouă de-a lungul vieții în fructe sau alte materiale organice. Ouăle, al căror diametru este de aproximativ 0,5 milimetri, sunt celule mononucleare haploide .
Lichidul seminal masculin are două efecte asupra femelei: primul este reglarea procentului de ouă depuse; al doilea este să scadă dorința sexuală la femeie și astfel să împiedice împerecherea cu un alt mascul. Cu toate acestea, același spermă contribuie și la subminarea sănătății femelei prin scăderea duratei sale de viață [4] .
Ouăle fecunde diploide finalizează dezvoltarea embrionară în doar 24 de ore și clocesc imediat după aceea. Larvele rezultate cresc timp de 5-6 zile [3] , folosind microorganisme care descompun fructele și zaharurile din fructe pentru a se hrăni. La sfârșitul acestei perioade larvele se transformă în pupe și, după aproximativ 4 zile, suferă metamorfoză , la sfârșitul căreia apar insectele adulte [3] .
Femelele se împerechează la aproximativ 12 ore după metamorfoză, acumulând spermatozoizii în niște saci pentru a-l folosi mai târziu la fertilizarea ouălor. Prin urmare, geneticienii trebuie să separe femelele de restul populației înainte ca acestea să aibă șansa de a se împerechea, pentru a fi siguri că crucea este doar cu tipul particular de mascul care urmează să fie folosit în experiment. Femelele inseminate pot fi "re-virginizate" cu incubație prelungită la -10 ° C, ucigând sperma (Ashburner și colab. 2005).
Organism model
Drosophila melanogaster este un organism model , care este unul dintre cele mai studiate organisme în cercetarea biologică , în special în genetică . Motivele sunt multe:
- este o insectă mică și ușor de crescut în laborator;
- are un ciclu de viață scurt (aproximativ 2 săptămâni) și productivitate ridicată (600 de embrioni);
- larvele mature prezintă cromozomi de polietilenă în glandele salivare ;
- au doar 4 perechi de cromozomi: 3 autozomi și 1 sexual;
- masculii nu prezintă recombinații genetice, facilitând studiile genetice;
- tehnicile de transformare genetică sunt disponibile din 1987 ;
- secvențierea genomului său a fost finalizată în 1998 ;
- mutațiile genetice la specie sunt foarte frecvente.


Charles W. Woodworth este creditat că a fost primul care a crescut exemplare de Drosophila și că a sugerat lui WE Castle în timpul șederii sale la Universitatea Harvard că acestea ar putea fi utilizate pentru cercetarea genetică.
Din 1910 încoace, muștele fructelor l-au ajutat pe Thomas Hunt Morgan să își desfășoare studiile despre ereditate . Morgan și colegii au extins activitatea lui Mendel prin descrierea mecanismelor ereditare legate de cromozomul X și arătând că genele situate pe același cromozom nu au prezentat recombinații genetice. Studiile asupra caracteristicilor legate de X au contribuit la confirmarea faptului că genele se găsesc în cromozomi, în timp ce alte studii privind caracteristicile morfologice ale Drosophila au condus la primele hărți care arată locațiile genelor pe cromozomi. Prima cartografiere cromozomială a Drosophila a fost finalizată de Alfred Sturtevant .
Genomul Drosophila

Drosophila are 4 perechi de cromozomi: o pereche X / Y și trei perechi autozomale etichetate 2, 3 și 4. Al patrulea cromozom este atât de subțire încât este adesea ignorat. Genomul, care a fost complet secvențiat, conține 132 de milioane de baze și aproximativ 13.767 de gene, inclusiv, de exemplu:
- Antennapedia (gena care controlează formarea picioarelor în timpul dezvoltării)
- Lungimea antenelor (lungă / scurtă)
- Forma aripilor (lung / scurt)
- Forma picioarelor (lung / scurt)
- Culoarea ochilor (roșu / violet / maro)
- Forma aripilor (lungă / vestigială)
Asemănări cu specia umană

Din punct de vedere genetic, oamenii și musca fructelor sunt destul de asemănătoare. Aproximativ 60% din bolile genetice cunoscute pot apărea în rezerva genetică a muștei, iar aproximativ 50% din proteinele Drosophila au un analog la mamifere. Drosophila este utilizat ca model genetic pentru diferite boli umane, inclusiv tulburări neurodegenerative, cum ar fi boala Parkinson , coreea Huntington și boala Alzheimer . Musca este, de asemenea, utilizată pentru a studia mecanismul biologic al sistemului imunitar , diabetul , cancerul , inteligența, îmbătrânirea și chiar abuzul de droguri .
Nomenclatura genetică

Genele numite după alele recesive au o inițială mică, în timp ce alelele dominante au o inițială capitală. Genele numite după proteina produsă au o literă mare. Numele genelor sunt de obicei scrise cu caractere cursive. Convenția de scriere a genotipului este X / Y; 2/2; 3/3 .
În comunitatea biologilor moleculari, nomenclatura genetică a Drosophila este cunoscută pentru numele fanteziste asociate cu mutațiile genetice descoperite. Mutațiile din drojdia de bere sau alte organisme, microscopice sau de altă natură, au de obicei nume precum „cdc4” și „cdk4”; în Drosophila, pe de altă parte, nume precum „data ieftină” (literalmente „dată ieftină”, o mutație care face musca mai sensibilă la efectul etanolului ) sau „snafu” (o mutație care provoacă anomalii anatomice grotești) sunt frecvent.
Dezvoltare și embriogeneză
Embriogeneza în Drosophila a fost studiată în profunzime. Mărimea mică, ciclul de viață foarte scurt și numărul mare de descendenți tipici Drosophila îl fac o probă ideală pentru studii genetice. Este, de asemenea, singurul organism model în care diviziunea ovulului fertilizat are loc într-o cavitate sincițială.
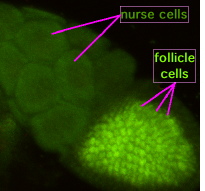
În timpul ovogenezei, ovocitul care se formează este conectat la celulele nutritive ale punților citoplasmatice . Nutrienții și alte molecule care controlează dezvoltarea sunt transferate către ovocit.
În figură se poate observa cum ovocitul care se formează este acoperit de celulele de sprijin foliculare. Aproximativ 5000 de nuclee celulare se grupează în citoplasma nedivizată a ovocitului înainte ca acestea să înceapă migrația lor la suprafață și sunt învelite de membrane plasmatice pentru a forma celulele care înconjoară sacul galbenus. La scurt timp după aceea, linia germinativă se separă de celulele somatice , prin formarea de celule polare în capătul posterior al embrionului.
Diviziunea celulară în stadiile incipiente ale embrionului Drosophila are loc atât de rapid încât nu există faze distincte, deci pot apărea erori în diviziunea ADN . Pentru a rezolva această problemă, nucleele care fac astfel de greșeli se detașează de centrosomii lor și cad în centrul embrionului, care nu va face parte din insectă.

Definiția axelor antero-posterioare și dorso-ventrale are loc conform unui mecanism numit specificație sincițială, în care gradienții genelor materne, introduși în ovocit grație celulelor hrănitoare, definesc axele corpului chiar înainte de fertilizarea ovocitului ; în special, nanosul matern și genele bicoide creează gradientul morfogen care specifică structurile anterioare ( bicoide ) și posterioare ( nanos ).
Genele materne acționează ca factori de transcripție prin activarea genelor zigotice. Primele gene zigotice care trebuie transcrise sunt clasa genelor gap, al căror nume derivă din faptul că mutația lor determină o pierdere mare de segmente în larva de mușchi. Cele mai importante gene decalaj sunt cocoasa, Kruppel, gigant, knirps și tailless fiecare dintre acestea, în funcție de gradienți de concentrație ale genelor materne, este exprimat în anumite regiuni de-a lungul anterior / posterior axa embrionului Drosophila impartirea in unitati distincte..
Combinațiile adecvate de gene gap sunt capabile să activeze amplificatori specifici ai promotorilor modulari ai genelor primare peer-rule care sunt exprimate periodic în șapte benzi de-a lungul axei anterior-posterioare, astfel delimitând un model de 14 zone discrete definite ca parasegmente. Principala caracteristică a acestei clase de gene este că unele dintre ele se vor exprima doar în parasegmentele ciudate, în timp ce altele doar în parasegmentele pare. Aceste gene sunt, de asemenea, importante pentru determinarea tiparului de expresie a genelor secundare ale regulilor de la egal la egal: acestea sunt inițial exprimate pe tot embrionul, dar în urma expresiei genelor primare de la regulile de la egal la egal, expresia lor va fi limitată în acele parasegmente care nu sunt ocupate de acestea din urmă, deoarece sunt capabil să lege potențialii genelor secundare ale regulilor de la egal la egal și să le regleze negativ expresia.
Odată exprimate genele cu regulă uniformă, acestea activează gene de polaritate segmentară care definesc partea anterioară și posterioară a unui parasegment. În special, în cele 14 parasegmente formate, rândurile de celule care prezintă o concentrație mare a produsului genetic al genelor cu regulă uniformă încep să exprime gena de polaritate segmentară numită gravată care definește marginea anterioară a parasegmentului; la rândul său, gravat acționează ca un factor de transcripție prin exprimarea genei arici care, fiind secretată de celule, interacționează cu receptorul celular plasat pe rândul de celule anterioare celor care exprimă gravat , aceasta activează o cale de semnalizare intracelulară care permite exprimarea unei alte genă de polaritate segmentară numită fără aripi care definește marginea posterioară a parasegmentului. La rândul său, Wingless este secretat și interacționează cu receptorul încrețit al rândului de celule gravate care induce o cale de semnalizare intracelulară care induce o expresie gravată suplimentară prin închiderea circuitului. Datorită acestui sistem, chiar și fără nici un stimul de către genele regulii uniforme, acest circuit se poate menține definind limitele fiecărui paraziment într-un mod definitiv.
Genele homeotice
Combinația dintre decalajul, regula colegilor și genele de polaritate segmentară exprimă ultima clasă de gene pentru dezvoltarea drosofilei numite gene selective homeotice . Aceste gene definesc cu precizie soarta de dezvoltare a fiecărui parasegment definind dacă va deveni mai degrabă o lăbuță decât o antenă.

Specificația axei dorsal-ventrală este stabilită de gradientul de concentrație al unui factor de transcripție numit dorsal. ARNm dorsal matern este plasat în ovocitul Drosophila de către celulele asistente de-a lungul întregii membrane citoplasmatice unde este tradus la 90 de minute după fertilizare. Deși dorsal este prezent pe tot ovocitul, acesta pătrunde doar în nucleul celulelor ventrale unde activează gene pentru ventralizare ( răsucire și melc ) și le reprimă pe cele pentru dorsalizare. Faptul că dorsalul intră numai în celulele ventrale se datorează poziției din citoplasma nucleului ovocitului în perioada premergătoare fecundării: fiind pe partea dorsală activează factori de transcripție care inhibă genele necesare activării dorsalei . Acest lucru nu se întâmplă pe partea ventrală, care este departe de nucleu și în acest fel dorsal își poate îndeplini funcția de ventralizare.
Genetica comportamentală și neuroștiința
În 1971 Ron Konopka și Seymour Benzer au publicat un articol intitulat „Ceasul mutanților Drosophila melanogaster ” în care au descris prima mutație care a influențat comportamentul unui animal. Muștele normale de Drosophila au prezentat un ritm de activitate și odihnă aproximativ coincident cu ziua solară (24 de ore). Mutanții Konopka și Benzer, pe de altă parte, au prezentat ritmuri mai rapide sau mai lente și, în unele cazuri, destul de neregulate: muștele mutante s-au odihnit și au fost active pentru perioade de timp complet aleatorii. Lucrările din următorii 30 de ani au arătat că aceste mutații (și altele ca acestea) afectează un grup de gene și produsele lor care formează un ceas biologic sau molecular. Acest ceas biologic a fost găsit în multe celule ale muștei, dar cele decisive pentru ciclul de activitate / odihnă sunt câteva zeci de celule din creierul central al Drosophila.
De atunci Benzer, studenții săi și mulți alții au folosit măști comportamentale pentru a izola genele implicate în viziune, miros, auz, învățare și memorie, curte, durere și alte procese biologice, cum ar fi longevitatea.
Viziunea în Drosophila

Ochiul compus al muștei fructului conține 800 de unități oculare sau ommatidia și este unul dintre cele mai sofisticate dintre toate insectele. Fiecare omatidă conține 8 celule fotoreceptoare (R1-8) și unele celule de susținere, celule pigmentate și o cornee. Midiile normale au celule pigmentare roșiatice pentru a absorbi excesul de lumină albastră și pentru a nu fi orbite de lumina zilei.
Fiecare fotoreceptor este format din două secțiuni principale, corpul celular și rabdomia. Corpul celulei conține nucleul celulei, în timp ce rândul este format din grupuri de membrane numite microvili care îl fac similar cu o periuță de dinți. Fiecare microvillo are o lungime de 1 µm până la 1,5 µm și are un diametru de 50 nm. Membrana rabdomiei conține aproximativ 100 de milioane de molecule de rodopsină , proteina vizuală care absoarbe lumina. Restul proteinelor vizuale sunt concentrate în spațiul microvillar, lăsând puțin loc citoplasmei .
Fotoreceptorii din Drosophila secretă multe varietăți de forme de rodopsină. Celulele fotoreceptoare R1-R6 sintetizează Rhodopsin1 (Rh1) care absoarbe lumina albastră (480 nm). Celulele R7 și R8 sintetizează formele Rh3 sau Rh4 care absoarbe lumina ultravioletă (345 nm și 375 nm) și Rh5 sau Rh6 care absorb lumina albastră (437 nm) și, respectiv, verde (508 nm). Fiecare moleculă de rodopsină constă dintr-o proteină numită opsină legată covalent de un cromofor carotenoid .
La fel ca în viziunea vertebratelor , transducția vizuală la nevertebrate are loc, de asemenea, printr-o cale de proteine G cuplate. Cu toate acestea, la vertebrate proteina G este transducină, în timp ce la nevertebrate este Gq (dgq în Drosophila). Când rodopsina (Rh) absoarbe un foton , cromoforul său carotenoid este izomerizat, producând o schimbare conformațională în Rh către forma sa activă, metarhodopsina. Activează Gq, care la rândul său activează o Cβ (PLCβ) fosfolipază cunoscută sub numele de NorpA.
PLCβ hidrolizează fosfatidilinozitol-4,5-bisfosfat ( PIP 2 ), un fosfolipid prezent în membrana celulară, în inozitol-trifosfat ( IP 3 ) și diacilglicerol ( DAG ). DAG, o moleculă hidrofobă , rămâne în membrană, generând deschiderea unui canal ionic selectiv pentru calciu (cunoscut sub numele de TRP, potențial de receptor tranzitor ) și, în aval, intrarea în celulă a ionilor de calciu și sodiu. Soarta IP 3 este, în schimb, de a lega receptori specifici (receptori IP 3 ) la cisternele specifice ale reticulului endoplasmatic , generând eliberarea altor ioni de calciu în citoplasmă, deși acest proces nu este considerat esențial pentru vederea normală.
Ionii de calciu leagă proteine precum calmodulina (CaM) și protein kinaza C specifică pentru viziune (PKC), cunoscută sub numele de InaC. InaC interacționează apoi cu alte proteine din aval și se crede că este principalul responsabil pentru semnalul de pornire / oprire a vederii. Arrestinele , proteine capabile să inhibe interacțiunea metarodopsinei cu Gq, joacă, de asemenea, un anumit rol în închiderea procesului vizual, inhibând transducția în amonte.
Oprirea are loc printr-un schimbător de sodiu / calciu membranar dependent de potasiu (cunoscut sub numele de NCKX30C), care pompează calciu din celulă. Acest lucru este posibil prin gradienții transmembranari favorabili de sodiu și potasiu. Pentru fiecare ion Ca ++ extrudat, intră 4 din Na + și unul din K + iese din celulă.
Proteina InaD acționează ca un suport pentru toate proteinele cheie ale căii de transducție: conține cinci domenii de legare (domenii PDZ) pentru regiunea C-terminală a moleculelor precum TRP, InaC și PLC. Mutațiile din domeniile PDZ generează probleme considerabile chiar în transducție. De exemplu, o scădere a interacțiunii dintre InaC și InaD generează o inactivare tardivă a răspunsului la lumină.
Spre deosebire de metarodopsina din vertebrate , metarodopsina din Drosophila (și Invertebrata) este convertită înapoi în rodopsină cu simpla absorbție a unui foton de lumină portocalie (580 nm).
Aproximativ două treimi din creierul Drosophila (aproximativ 200.000 de neuroni) este dedicat procesului vizual. Deși rezoluția spațială este considerabil mai mică decât cea a oamenilor, viteza răspunsului vizual este de aproape zece ori mai mare.
Zbor în Drosophila
Aripile unei muște precum Drosophila pot bate de până la 250 de ori pe secundă. Zborul este compus în esență din secțiuni liniare lungi, intercalate cu schimbări rapide de direcție numite sacade. În timpul acestor „viraje”, este capabil să se rotească cu 90 de grade în mai puțin de 50 de milisecunde.
Drosophila, și probabil multe alte muște, au nervi optici conectați direct la mușchii aripilor (în timp ce la alte insecte există, în orice caz, o trecere prin creier), făcând timpul de reacție și mai scurt, dacă este posibil.
S-a crezut mult timp că caracteristicile de zbor ale Drosophila au fost supuse mai mult vâscozității aerului decât inerției corpului animalului. Cercetări recente efectuate de Michael Dickinson și Rosalyn Sayaman au indicat în schimb că inerția este forța dominantă.
Determinarea sexului în Drosophila
Sexul în Drosophila în mod similar cu alți câțiva indivizi este un caracter cantitativ, adică acest lucru este determinat de relația dintre numărul de cromozomi X și numărul de aranjamente haploide ale autozomilor. Acest sistem de determinare a sexului se numește „Sistemul de echilibru genic”.
- Dacă această valoare este <0,5 vom avea muște metamous (adică masculi sterili și slabi)
- Dacă este 0,5 vom avea niște mușchi masculi
- Dacă este 1.0 vom avea muște femele
- Dacă este> 1.0 vom avea mușchi Metafemale (moșii cu probleme severe de dezvoltare)
- Dacă valoarea este între 0,5 și 1,0 vom avea muște intersexuale (adică cu caracteristici atât masculine, cât și feminine).
Deși relația cromozomilor X cu aspectele haploide ale autozomilor X: A prezice sexul în D. melanogaster , s-a demonstrat că rolul acestei relații este indirect și că de fapt numărul cromozomilor X determină sexul, prin acțiunea proteinelor XSE (element de semnal codat X). Conform acestui model, acțiunea autozomilor asupra numărului de cromozomi X este în mare măsură indirectă. Erickson JW și Quintero JJ au studiat, în special, haploizi (1X: 1A), deci cu cromozom X unic (masculin), dar cu fenotip feminin, și triploizi intersexuali (XX: AA), cu cromozom X dublu (feminin). Promotorul specific la femele Sex-letale (Sxl), Sxl-Pe, sa dovedit a fi activ în haploizi, deoarece o diviziune nucleară extra-celulară ridică numărul de cromozomi X, aducând nivelurile de XSE peste pragul pentru determinarea sexului feminin. Embrionii triploizi, pe de altă parte, celularizează un ciclu mai devreme decât diploizii, rezultând un bloc prematur al SxlPe. Mecanismul de autoreglare care menține nivelurile Sxl ridicate nu se dezvoltă în toate celulele și vor forma un mozaic sexual. Se presupune că în alte organisme care prezintă determinare sexuală bazată pe echilibrul genelor, cum ar fi în cazul Drosophila , raportul X: A ar putea să nu joace un rol direct. [5]
Notă
- ^ (EN) Gary D. Ewart și Anthony J. Howells, Methods in Enzymology , vol. 292, Elsevier, 1998, pp. 213-224, DOI : 10.1016 / s0076-6879 (98) 92017-1 , ISBN 978-0-12-182193-7 . Adus pe 27 august 2020 .
- ^ (EN) Nancy J. Linford, Ceyda Bilgir și Jennifer Ro, Măsurarea duratei de viață în Drosophila melanogaster , în Journal of Visualized Experiments, n. 71, 7 ianuarie 2013, p. 50068, DOI : 10.3791 / 50068 . Adus pe 27 august 2020 .
- ^ a b c Peter J. Russel, Genetica, o abordare moleculară , ediția a IV-a, Pearson, p. 496, ISBN 9788865183793 .
- ^ Arnqvist G și Rowe L (2005) Conflict sexual. Princeton University Press, Princeton New Jersey
- ^ James W Erickson și Jerome J Quintero, Efectele indirecte ale Ploidy sugerează doza de cromozom X, nu X: un raport, semnale sexuale în Drosophila , în PLoS Biology , vol. 5, nr. 12, 2007-12, DOI : 10.1371 / journal.pbio.0050332 . Adus la 17 mai 2019 .
Bibliografie
- Ashburner, M., Golic, K. și Hawley, SH „Drosophila: A Laboratory Handbook”. Cold Spring Harbor Laboratory Press, New York (2005)
- K. Haug-Collet și colab. (1999). „Clonarea și caracterizarea unui schimbător de sodiu / calciu dependent de potasiu în Drosophila ”. J. Cell Biol. 147 (3): 659-669.
- P. Raghu și colab. (2000). "Normal fototransducția Drosophila Fotoreceptorii Lipsit un receptor de Gene insp 3". Molec. & Cell. Neurosci. 15: 4289-445.
- R. Ranganathan și colab. (1995). „Transducția semnalului în fotoreceptorii Drosophila ”. Încuviințat. Pr. Nevroză. 18: 283-317.
- S. Fry și M. Dickinson (2003). „Aerodinamica manevrelor de zbor liber în Drosophila ”. Știință . 300: 495-498.
- A. Keller (2007). "Povestea lui Drosophila melanogaster ca un commensal uman". Biologie curentă 17: R77-R81.
Elemente conexe
Alte proiecte
-
 Wikimedia Commons conține imagini sau alte fișiere pe Drosophila melanogaster
Wikimedia Commons conține imagini sau alte fișiere pe Drosophila melanogaster -
 Wikispecies conține informații despre Drosophila melanogaster
Wikispecies conține informații despre Drosophila melanogaster
linkuri externe
- ( EN ) Drosophila melanogaster / Drosophila melanogaster (altă versiune) , în Encyclopedia Britannica , Encyclopædia Britannica, Inc.
- ( RO ) O introducere rapidă și ușoară a acestei insecte , pe ceolas.org .
- (RO) FlyBase , pe flybase.org.
- ( RO ) Biblioteca virtuală: Drosophila , pe ceolas.org .
- ( RO ) Proiectul Berkeley Drosophila Genome , pe fruitfly.org .
- ( EN ) Liniile directoare pentru creșterea și întreținerea Drosophila , la easyinsects.co.uk . Adus la 24 decembrie 2005 (arhivat din original la 18 februarie 2006) .
- (EN) FlyMove - baza de date a Drosophila , a flymove.uni-muenster.de.
- ( RO ) The Interactive Fly - Ghid pentru înțelegerea rolului genelor Drosophila în dezvoltare , la sdbonline.org .
- (EN) Nomenclatura genelor Drosophila , de la flynome.com. Adus la 24 decembrie 2005 (arhivat din original la 29 august 2005) .
| Controllo di autorità | Thesaurus BNCF 35002 |
|---|