Hemoglobină
Această intrare sau secțiune despre proteine nu menționează sursele necesare sau cei prezenți sunt insuficienți . |
| Hemoglobină, lanț α1 | |
|---|---|
| Gene | |
| HUGO | HBA1 |
| Entrez | 3039 |
| Locus | Chr. 16 p13.3 |
| Proteină | |
| numar CAS | |
| OMIM | 141800 |
| UniProt | Q3MIF5 |
| Hemoglobină, lanț α2 | |
|---|---|
| Gene | |
| HUGO | HBA2 |
| Entrez | 3040 |
| Locus | Chr. 16 p13.3 |
| Proteină | |
| numar CAS | |
| OMIM | 141850 |
| Hemoglobina, lanțul β | |
|---|---|
| Gene | |
| HUGO | HBB |
| Entrez | 3043 |
| Locus | Chr. 11 p15.5 |
| Proteină | |
| numar CAS | |
| OMIM | 141900 |
| UniProt | P68871 |
Hemoglobina (indicată prin abrevierea Hb sau HB ) este o proteină globulară cu o structură cuaternară formată din patru subunități . Este solubil și de culoare roșie ( cromoproteină ). Este prezent în celulele roșii din sânge ale vertebratelor (cu excepția anumitor pești din Antarctica ), pentru care îndeplinește funcția vitală de transport al „compartimentului molecular al oxigenului printr-o concentrație ridicată de O 2, sângele arterial , către țesături .
Fiecare dintre cele 4 lanțuri polipeptidice ale sale este legată covalent de un grup protetic numit hem , constând dintr-o moleculă de protoporfirină care coordonează un ion Fe 2+ de fier care iese ușor din planul moleculei. Hemoglobina este, de asemenea, o proteină alosterică . Se sintetizează inițial la nivelul proeritroblastelor policromatofile (precursori ai celulelor roșii din sânge ), rămânând apoi în concentrații mari în interiorul eritrocitului matur (care a pierdut nucleul ).
Modificările de origine genetică ale structurii primare a moleculei, care îi modifică funcția sau expresia, care modifică cantitatea în circulație, sunt denumite hemoglobinopatii (exemplele sunt anemia falciformă și talasemia ).
Structura

Hemoglobina a făcut obiectul a nenumărate lucrări. Primul care a reușit să - l cristalizeze și de a determina structura cu raze X cristalografie a fost Max Perutz în 1959 .
Structura și genele cuaternare
Este o proteină conjugată tetramerică solubilă și cântărește aproximativ 64.000 daltoni . Lanțurile tetramerului sunt fiziologic două câte două (vezi hemoglobinopatia pentru aberații): două aparținând clasei α (în galben în figură) și două din clasa β (în roșu). Grupul α include lanțurile α 1 , α 2 și ζ în timp ce grupul β include lanțurile β, Aγ, Gγ (aceste două sunt foarte asemănătoare ca structură și expresie), δ și, în final, ε. La om, genele lanțurilor α se găsesc pe cromozomul 16 și sunt intercalate cu 2 pseudogene , în timp ce cele din grupa β se află pe cromozomul 11 cu un singur pseudogen interpus.
Fiecare subunitate este în contact cu celelalte: interacțiunile α 1 -β 1 (și α 2 -β 2 ) implică 35 de aminoacizi , α 1 -β 2 (și α 2 -β 1 ) implică 19 și interfața dintre α 1 -α 2 și β 1 -β 2 sunt formate dintr-un număr variabil mic de reziduuri , întrucât este interpus un canal apos central.
Structura primară și secundară
Monomerii au o structură secundară aproape total identică, în ciuda faptului că aminoacizii în comun sunt în jur de 20%. Toate sunt formate din 8 helice α (indicate cu literele de la A la H) intercalate cu segmente de legătură scurte indicate cu perechea de litere a helicilor care o precedă și o urmează (de ex. Bucla BC). Lanțului α îi lipsește segmentul D.
Structura terțiară
Fiecare monomer are o structură terțiară similară cu cea a mioglobinei și fiecare găzduiește într-un buzunar hidrofob grupul protetic (grupul hem ), adevărata inimă a macromoleculei, legată covalent de o histidină responsabilă de funcții importante care vor fi examinate ulterior; interacțiunile ionice și hidrofobe și interacțiunile π-π cu un reziduu de fenilalanină ajută la menținerea hemului la locul său. Fiecare hem este format dintr-un complex fier-protoporfirină IX responsabil pentru legătura ușor reversibilă cu oxigenul. În proteina hemul dezoxigenat își asumă o structură ușor concavă (vezi figurile). La legarea cu oxigen, grupul protetic suferă o schimbare, aplatizându-se. „Colapsul domului” determină deplasarea lui 0,6 Å ale ionului feros și ale histidinei legate de acesta, ceea ce determină mișcarea helixului α din care face parte față de părțile proteinei (în mod ideal fixate) care leagă complexul de porfirină, provocând o schimbare conformațională care implică toate și patru subunități; consecințele acestui eveniment vor fi detaliate în secțiunea următoare.
Hemoglobina în respirație
Hemoglobina (la fel ca diferite molecule care îndeplinesc funcții similare la alte animale) este o proteină indispensabilă deoarece cantitatea de oxigen care se dizolvă în apă este în mod normal prea mică pentru nevoile metabolice ale unui animal mai mare de 1 mm; prezența sa permite să se elibereze de legea lui Henry și, astfel, să aducă oxigenul necesar în toate părțile corpului prin celulele roșii din sânge.
Reacțiile de schimb de gaze sunt posibile prin prezența ionilor de fier în fiecare grup. Acest cofactor metalic în forma sa Fe 2+ leagă oxigenul în timpul trecerii sângelui în plămâni și ulterior îl eliberează în țesuturile din circulația periferică. Trecerea oxigenului (O 2 ) are loc deoarece legăturile cu hemul sunt labile și pentru alți factori detaliați în paragraful următor.
Saturația hemoglobinei
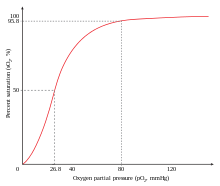
Când se leagă de oxigen, hemoglobina se numește oxihemoglobină ; în schimb în forma nelegată deoxihemoglobina .
Saturația indică raportul procentual dintre numărul mediu de molecule de oxigen (O 2 ) legate efectiv de moleculele de hemoglobină și numărul maxim de molecule de oxigen care ar putea fi legate de aceleași molecule de hemoglobină. Nu indică doar procentul de oxigen legat de hemoglobină în raport cu oxigenul total din sânge.
Deși există un procent mic de oxigen care nu este legat de hemoglobină (aproximativ 0,3 ml / 100 ml de sânge, 0,3%), restul de oxigen este legat și transportat de hemoglobină. Saturația hemoglobinei indică, prin urmare, procentul de hemoglobină utilizat în transportul oxigenului în raport cu totalul utilizabil.
Fiecare moleculă de hemoglobină este capabilă să se lege, cel mult, cu 4 molecule de O 2 și atunci când se întâmplă acest lucru, se spune că este saturată. Dacă toate moleculele de hemoglobină ar fi legate de molecule de 4 O 2 , atunci saturația ar fi de 100%; dacă în schimb, de exemplu, fiecare moleculă de hemoglobină ar fi legată la 2 molecule de oxigen, atunci saturația ar fi egală cu 50%.
Factori care afectează saturația
Saturația hemoglobinei pentru oxigen este influențată de presiunea parțială a oxigenului care, cu o legătură cooperativă, crește afinitatea hemoglobinei față de oxigen ca răspuns la o creștere a cantității de oxigen, astfel încât hemoglobina își modifică conformația pe măsură ce trece de la o stare conformațională cu afinitate scăzută pentru oxigen, numită stare T (tensionată), la una cu afinitate mare pentru oxigen, numită stare R (relaxată). Acest lucru se întâmplă la nivel alveolar, permițând captarea oxigenului de către hemoglobină, în timp ce opusul are loc la nivel periferic, unde hemoglobina trece de la starea R la starea T, eliberând oxigenul în țesuturi, care îl pot lua astfel pentru utilizare prin oxidări. . Hemoglobina are un comportament extrem de cooperant, cu un coeficient teoretic Hill de 4 și unul experimental de 2,8.
Alte influențe asupra saturației hemoglobinei pentru oxigen sunt: temperatura și 2,3-bisfosfogliceratul (BPG). Acesta din urmă, prezent în celulele roșii din sânge , reduce afinitatea hemoglobinei pentru oxigen, deoarece își stabilizează forma dezoxigenată. Cantitatea sa prezentă în celulele roșii din sânge este reglată de presiunea parțială a oxigenului din mediu pentru o eliberare optimă în țesuturi. Deci, dacă o persoană care trăiește la nivelul mării se mută la munte, unde presiunea oxigenului este mai mică, în primele zile are o lipsă de BPG și, prin urmare, o capacitate redusă de a lucra, deoarece hemoglobina are o afinitate mare pentru oxigen și nu eliberați suficient din ea în țesuturi. Pe măsură ce concentrația de BPG crește în eritrocitele sale scăzând afinitatea pentru oxigen, el poate face mai multă muncă și se spune că dobândește „pasul montan”. BPG joacă, de asemenea, un rol important în alimentarea cu oxigen a fătului. De fapt, hemoglobina fetală are o afinitate mult mai mică pentru BPG decât cea a individului adult. Acest lucru permite oxigenului să treacă de la sângele mamei la cel al fătului.
În cele din urmă, eficacitatea hemoglobinei este influențată și de prezența mai mare a CO 2 în țesuturi care, datorită comportamentului său acid în apă, eliberează protoni care se leagă de hemoglobină, favorizând formarea de punți saline caracteristice conformației T; apoi există eliberarea de oxigen. Mai mult, CO 2 care nu s-a disociat încă se leagă de grupările amino ale lanțurilor β ale hemoglobinei formând astfel carboxihemoglobină și, prin urmare, oxigenul este eliberat din aceasta. Aceste două fenomene împreună sunt numite efect Bohr .
Otravuri respiratorii
Fiziologic sau prin acțiunea anumitor substanțe oxidante ( peroxid de hidrogen , permanganat de potasiu , nitriți etc.) sau a anumitor medicamente sau a anumitor substanțe conținute în fasole (vezi favismul ), ionul Fe 2+ ( ionul feros ) se transformă în ionul Fe 3 + (ion feric) și hemoglobina se transformă în methemoglobină (MetHb), incapabilă să lege oxigenul. Prezența a 2% din MetHb este normală în sânge; dacă acest procent crește, respirația este afectată. Conversia în ion feros are loc prin enzima reducătoare methemoglobin reductază, cu o reacție în lanț care implică oxidarea NADPH și reducerea glutationului , care menține celula într-o stare reducătoare prin prevenirea formării MetHb.
Un alt pericol pentru respirație este monoxidul de carbon (CO): hemoglobina are o afinitate de aproximativ 250 de ori mai mare pentru acest gaz decât pentru oxigen, de aceea se leagă rapid de CO și legătura formată este extrem de dificil de despărțit (aproape ireversibil în mod normal). Dacă în aer există un procent de CO egal cu 1/250 cel de oxigen (aproximativ o parte la mia de aer), jumătate din hemoglobină se va combina cu monoxid de carbon, dând naștere carboxihemoglobinei (HbCO), incapabilă să lege oxigenul . Carboxihemoglobina este, de asemenea, produsă în cantități minime în interiorul țesuturilor, deoarece CO acționează ca un emițător de semnal.
Hemoglobina umană
Hemoglobina poate apărea în diferite izoforme fiziologice: Portland Hb , Hb Gower , HbF , HbA, HbA 2 .
Există, de asemenea, forme de hemoglobine patologice: Hb di Barts , HbH , HbS , HbC , HbSC , HbE , Hb Lepore , Hb Philly , Hb Genova , Hb Köln , Hb Yakima , Hb Kansas .
Legăturile chimice ale hemoglobinei determină diferite stări fizico-chimice:
- oxihemoglobina, când se leagă de O 2 .
- carboxihemoglobina (sau carbaminohemoglobina ), cand a eliberat O 2 si a captat o parte din dioxidul de carbon ( CO 2 ).
- deoxihemoglobina (sau hemoglobina redusa ), cand a eliberat O 2 . Dacă este prezent în cantități mai mari de 5 g / dl, va apărea cianoza.
- cianohemoglobina , Hb care leagă CN - ;
- carboxihemoglobina , Hb care leagă CO .
- methemoglobina , când fierul prezent în moleculă se oxidează , trecând de la starea feroasă (Fe 2+ ) la ferică (Fe 3+ ) .
- sulfohemoglobina , formată prin combinarea fierului hemic cu un atom de sulf . [1]
Hemoglobina în etapele vieții

Perioada intrauterină
Compoziția moleculei de hemoglobină variază pe parcursul vieții intrauterine și este diferită de cea a adulților (care începe să se formeze în primele luni de viață).
Principala diferență este predominanța hemoglobinei fetale (HbF) cu o afinitate foarte mare, in vivo , pentru O 2 . Această diferență duce, în condiții fiziologice, la o trecere favorabilă a O 2 de la HbA (hemoglobina adultului) a mamei la HbF a fătului. Faptul nu a fost clarificat până la descoperirea că o altă moleculă, 2,3-bisfosfogliceratul (BPG), este prezentă în condiții fiziologice și se leagă de deoxihemoglobine. HbF leagă BPG mai slab decât HbA, deoarece cele două subunități γ ale hemoglobinei fetale conțin mai puțini aminoacizi încărcați pozitiv capabili să interacționeze cu sarcinile negative ale 2,3-BPG; deoarece 2,3-BPG scade afinitatea pentru O 2 a hemoglobinelor, rezultatul în HbF este o creștere a afinității pentru O 2 a aceleiași. Același BPG este implicat și în adaptarea hemoglobinei în condiții de hipoxie (patologice sau datorate altitudinii mari).
Adult
Hemoglobina la adulți este formată din 96% HbA (α 2 β 2 ), 3 % din HbA 2 (α 2 δ 2 ) și 1% din HbF (α 2 γ 2 ).
Valoarea diagnosticului hemoglobinei
Pentru adulți, valoarea ar trebui să fie între 12 și 16 g / dL pentru femei și între 13,5 și 17 g / dL pentru bărbați. Pentru copii, valoarea minimă de 10 g / dL poate fi considerată normală. În cazul în care sunt supuși chimioterapiei, au suferit o intervenție chirurgicală majoră sau pierderi de sânge, valoarea poate scădea considerabil și în acest caz majoritatea medicilor continuă cu o transfuzie de sânge [ fără sursă ] .
În medicina criminalistică, testul Kastle-Meyer a fost folosit pentru a evidenția orice urmă de hemoglobină la locul crimei.
Notă
- ^ sulfohemoglobina în Enciclopedia Treccani , pe treccani.it . Adus la 31 octombrie 2016 .
- ^ (EN) Constance Holden, Blood and Steel (PDF), în Știință , vol. 309, nr. 5744, 30 septembrie 2005, p. 2160, DOI : 10.1126 / science.309.5744.2160d .
- ^ (EN) Moran L, Horton RA, Scrimgeour G, Perry M, Principles of Biochemistry, Boston , Pearson, 2011, p. 127, ISBN 0-321-70733-8 .
Elemente conexe
- HDW (măsura lățimii distribuției hemoglobinei)
- Mioglobina
- Carboxihemoglobina
- Eme
- Anemie cu deficit de fier
- Eritropoietina
- Variante de hemoglobină
Alte proiecte
-
 Wikționarul conține dicționarul lema « hemoglobină »
Wikționarul conține dicționarul lema « hemoglobină » -
 Wikimedia Commons conține imagini sau alte fișiere despre hemoglobină
Wikimedia Commons conține imagini sau alte fișiere despre hemoglobină
linkuri externe
- Hemoglobina , pe Treccani.it - Enciclopedii online , Institutul Enciclopediei Italiene .
- ( EN ) Hemoglobin / Hemoglobin (altă versiune) , în Encyclopedia Britannica , Encyclopædia Britannica, Inc.
- Reprezentarea 3D a moleculei , pe umass.edu .
- ( RO ) IUPAC Gold Book, „hemoglobine” , pe goldbook.iupac.org .
| Controlul autorității | Tezaur BNCF 16584 · LCCN (EN) sh85060198 · GND (DE) 4022814-9 · BNF (FR) cb11959704c (data) · NDL (EN, JA) 00.563.085 |
|---|