Extincția în masă permian-triasică

Extincția în masă a Permian-Triasic (P-Tr) , cunoscută și sub numele de Extincția Permiană , cunoscută și în mod informal în limba engleză sub numele de Great Dying [1] ( The Great Dying în italiană), a fost o extincție în masă care a avut loc în jur de 251,4 milioane cu ani în urmă, [2] [3] ca graniță între perioadele geologice permian și triasic .
A fost cel mai grav eveniment de extincție în masă care a avut loc vreodată pe Pământ , cu dispariția a 81% din speciile marine [4] și 70% din speciile de vertebrate terestre; a fost singura dispariție în masă cunoscută a insectelor . [5] [6] Se estimează că 57% din toate familiile și 83% din toate genurile au dispărut. Deoarece s-a pierdut atât de multă biodiversitate, reluarea vieții pe Pământ a fost un proces mult mai lung (presupunând 10 milioane de ani) decât alte extincții în masă. [4] Acest eveniment a fost descris ca „ mama tuturor extincțiilor în masă ”. [7]
Modelul dispariției este încă în discuție, [8] deoarece diferite studii sugerează că s-au produs una [2] până la trei [9] faze diferite. Au fost propuse mai multe mecanisme pentru a le explica: primul vârf de dispariție s-a datorat probabil unei schimbări treptate a mediului, în timp ce următorul sau mai multe s-au datorat probabil unui eveniment catastrofal. Unele scenarii posibile pentru aceste vârfuri ulterioare includ coliziuni cu obiecte astronomice, o creștere a activității vulcanice sau eliberarea bruscă de hidrați de metan de pe fundul mării; schimbările treptate includ schimbarea nivelului mării, anoxia , ariditatea crescută [10] și o schimbare a circulației curenților oceanici ca urmare a schimbărilor climatice.
Extincția întâlnirilor
| Perioadă | Podea | Varsta (Ma) | Evenimente |
|---|---|---|---|
| triasic | Anisic | 237,0–245,0 | Madrepores și bureții calcificați reapar. [11] Nivelurile carbonifere reapar. [12] Δ 13 C se stabilizează la + 2 ‰ și algele verzi reapar. [11] |
| Olenekian | 245,0–249,7 | ↑ Recuperarea florei lemnoase. [13] | |
| hindus | 249.7-251.0 | ||
| permian | Changhsingiano | 251,0–253,8 | ↑ 251,4 ± 0,03 Extincție |
| Wuchiapingian | 253,8-260,4 | ||
| Căpitan | 260,4-265,8 | ||
| Cronologia dispariției în masă permian-triasic. [14] | |||
Până la începutul secolului 21 se credea că secvențele stâncoase, cu continuitate stratigrafică, care includeau perioada permian-triasică, erau prea puține și că conțineau prea multe pauze în secvența cronostratigrafică pentru ca oamenii de știință să poată estima în mod fiabil când dispariția a avut loc, cât a durat și dacă s-a întâmplat în același timp în toate părțile lumii. [15] Cu toate acestea, unele studii ale raporturilor izotopice de uraniu-plumb efectuate pe zirconiu găsite în secvențe de roci lângă Meishan, în provincia Zhejiang , China [3] ne-au permis să datăm extincția cu 251,4 ± 0,03 milioane de ani în urmă, cu o rată ridicată și continuă de dispariție care a durat o perioadă echitabilă. [2] O mare (-9 ‰ [13] ) schimbare globală bruscă a raportului carbon-13 la carbon-12 , numită δ 13 C, coincide cu această dispariție, [16] [17] [18] [19] și este uneori folosit pentru a identifica limita Permian-Triasic în roci care nu sunt potrivite pentru datarea radiometrică. [18]
S-a emis ipoteza că granița Permian-Triasic a fost asociată cu o creștere marcată a abundenței ciupercilor marine și terestre, cauzată de creșterea rapidă corespunzătoare a cantității de plante și animale moarte. [20] Pentru o vreme acest „vârf micotic” a fost folosit de unii paleontologi pentru a identifica granița dintre cele două perioade geologice în stratificări care nu erau potrivite pentru datarea radiometrică sau care nu prezentau indicii fosile, dar susținătorii acestei teorii au subliniat, de asemenea, afirma că „vârfurile micotice” ar putea fi un fenomen frecvent creat de ecosistemul post-extincție din Triasicul inferior. [20] Ideea vârfului micotic a fost criticată din diverse motive, inclusiv: Reduviasporoniții , cei mai comuni „spori fungici” ipotizați , erau de fapt alge fosile; [13] [21] vârful nu a avut loc la nivel mondial; [22] [23] și în multe locuri nu a coincis cu granița permian-triasică. [24] Algele care au fost confundate cu spori de ciuperci pot reprezenta mai degrabă o tranziție către o lume a lacului triasic decât o fază a morții și decăderii. [25] Cu toate acestea, noi dovezi chimice sugerează o origine fungică a reduviasporoniților , diminuând aceste critici. [26]
Există încă incertitudine cu privire la durata totală a dispariției și la momentul dispariției diferitelor grupuri în cadrul întregului proces. Unele dovezi sugerează că dispariția a avut loc în câteva milioane de ani, cu un vârf acut în ultimele milioane de ani ai Permianului. [24] [27] Analizele statistice ale unor straturi bogate în fosile din Meishan, sudul Chinei, sugerează că principala dispariție s-a grupat în jurul unui vârf. [2] Unele cercetări arată că diferitele grupuri au dispărut în momente diferite; de exemplu, deși rămâne dificil de datat absolut, dispariția ostracodelor și cea a brahiopodelor au fost separate de 0,72-1,22 milioane de ani. [28] Într-o secvență bine conservată în estul Groenlandei , declinul animalelor este concentrat într-o perioadă de 10.000 până la 60.000 de ani, unde plantele au durat câteva sute de mii de ani pentru a arăta impactul complet al evenimentului. [29] O teorie anterioară, încă susținută în unele documente, [30] susține că au existat două faze majore de dispariție separate de o perioadă de 9,4 milioane de ani în care rata de dispariție a speciilor vii era încă mult sub. cu al doilea vârf ucigând „doar” 80% din speciile marine care au supraviețuit perioadei anterioare, în timp ce celelalte pierderi ar avea loc în timpul primului vârf sau în intervalul dintre cele două. Conform acestei teorii, primul vârf de dispariție s-a produs la sfârșitul erei Guadalupiene a Permianului . [31] De exemplu, toate genurile de dinocefali, cu excepția unuia, au dispărut la sfârșitul Guadalupianului, [32] la fel ca și Verbeekinidae , o familie de foraminifere mari fusulinide . [33] Se pare că impactul dispariției la sfârșitul Guadelupei asupra organismelor marine a variat între locații și grupuri taxonomice - brahipodele și coralii au suferit pierderi mari. [34] [35]
Modele de dispariție
Extincția a avut un efect atât de profund asupra ecosistemului Pământului, încât urme ale acestuia pot fi găsite aproximativ un sfert de miliard de ani mai târziu. [6] În Permianul Superior s-au găsit pe pământ multe tipuri de reptile și amfibieni, împreună cu multe plante, în special ferigi, dar și conifere și multe tipuri de ginkgo . Au existat, de asemenea, sisteme ecologice complicate de corali în mare. [36] În această perioadă continentele erau unite în supercontinentul Pangea și animalele se puteau deplasa liber. Erau jungle luxuriante, deșerturi și medii oceanice. După dispariție, un gen de vertebrate s-a aflat într-o poziție dominantă: un erbivor de dimensiuni medii numit Listrosaurus . Chiar și în mediul marin a existat prevalența unui singur tip de organism: un brahiopod numit Lingula . În cele din urmă, au reapărut și alte genuri și specii - așa-numitul „taxon Lazarus”, așa-numit cu referire la personajul biblic care a înviat din morți. Este clar că au supraviețuit cumva la dispariție, dar în număr foarte mic. [36] La fel ca dispariția de la sfârșitul lui Ordovician , pare să fi fost împărțită în două faze, separate printr-un interval de aproximativ 10 milioane de ani, cu a doua mai severă decât prima. A existat dispariția diferitelor grupuri printre brahiopode, amoniți , corali, precum și printre gastropode și, neobișnuit, printre insecte. A durat aproximativ 50 de milioane de ani pentru viață pentru a-și recupera pe deplin biodiversitatea. Nimic asemănător unui recif de corali nu a mai apărut pe fața Pământului până la 10 milioane de ani de la dispariție, iar recuperarea completă a vieții marine a durat aproximativ 100 de milioane de ani. [36]
| Extinctiile marine | Tipuri dispărute | Notă | |
|---|---|---|---|
| Nevertebrate marine | |||
| 97% | Fusulinidele au dispărut, dar au fost aproape complet dispărute înainte de catastrofă | ||
| 99% [37] | |||
| 96% | Coralii tabulați și tetracoralii au dispărut | ||
| 79% | Fenestrati, trepostomata și cryptostomata au dispărut | ||
| 96% | Ortizele și cele productive au dispărut | ||
| 59% | |||
Gastropode (melci) | 98% | ||
Amoniți ( cefalopode ) | 97% | ||
Crinoizi ( echinodermi ) | 98% | Inaptul și tovarășii au dispărut | |
Blastoizi ( echinoderme ) | 100% | Probabil că erau deja dispăruți chiar înainte de limita P-Tr | |
| 100% | În declin de la Devonian; doar două genuri erau încă în viață înainte de dispariție | ||
Euripteride („scorpioni de mare”) | 100% | Probabil că erau deja dispăruți chiar înainte de limita P-Tr | |
| 59% | |||
| Peşte | |||
| 100% | Deja în declin de la Devonian , o singură familie era încă în viață | ||
Organisme marine
Nevertebratele marine au suferit cele mai multe pierderi în timpul dispariției permian-triasice. În secțiunile reprezentate pe scară largă în sudul Chinei de limita P-Tr, de exemplu, peste 280 din 329 de tipuri de nevertebrate marine au dispărut în ultimele două zone sedimentare care conțineau conodonti Permian. [2]
Unele analize statistice ale pierderilor pe mare la sfârșitul Permianului sugerează că scăderea diversității a fost cauzată de o creștere în creștere a extincțiilor, mai degrabă decât de o scădere a speciației . [38] Extincția a afectat în primul rând organismele cu schelete compuse din carbonat de calciu, în special cele dependente de nivelurile de CO 2 din mediu pentru a-și produce scheletele. [39]
Dintre organismele bentice , evenimentul de dispariție a multiplicat ratele de dispariție și, prin urmare, a cauzat cea mai mare parte a pagubelor grupurilor taxonomice ale oricărei rate de extincție normală anterioară. [40] [41] Rata de dispariție a organismelor marine a fost catastrofală. [2] [7] [42] [43]
Grupurile de nevertebrate marine care au supraviețuit au inclus: brahiopodele articulate (cele cu știft), care au suferit un declin lent în număr până la dispariția P-Tr; Amonit ordinea ceratitids și crinoids ( „crini de mare“), care au fost aproape pe cale de disparitie, dar mai târziu a devenit abundent și diversificat.
Grupurile cu cele mai ridicate rate de supraviețuire au avut, în general, control activ al circulației, mecanisme elaborate de schimb de gaze și calcificare ușoară; organismele mai puternic calcificate și cu aparate de respirație simple au fost cele mai afectate. [44] [45] Cel puțin în cazul brahiopodelor, taxonii supraviețuitori erau în general membri mici, rari ai unei comunități diferite. [46]
Amoniții , care se aflau într-un declin îndelungat în ultimii 30 de milioane de ani de la planul Roadian (Permianul Mijlociu), au cunoscut un vârf extrem de selectiv de dispariție la sfârșitul Guadalupianului . Această dispariție a redus foarte mult diversitatea morfologică, sugerând că factorii de mediu au fost responsabili de această dispariție. Varietatea speciilor și diversitatea morfologică au fost reduse în continuare la limita P-Tr; dispariția de aici nu a fost selectivă, compatibilă cu o cauză catastrofală, de scurtă durată, cu impact global. În timpul triasicului, varietatea speciilor a crescut rapid, în timp ce diversitatea morfologică a rămas scăzută. [47] Spațiul ocupat de amoniți a devenit mai restrâns în perioada permiană. La doar câteva milioane de ani de la începutul triasicului, spațiul morfologic original a fost din nou ocupat, dar împărțit diferit între clade . [48]
Nevertebrate terestre
Permianul avea o mare diversitate de insecte și alte specii de nevertebrate, inclusiv cele mai mari insecte care au existat vreodată. Că la sfârșitul Permianului a fost singura dispariție în masă cunoscută a insectelor [5], cu dispariția a opt sau nouă ordine de insecte și a altor zece foarte reduse în diversitate. Paleodichtiopterele (insecte cu părți ale gurii capabile să usture și să suge) au intrat în declin în Permianul Mijlociu; aceste dispariții au fost legate de o schimbare a florei. Cel mai mare declin a avut loc, totuși, în timpul Permianului superior și nu a fost cauzat direct de tranzițiile în floră din cauza schimbărilor climatice. [7]
Majoritatea grupurilor de insecte fosile care au fost găsite după limita Permian-Triasic diferă semnificativ de cele care au trăit înainte de dispariția P-Tr. Cu excepția glosselitrodilor , miopterilor și protopterelor , nu s-au găsit grupuri de insecte paleozoice în depozite datate după limita P-Tr. The caloneurodes , a monurans , a paleodichthopteraids , a protelitropterans și protodonates a dispărut la sfârșitul Permian. În depozitele bine documentate din triasicul târziu, fosilele constau în principal din grupuri moderne de insecte. [5]
Plantele terestre
Răspunsul ecosistemului plantelor
Dovezile plantelor terestre sunt rare, bazate în principal pe studiul polenului și sporilor . Interesant este faptul că plantele erau relativ imune la extincția în masă, iar impactul tuturor extincțiilor majore a fost „nesemnificativ” la nivel de familie. [13] Reducerea observată a diversității speciilor (cu 50%) se poate datora, de asemenea, în principal proceselor tahonomice . [13] Cu toate acestea, a existat o reamenajare masivă a ecosistemului, cu o schimbare profundă în abundența și distribuția plantelor. [13]
La limita P - Tr, grupurile dominante de floră s-au schimbat și multe grupuri de plante terestre au intrat într-un declin brusc, cum ar fi Cordaites ( gimnosperme ) și Glossopteris ( ferigi semințe ). [49] Genurile dominante de gimnosperme au fost înlocuite cu licofite , care au colonizat zonele lăsate goale. [50]
Studiile palinologice (polen) din estul Groenlandei de straturi de roci care s-au instalat în perioada de dispariție indică păduri dense de gimnosperme înainte de evenimentul de dispariție. În același timp în care microfauna de nevertebrate marine a intrat în declin, aceste păduri mari au dispărut și au fost urmate de o creștere diversă de plante erbacee mici, inclusiv licofite , atât Selaginellales , cât și Isoetales . Mai târziu, alte grupuri de gimnosperme au revenit la flora dominantă pentru a intra din nou în declin; aceste modificări ciclice ale florei au avut loc de câteva ori în timpul perioadei de dispariție și chiar după aceea. Aceste fluctuații între taxoni arbustivi și taxoni erbacei indică stresul mediului care a dus la pierderea speciilor de plante arbustive mai mari. Succesiunea și dispariția comunităților de plante nu au coincis cu schimbarea valorilor de δ 13 C, dar au avut loc câțiva ani mai târziu. [51] Recuperarea pădurilor de gimnosperme a durat aproximativ 4-5 milioane de ani. [13]
Gama de cărbune
Nu sunt cunoscute zăcăminte de cărbune din triasicul inferior, iar zăcămintele din triasicul mediu sunt subțiri și de calitate scăzută. [52] Acest „interval de cărbune” a fost explicat în mai multe moduri. S-a speculat că au evoluat ciuperci, insecte și vertebrate noi, mai agresive, care ar ucide un număr mare de copaci. Totuși, acești descompunători au suferit și pierderi mari de specii în timpul dispariției și nu sunt considerați o cauză plauzibilă a zonei. [52] O posibilă ipoteză este că toate plantele care ar forma ulterior cărbune au dispărut în dispariția permian-triasică și că un nou set de plante a durat 10 milioane de ani pentru a se adapta la condițiile umede și acide ale mlaștinilor de turbă. [52] Pe de altă parte, pot fi imputabili alți factori abiotici (care nu sunt determinați de organismele vii), cum ar fi o scădere a precipitațiilor sau o contribuție mai mare a sedimentelor clastice . [13] În cele din urmă, este la fel de adevărat că există puține sedimente cunoscute de orice fel din Triasicul inferior și lipsa cărbunelui poate reflecta pur și simplu această deficiență. Acest lucru oferă posibilitatea ca ecosistemele tipice de producție a cărbunelui să fi reacționat la condițiile modificate mutându-se, probabil, în zone în care nu rămân urme sedimentare din Triasicul inferior. [13] De exemplu, în estul Australiei un climat rece a fost norma pentru o perioadă lungă de timp, cu un ecosistem mlăștinos specializat pentru aceste condiții. Aproximativ 95% din aceste plante producătoare de turbă au dispărut local la limita P-Tr; [53] Rămâne interesant faptul că zăcămintele de cărbune din Australia și Antarctica au dispărut semnificativ înainte de limita P-Tr. [13]
Vertebrate terestre

Grupurile care au reușit să supraviețuiască au suferit de asemenea pierderi mari în numărul de specii, iar unele grupuri de vertebrate au ajuns aproape de dispariție la sfârșitul Permianului. Unele dintre grupurile care au supraviețuit nu au supraviețuit mult după perioada extincției în masă, în timp ce alții care abia au supraviețuit au reușit să producă diferite tipuri de grupuri descendente care au continuat în evoluție. Există suficiente dovezi care să indice că peste două treimi din familiile de amfibieni terestre, sauropsidele (reptilele) și terpsidele („reptilele de tip mamifer”) au dispărut. Ierbivorele mari au suferit cele mai mari pierderi. Toate reptilele anapsid permiene au dispărut, cu excepția procolofonidelor ( broaștele țestoase au cranii anapsidice, dar se crede de obicei că au evoluat ulterior din strămoșii diapsidici). Pelicozaurii au dispărut cu puțin timp înainte de sfârșitul Permianului. S-au găsit prea puține fosile de reptile diapsidice permiene care susțin orice concluzie cu privire la efectele dispariției în masă asupra diapsidelor (grupul de reptile din care au evoluat șopârle , șerpi , crocodili și dinozauri , deci păsări ). [54] [55]
Posibile explicații ale acestor modele
Cele mai vulnerabile organisme marine au fost cele care produceau părți solide de calcar (adică din carbonat de calciu ) și aveau rate metabolice scăzute și sisteme respiratorii slabe - în special bureți calcari, corali tabulați și încrețiți, brahiopode calcaroase, briozoice și echinoderme; aproximativ 81% din aceste genuri au dispărut. Organismele înrudite care nu produceau părți solide de calcar au suferit pierderi minore, cum ar fi anemonele , din care au evoluat coralii moderni. Animalele care posedau rate metabolice ridicate, sisteme respiratorii bine dezvoltate și părți solide non-calcaroase au suferit pierderi nesemnificative - cu excepția conodontilor , dintre care 33% din genuri au dispărut. [56]

Acest model este compatibil cu efectele cunoscute ale hipoxiei , o penurie, dar nu cu o lipsă totală de oxigen . Cu toate acestea, este posibil ca hipoxia să nu fi fost singurul mecanism care a provocat dispariția organismelor marine. Majoritatea apelor de pe platoul continental ar fi devenit extrem de hipoxice, dar o astfel de catastrofă ar face dificilă explicarea tiparelor selective de dispariție. Unele modele matematice din perioadele permianului superior și ale perioadei triasice inferioare arată o scădere semnificativă și prelungită a nivelurilor de oxigen atmosferic, fără vârf în apropierea limitei P-Tr. Nivelurile minime de oxigen din atmosferă în triasicul inferior nu sunt niciodată sub nivelurile actuale - scăderea nivelurilor de oxigen nu este în concordanță cu modelul temporal de dispariție. [56]
Modelul de extincții marine observat este compatibil cu hipercapnia (niveluri excesive de dioxid de carbon ). Dioxidul de carbon sau dioxidul de carbon (CO 2 ) este foarte toxic la concentrații peste normal, reduce capacitatea pigmentului respirator de a oxigena țesuturile și face fluidele corpului mai acide, împiedicând formarea de părți solide în carbonat . La concentrații mari dioxidul de carbon provoacă narcoză (intoxicație). În plus față de aceste efecte directe, CO 2 reduce concentrația de carbonați din apă, ceea ce crește dificultatea de a produce părți solide de carbonat.
Organismele marine sunt mai sensibile decât organismele terestre la modificările nivelurilor de CO 2 , dintr-o varietate de motive. CO 2 este de 28 de ori mai solubil în apă decât oxigenul. Organismele marine funcționează în mod normal cu concentrații de CO 2 în corp mai mici decât cele de pe uscat, deoarece eliminarea CO 2 pentru animalele care respiră aer este împiedicată de necesitatea ca gazul să treacă prin membranele sistemului respirator (plămâni, trahee și ca). La organismele marine, chiar și o creștere modestă, dar susținută a CO 2, împiedică sinteza proteinelor , reduce nivelul fertilității și provoacă deformări în părțile solide de calcar. [56]
Este dificil de analizat în detaliu ratele de dispariție și supraviețuire ale organismelor terestre, deoarece puține fosile rămân de la limita Permian-Triasic. Insectele din Triasic sunt foarte diferite de cele ale Permianului, dar există un decalaj în seria fosilelor de insecte de la Permianul superior până la Triasicul inferior, de aproximativ 15 milioane de ani. Cele mai cunoscute dovezi ale modificărilor vertebratelor la limita P-Tr se găsesc în zona Karoo din Africa de Sud , dar analizele statistice nu au dat concluzii clare. [56]
Diferenții factori luați în considerare, hipercapnia , hipoxia , nivelurile ridicate de sulfură de hidrogen și încălzirea globală care au cauzat dispariția în masă a P-Tr sunt considerați ca o sinergie a factorilor fizici care au condus la distrugerea metabolismului organismelor la nivelul sfârșitul Permianului. Acești factori au persistat sau s-au repetat timp de 4-5 milioane de ani. [56]
Recuperarea biologică
Analizele timpurii indică faptul că viața de pe Pământ s-a recuperat rapid după disparițiile permiene, dar mai ales sub forma unor organisme pioniere sau grupuri taxonomice post-dezastru, cum ar fi Listrosaurusul rezistent. Cercetările indică, de asemenea, că animalele specializate care au format ecosisteme complexe, cu biodiversitate ridicată, rețele nutriționale complexe și o mare varietate de nișe, au durat mult mai mult pentru a se recupera. Se crede că această lungă recuperare s-a datorat valurilor succesive de dispariție care l-au inhibat, precum și streselor de mediu prelungite pe care le-au suferit organismele, prelungite în Triasicul inferior. Alte cercetări indică faptul că recuperarea completă nu a început până la triasicul mijlociu, la 4 sau 6 milioane de ani după dispariție; [57] și unii autori estimează că recuperarea nu a fost completă până la cel puțin 30 de milioane de ani de la dispariție, adică în Triasicul superior. [58]

În timpul triasicului inferior (4-6 Ma după dispariția P-Tr), biomasa plantelor nu a fost suficientă pentru a forma zăcăminte de cărbune, ceea ce a presupus resurse alimentare limitate pentru ierbivore. [52] Cursurile fluviale din regiunea Karoo s-au schimbat de la o structură meandrată la una care curge, sugerând că vegetația a fost foarte rară pentru o perioadă lungă de timp. [59]
Fiecare sector major al ecosistemului triasic - vegetale și animale marine și terestre - a fost dominată de un număr mic de genuri, aparent răspândite pe fața pământului, de exemplu: ierbivore therapsid Listrosaurus (care a constituit 90% din vertebrate pe de continentală a primului Triasic) și bivalve Claraia, Eumorphotis, Unionites și Promylina. Un ecosistem sănătos are un număr mare de genuri, fiecare dintre ele rezidând în câteva tipuri de habitate pe care le preferă. [49] [60]
Taxonii post-dezastru (organisme oportuniste) au profitat de ecosistemul devastat și s-au bucurat de o creștere temporară a populației și de o creștere a terenurilor disponibile. De exemplu: Lingula (un brahiopod ); stromatoliții , care fuseseră limitați la medii marginale de către ordovician ; Pleuromeia (o mică plantă subțire); Dicroidium (o ferigă). [10] [60] [61] [62]

Studii recente (2012) asupra a 15.000 de fosile de conodont recuperate în rocile din sudul Chinei, au permis o reconstrucție precisă a tendinței temperaturii suprafeței mării la începutul Triasicului, care a arătat că era atât de ridicată (între 50 și 60 ° C) pentru a face supraviețuirea organismelor marine din regiunile intertropicale ale planetei extrem de dificilă. [63] Determinarea temperaturilor a fost posibilă prin studierea raportului izotopilor de oxigen ( 18 O / 16 O) prezenți în scheletul fosil al conodontilor. [63] Temperaturile extreme au provocat o întrerupere a ciclului carbonului prin creșterea în continuare a dioxidului de carbon din atmosferă și prelungirea perioadei necesare pentru recuperarea biologică a zonelor intertropicale timp de 5 milioane de ani. [63]
Modificări ale ecosistemelor marine
Înainte de dispariție, aproximativ 67% dintre animalele marine erau atașate de fundul mării, dar în timpul Mesozoicului doar jumătate erau sesile , în timp ce restul se deplasau liber. Unele analize ale fosilelor marine ale perioadei indică o scădere a numărului de organisme sesile care se hrănesc prin filtrare, cum ar fi brahiopodele și crinii de mare , și o creștere a speciilor mobile, cum ar fi melcii , arici de mare și crabi . Înainte de dispariția permiană, ecosistemele marine simple și complexe erau ambele comune; după recuperarea de la dispariția în masă, comunitățile complexe au depășit numărul comunităților simple cu aproape trei la unu [64], iar presiunea crescută a vânătorii a dus la revoluția marină mezozoică .
Bivalvele erau destul de rare înainte de dispariția permian-triasică, dar au devenit foarte numeroase și diversificate în timpul triasicului și un grup, rudiștii , au devenit principalele componente ale recifelor mezozoice. Unii cercetători cred că majoritatea acestor schimbări au avut loc în cei cinci milioane de ani dintre cele două vârfuri majore de dispariție. [65]
Crinoizii („crinii de mare”) au suferit o dispariție selectivă, ceea ce a dus la o scădere a varietății formelor în care au crescut. [66] Radiația lor adaptivă ulterioară a fost rapidă, ducând la extinderea structurilor flexibile ale membrelor; mobilitatea, care a fost în primul rând un răspuns la prădarea crescută, a devenit o trăsătură predominantă. [67]
Vertebrate terestre
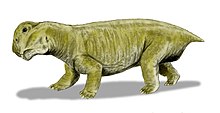
Il listrosauro , un terapside dicinodonte erbivoro della taglia di un maiale, costituiva il 90% della fauna di vertebrati terrestri del primo Triassico inferiore. [10] Sopravvissero anche altri piccoli terapsidi cinodonti carnivori, tra cui gli antenati dei mammiferi . Nella regione del Karoo , in Sudafrica , sopravvissero i terocefali Tetracynodon , Moschorhinus e Ictidosuchoides , ma sembra che non ebbero un notevole sviluppo nel Triassico. [68]
Gli arcosauri (che includevano gli antenati dei dinosauri e dei coccodrilli ) all'inizio erano meno diffusi dei terapsidi, ma cominciarono a prevalere sui terapsidi nel Triassico medio. [10] Tra il Triassico medio e quello superiore i dinosauri si evolsero da un gruppo di arcosauri e divennero il gruppo dominante dell'ecosistema terrestre per il resto del Mesozoico . [69] Questa "rivoluzione del Triassico" può aver contribuito all'evoluzione dei mammiferi costringendo i terapsidi ed i loro successori mammaliformi a vivere soprattutto come piccoli insettivori notturni; è probabile che lo stile di vita notturno abbia spinto almeno i mammaliformi a sviluppare il pelo ed alti tassi metabolici . [70]
Alcuni anfibi temnospondili ebbero una ripresa relativamente veloce, invece di estinguersi. Il mastodontosauro ed i trematosauri furono i principali predatori acquatici e semiacquatici durante la maggior parte del Triassico , alcuni di tetrapodi ed altri di pesci. [71]
I vertebrati terrestri impiegarono un tempo insolitamente lungo per riprendersi dall'estinzione di massa del Permiano; un autore stima che il recupero non fu completo fino a 30 milioni di anni dopo l'estinzione, cioè fino al Triassico superiore, nel quale abbondavano gli arcosauri (i dinosauri, gli pterosauri ed i coccodrilli), gli anfibi ed i mammaliformi. [4]
Cause dell'estinzione
Sono stati proposti vari meccanismi per spiegare le cause dell'estinzione di massa, tra cui eventi catastrofici e processi graduali, simili a quelli teorizzati per l' estinzione di massa del Cretacico–Terziario . Gli eventi catastrofici includono collisioni (anche multiple) con corpi celesti, un aumento del vulcanismo , o l'improvviso rilascio di idrati di metano. I processi graduali includono fluttuazioni del livello del mare, un' anossia , od un aumento dell' aridità . [10] Ogni ipotesi riguardo alle cause deve spiegare la selettività dell'evento, che influenzò soprattutto gli organismi con scheletri di carbonato di calcio .
Collisione

Le testimonianze che un impatto astronomico fu la causa dell'estinzione di massa del Cretacico-Terziario ha portato alla speculazione che degli impatti simili potrebbero aver causato altre estinzioni di massa, tra cui l'estinzione del Permiano-Triassico, e quindi alla ricerca di prove di collisioni databili all'età di questo evento e di crateri da impatto risalenti al periodo.
Le prove a supporto di un impatto durante il limite P–Tr includono rari granelli di quarzo lamellare in Australia ed Antartide, [72][73] fullereni che avevano intrappolato gas nobili di origine extraterrestre,[74] frammenti di meteorite in Antartide [75] e granelli ricchi di ferro, nickel e silicio, che potrebbero essere stati creati in un impatto. [76] Ad ogni modo, la veridicità di questi ritrovamenti è stata messa in discussione.[77] [78][79] [80] Il quarzo lamellare da Graphite Peak in Antartide è stato riesaminato nel 2005 con tecnologie più avanzate ed è stato concluso che le caratteristiche osservate non sono dovute ad uno shock, ma piuttosto ad una deformazione plastica, compatibile con la formazione in un ambiente tettonico come quello vulcanico. [81]
Sono stati proposti svariati crateri da impatto come possibile causa dell'estinzione P-Tr, tra cui la struttura Bedout al largo della costa nord-occidentale dell'Australia,[73] il cosiddetto cratere della Terra di Wilkes nell'Antartide orientale, [82] [83] o, anche più speculativamente, il golfo del Messico . [84] In ognuno di questi casi l'ipotesi che un impatto sia stato l'evento responsabile non è stata ancora provata, e spesso anche ampiamente criticata. Nel caso della Terra di Wilkes, l'età della struttura geofisica al di sotto dello strato di ghiaccio è molto incerta - può essere successiva all'estinzione del Permiano-Triassico.
Se la causa principale dell'estinzione P-Tr è stato un impatto, è probabile che il cratere non esista più. Dato che il 70% della superficie del nostro pianeta è ricoperto da acqua, la possibilità che un asteroide o un frammento di cometa colpisca l'oceano è due volte superiore alla possibilità che colpisca la terraferma. Inoltre la Terra non ha superficie oceanica più vecchia di 200 milioni di anni, dato che il processo dei moti convettivi e della subduzione ha ormai distrutto la crosta oceanica risalente a quel periodo. È stato inoltre speculato che i crateri formati da impatti molto grandi potrebbero essere stati mascherati da un'estesa colata lavica provocata dalla rottura o dall'indebolimento della crosta in seguito all'impatto stesso. [85]
L'idea della collisione gode anche di un altro sostegno, poiché teoricamente può aver scatenato altri fenomeni [86] considerati come cause dell'estinzione, come le eruzioni del Trappo siberiano (si veda a seguito) se fossero stati il sito dell'impatto [87] o se ne fossero trovati agli antipodi . [86] [88] La subduzione non può però essere presa come scusante del fatto che non si trovino testimonianze di un impatto; come l'estinzione alla fine del Cretacico, ci dovrebbe essere uno strato di materiali ricaduti ricco dielementi siderofili (ad esempio l' iridio ) nelle formazioni geologiche del periodo. Il brusco cambiamento provocato da un impatto spiegherebbe inoltre perché le specie non si sono evolute rapidamente adattandosi a fenomeni più lenti e su scala non globale.
Vulcanismo
Le fasi finali del periodo Permiano testimoniarono due eventi vulcanici su larga scala. Il più piccolo, che formò il Trappo Emeisciano in Cina , avvenne nello stesso periodo del picco d'estinzione alla fine del Guadalupiano , in un'area vicina all'equatore. [89] [90] Le eruzioni effusive di basalto che formarono il Trappo Siberiano costituirono uno degli eventi vulcanici più grandi che siano mai avvenuti sulla Terra e ricoprirono di lava oltre 2.000.000 di km 2 di superficie. [91] In precedenza si riteneva che le eruzioni del Trappo siberiano fossero durate per milioni di anni, ma alcune ricerche le hanno datate a 251,2 ± 0,3 milioni di anni — immediatamente prima della fine del Permiano. [2] [92]
Le eruzioni del Trappo Emeisciano e di quello Siberiano avrebbero prodotto nuvole di polvere ed aerosol acidi che avrebbero bloccato la luce solare e quindi interrotto la fotosintesi sia in mare sia sulla terraferma, facendo collassare la catena alimentare. Queste eruzioni potrebbero aver causato inoltre delle piogge acide quando gli aerosol sono stati lavati via dall'atmosfera. Ciò avrebbe ucciso le piante terrestri, i molluschi e il plancton con un guscio di carbonato di calcio. Le eruzioni avrebbero inoltre emesso anidride carbonica , causando un riscaldamento globale . Quando le nuvole di polvere e gli aerosol furono eliminati dall'atmosfera, l'anidride carbonica in eccesso sarebbe rimasta ed il riscaldamento sarebbe proseguito senza effetti mitigatori. [86]
Il Trappo Siberiano possiede caratteristiche particolari che lo resero ancora più dannoso. Effusioni di puro basalto produssero flussi di lava ad alta velocità e non scagliarono detriti nell'atmosfera. Sembra, comunque, che il 20% delle emissioni del Trappo siberiano furono piroclastiche , cioè consistettero di cenere ed altri detriti espulsi fino ad alta quota, aumentando l'effetto refrigerante a breve termine. [93] La lava basaltica eruttò o si intromise in rocce di carbonato e nei sedimenti che avrebbero dato origine a larghi giacimenti di carbone, e da qui avrebbero prodotto grandi quantità di anidride carbonica, portando ad un riscaldamento globale più forte dopo che la polvere e gli aerosol si furono depositati. [86]
Rimane in dubbio, ad ogni modo, se queste eruzioni siano state sufficienti a causare un'estinzione di massa così grave come quella alla fine del Permiano. Sono necessarie eruzioni equatoriali per produrre polvere ed aerosol sufficienti ad avere effetto sulla vita in tutto il mondo, mentre le eruzioni del Trappo Siberiano si trovavano all'interno o vicino al circolo polare artico. Inoltre se le eruzioni siberiane sono avvenute in un arco di 200.000 anni il contenuto di anidride carbonica nell'atmosfera sarebbe raddoppiato. Alcuni modelli climatici suggeriscono che un tale aumento di CO 2 avrebbe alzato la temperatura globale di 1,5 °C fino a 4,5 °C, un aumento che si ritiene improbabile come causa di una catastrofe come l'estinzione P-Tr. [86]
Comunque una teoria, resa popolare dal documentario del 2005 Miracle Planet , è che un minimo riscaldamento vulcanico abbia causato un rilascio di idrato di metano, e ciò avrebbe creato un circolo vizioso di riscaldamento, dato che il metano è 25 volte più efficiente della CO 2 nell'esacerbare il riscaldamento globale.
Idrati di metano

Gli scienziati hanno trovato testimonianze globali di un rapido decremento di circa il 10 ‰ (parti per migliaia) del rapporto 13 C / 12 C nelle rocce carbonatiche risalenti alla fine del Permiano (δ 13 C di -10 ‰). [43] [94] Questa è la prima, la più larga e la più veloce di una serie di escursioni negative e positive (aumenti e decrementi nel rapporto 13 C/ 12 C) che sono proseguite finché il tasso di isotopi non si stabilizzò repentinamente nel Triassico medio, seguito dal recupero delle forme di vita che usavano il carbonato di calcio per costruire parti solide come le conchiglie). [44]
Vari fattori possono aver contribuito a questo calo del rapporto 13 C / 12 C , ma la maggior parte non è sufficiente a spiegarlo del tutto: [95]
- I gas eruttati dai vulcani hanno un rapporto 13 C/ 12 C di circa dal 5 all'8 ‰ al di sotto dello standard (δ 13 C circa da -5 a -8 ‰). Ma la quantità necessaria per provocare una riduzione di circa il 10 ‰ su tutto il globo richiederebbe delle eruzioni più potenti di qualsiasi altra mai avvenuta sulla faccia della Terra di cui si abbia testimonianza. [96]
- Una riduzione dell'attività organica avrebbe sottratto 12 C dall'ambiente in modo più lento, così che ne sarebbe rimasto di più per incorporarsi nei sedimenti, riducendo così il rapporto 13 C/ 12 C. I processi biochimici utilizzano gli isotopi più leggeri, dato che le reazioni chimiche sono fondamentalmente condotte dalle forze elettromagnetiche tra gli atomi e gli isotopi più leggeri reagiscono più rapidamente a queste forze. Una valutazione di un piccolo decremento dal 3 al 4 ‰ nel rapporto 13 C/ 12 C (δ 13 C da -3 a -4 ‰) al Massimo termico del Paleocene-Eocene (PETM) ha però stabilito che anche trasferendo tutto il carbonio organico (degli organismi, dei suoli e dissolto negli oceani) nei sedimenti sarebbe insufficiente: anche un tale interramento di materiale ricco di 12 C non avrebbe prodotto neanche il decremento più piccolo del rapporto 13 C/ 12 C osservato nel periodo del PETM. [96]
- La materia organica sedimentaria seppellita ha un rapporto 13 C/ 12 C dal 20 al 25 ‰ al di sotto della norma (δ 13 C da -20 a -25 ‰). Teoricamente, se il livello del mare fosse calato bruscamente, i sedimenti marini superficiali sarebbero rimasti esposti all'ossidazione. Ma sarebbero stati necessari da 6.500 a 8.400 miliardi di tonnellate di carbonio organico ossidato e restituito all'oceano ed in atmosfera in meno di poche centinaia di migliaia di anni per ridurre il rapporto 13 C/ 12 C del 10 ‰. Non si ritiene che possa essere stata una possibilità realistica. [7]
- Piuttosto che un improvviso calo del livello del mare, dei periodi intermittenti di iperossia e di anossia del fondale oceanico (condizioni di alta ossigenazione / bassa ossigenazione) potrebbero aver causato le fluttuazioni nel rapporto 13 C/ 12 C nel Triassico inferiore, [44] e l'anossia globale potrebbe essere responsabile dell'estinzione alla fine del Permiano. I continenti alla fine del Permiano e nel Triassico inferiore erano più raggruppati nella zona tropicale di adesso (si veda la mappa al di sopra), e grandi fiumi tropicali avrebbero accumulato sedimenti in piccoli bacini oceanici parzialmente chiusi alle basse latitudini. Tali condizioni avrebbero favorito episodi di anossia intervallati da episodi di iperossia, i quali avrebbero rilasciato grandi quantità di carbonio organico che possiede un basso rapporto 13 C/ 12 C dato che i processi biochimici utilizzano gli isotopi più leggeri. [97] Questa od un'altra motivazione a base organica potrebbe essere stata responsabile di questa fluttuazione del rapporto 13 C/ 12 C e di quella avvenuta in precedenza nel limite Proterozoico-Cambriano. [44]
Altre ipotesi includono un avvelenamento oceanico su larga scala in seguito ad un rilascio di una grande quantità di CO 2 [98] ed una riorganizzazione a lungo termine del ciclo globale del carbonio. [95]
Ad ogni modo è stata proposta una sola causa sufficientemente potente per giustificare la riduzione globale del 10 ‰ del rapporto 13 C/ 12 C: il rilascio di metano dagli idrati di metano, [7] ed i modelli sul ciclo del carbonio confermano che questo sarebbe stato sufficiente a produrre la riduzione osservata. [95] [98] Gli idrati di metano, noti anche come clatrati di metano, sono molecole di metano intrappolate in gabbie di molecole d'acqua. Il metano viene prodotto dai metanogeni (microscopici organismi monocellulari) ed ha un rapporto 13 C/ 12 C del 60 ‰ al di sotto della norma (δ 13 C -60 ‰). Con la giusta combinazione di pressione e temperatura viene intrappolato nei clatrati molto vicino alla superficie di permafrost ed in quantità ancora maggiori ai margini della piattaforma continentale e nei fondali oceanici nelle vicinanze. Gli idrati di metano oceanici sono stati ritrovati in sedimenti risalenti a profondità di almeno 300 m sott'acqua. Possono essere trovati fino a 2.000 m di profondità, ma generalmente non si formano al di sotto dei 1.100 m. [99]
L'area coperta dalla lava delle eruzioni del Trappo Siberiano è ampia circa il doppio di quanto si riteneva inizialmente e la maggior parte di quest'area era costituita da un mare poco profondo durante la fine del Permiano. È probabile che il fondale marino contenesse depositi di idrati di metano e che la lava abbia fatto dissociare tali depositi, rilasciano grandi quantità di metano. [100]
Poiché il metano è un gas serra molto potente, ci si aspetta un enorme rilascio di metano per causare un riscaldamento globale significativo. Un'"eruzione di metano" potrebbe aver rilasciato l'equivalente di 10.000 miliardi di tonnellate di anidride carbonica - il doppio di quanto si possa trovare nei combustibili fossili sulla Terra. [36] Ci sono rilevanti testimonianze che la temperatura globale aumentò di circa 6 °C vicino all'equatore e forse ancora di più a latitudini più elevate: un'improvvisa diminuzione nel rapporto degli isotopi di ossigeno ( 18 O/ 16 O), [101] l'estinzione delle piante del tipo Glossopteris (le Glossopteris e le altre piante che crescevano nelle medesime aree), che necessitavano di un clima freddo e la sostituzione con una flora tipica di latitudini più basse. [10] [102]
Comunque il modello atteso dei cambiamenti degli isotopi come risultato di un massiccio rilascio di metano non si accorda con la situazione rilevata attraverso il Triassico inferiore. Non solo l'ipotesi del metano richiederebbe il rilascio di cinque volte la quantità di metano postulata per il PETM, [44] ma questa quantità dovrebbe essere stata riassorbita ad una velocità irrealistica per spiegare il rapido aumento del rapporto 13 C/ 12 C (episodi di δ 13 C molto positivo) durante il Triassico inferiore, prima di essere rilasciato ulteriormente varie volte. [44]
Fluttuazioni del livello del mare
Una regressione marina avviene quando le aree sommerse vicino alla linea costiera rimangono esposte al di sopra del livello del mare. Questo abbassamento del livello del mare causa una riduzione degli habitat marini superficiali, portando ad una sostituzione biotica. Gli habitat marini superficiali sono aree produttive per gli organismi alla base della catena alimentare e la loro scomparsa aumenta la competizione per le fonti di cibo. [103] Esiste qualche correlazione tra gli effetti di una regressione del livello del mare e le estinzioni di massa, ma altre testimonianze indicano che non esiste una relazione e che la regressione crea comunque altri habitat. [10] È stato ipotizzato che i cambiamenti del livello del mare ebbero come risultato dei cambiamenti nella deposizione dei sedimenti ed ebbero effetto sulla temperatura dell'acqua e sulla salinità, e quindi portando ad un declino nella diversità nel mare. [104]
Anossia
Esistono testimonianze che gli oceani divennero anossici (gravemente carenti in ossigeno) verso la fine del Permiano. Ci fu un notevole e rapido inizio di deposizione anossica nei sedimenti marini intorno alla Groenlandia orientale. [105] Il rapporto uranio / torio di vari sedimenti del Permiano indica che gli oceani erano fortemente anossici durante il periodo dell'estinzione di massa. [106]
Ciò sarebbe stato devastante per la vita nel mare, causando un'alta mortalità, ad eccezione dei batteri anaerobici che abitavano i fanghi del fondale oceanico. Ci sono inoltre testimonianze che gli eventi anossici possono causare catastrofiche emissioni di idrogeno solforato dal fondale marino.
La possibile sequenza di eventi che portarono a degli oceani anossici potrebbe aver coinvolto un periodo di riscaldamento globale che ridusse il gradiente di temperatura tra l'equatore ed i poli, cosa che avrebbe rallentato, o persino interrotto, la circolazione termoalina . Il rallentamento o l'interruzione di tale circolazione avrebbe ridotto il mescolamento dell'ossigeno nell'oceano. [106]
Ad ogni modo una ricerca ha ipotizzato che i tipi di circolazione oceanica termoalina che esistevano alla fine del Permiano non avrebbero potuto sostenere un'anossia in alto mare. [107]
Emissioni di acido solfidrico
Un evento anossico grave alla fine del Permiano potrebbe aver reso i batteri solforiduttori le specie dominanti degli ecosistemi oceanici, causando vaste emissioni di idrogeno solforato (H 2 S) che avrebbe avvelenato la vita vegetale ed animale sia sulla terraferma che nei mari, così come avrebbe ridotto gravemente lo strato di ozono , esponendo gli organismi sopravvissuti a livelli letali di radiazioni ultraviolette . [108] La fotosintesi anaerobica dei clorobi (batteri sulfurei verdi), e le conseguenti emissioni di idrogeno solforato, avvennero dalla fine del Permiano fino nel Triassico inferiore. Il fatto che questa fotosintesi anaerobica continuò nel Triassico inferiore è compatibile con le testimonianze fossili del recupero notevolmente lento dall'estinzione di massa. [109]
Questa teoria presenta il vantaggio di spiegare l'estinzione di massa delle piante, che altrimenti avrebbero dovuto prosperare in un'atmosfera con alti livelli di anidride carbonica. Le spore fossili indicano un altro fattore a sostegno della teoria: molte mostrano delle deformazioni che potrebbero essere state causate dalle radiazioni ultraviolette, che sarebbero state molto più intense dopo che le emissioni di idrogeno solforato avevano indebolito l'ozonosfera.
Il supercontinente Pangea

All'incirca a metà del periodo Permiano (nell'età kunguriana dell'epoca cisuraliana ) tutti i continenti si riunirono a formare il supercontinente Pangea , circondato dal superoceano Panthalassa , anche se alcuni blocchi dell'odierna Asia non si unirono al supercontinente fino al Permiano superiore. [110] Questa configurazione diminuì molto l'estensione degli ambienti acquatici superficiali, la sezione più produttiva del mare, ed espose gli organismi isolati delle ricche piattaforme continentali alla competizioni con altri organismi invasori. La formazione della Pangea inoltre avrebbe alterato sia la circolazione oceanica sia i modelli meteorologici dell'atmosfera, creando monsoni stagionali vicino alle coste ed un clima arido all'interno del supercontinente.
La vita marina soffrì tassi veramente alti, ma non catastrofici, di estinzione dopo la formazione della Pangea, quasi come quelli delle altre estinzioni di massa . Sembra che la formazione della Pangea non abbia causato un aumento significativo dei livelli di estinzione sulla terraferma, ed infatti l'espansione e l'aumento in diversità dei terapsidi pare che sia avvenuto nel Permiano superiore, dopo che la Pangea si era quasi completamente formata. Così sembra probabile che la Pangea abbia dato inizio ad un lungo periodo di estinzioni nel mare, ma non fosse direttamente responsabile della "grande morìa" e della fine del Permiano.
Combinazione di cause
Le possibili cause che sono sostenute da testimonianze solide sembrano descrivere una sequenza di catastrofi, ognuna peggiore delle precedenti: le eruzioni del Trappo Siberiano furono sufficientemente gravi di per sé, ma poiché avvennero vicino a delle stratificazioni di carbone ed alla piattaforma continentale, provocarono anche il rilascio di grandi quantità di anidride carbonica e di metano. [56] Il riscaldamento globale risultante potrebbe aver causato il più grave evento anossico nella storia degli oceani: secondo questa teoria, gli oceani divennero talmente anossici che gli organismi solforiduttori anaerobi finirono per dominare la biochimica degli oceani e causarono grandi emissioni di idrogeno solforato. [56]
Ad ogni modo ci possono essere alcuni collegamenti relativamente deboli all'interno di questa catena di eventi: i cambiamenti previsti nel rapporto 13 C/ 12 C come risultato di un massiccio rilascio di metano non si accordano con i modelli osservati durante il Triassico inferiore, [44] e non sembra che i tipi di circolazione termoalina oceanica che esistevano alla fine del Permiano fossero in grado di sostenere un'anossia anche in alto mare. [107]
Note
- ^ Patrick L. Barry, The Great Dying , su Science@NASA , Science and Technology Directorate, Marshall Space Flight Center, NASA, 28 gennaio 2002. URL consultato il 26 marzo 2009 (archiviato dall' url originale il 7 aprile 2009) .
- ^ a b c d e f g Jin YG, Wang Y, Wang W, Shang QH, Cao CQ, Erwin DH, Pattern of Marine Mass Extinction Near the Permian–Triassic Boundary in South China , in Science , vol. 289, n. 5478, 2000, pp. 432-436, DOI : 10.1126/science.289.5478.432 , PMID 10903200 .
- ^ a b Bowring SA, Erwin DH, Jin YG, Martin MW, Davidek K, Wang W, U/Pb Zircon Geochronology and Tempo of the End-Permian Mass Extinction , in Science , vol. 280, n. 1039, 1998, pp. 1039-1045, DOI : 10.1126/science.280.5366.1039 .
- ^ a b c Steven M. Stanley, Estimates of the magnitudes of major marine mass extinctions in earth history , Department of Geology and Geophysics, University of Hawaii, Honolulu, HI 96822, 2016.
- ^ a b c Labandeira CC, Sepkoski JJ, Insect diversity in the fossil record , in Science , vol. 261, n. 5119, 1993, pp. 310-5, PMID 11536548 .
- ^ a b Sole, RV, and Newman, M., 2002. "Extinctions and Biodiversity in the Fossil Record - Volume Two, The earth system: biological and ecological dimensions of global environment change " pp. 297-391, Encyclopedia of Global Enviromental Change John Wilely & Sons.
- ^ a b c d e Erwin DH, The great Paleozoic crisis; Life and death in the Permian , Columbia University Press, 1993, ISBN 0-231-07467-0 .
- ^ Yin H, Zhang K, Tong J, Yang Z, Wu S, The Global Stratotype Section and Point (GSSP) of the Permian-Triassic Boundary , in Episodes , vol. 24, n. 2, pp. 102-114.
- ^ Yin HF, Sweets WC, Yang ZY, Dickins JM,, Permo-Triassic Events in the Eastern Tethys , Cambridge University Press, 1992.
- ^ a b c d e f g Tanner LH, Lucas SG & Chapman MG, Assessing the record and causes of Late Triassic extinctions ( PDF ), in Earth-Science Reviews , vol. 65, n. 1-2, 2004, pp. 103-139, DOI : 10.1016/S0012-8252(03)00082-5 . URL consultato il 22 ottobre 2007 (archiviato dall' url originale il 17 gennaio 2010) .
- ^ a b Payne, JL, Lehrmann, DJ; Wei, J.; Orchard, MJ; Schrag, DP; Knoll, AH, Large Perturbations of the Carbon Cycle During Recovery from the End-Permian Extinction , in Science , vol. 305, n. 5683, 2004, pp. 506–9, DOI : 10.1126/science.1097023 , PMID 15273391 .
- ^ Retallack, JL, JJ Veevers; R. Morante, Global coal gap between Permian–Triassic extinction and Middle Triassic recovery of peat-forming plants , in Geological Society of America , vol. 108, n. 2, 2004, pp. 195-207, DOI : 10.1130/0016-7606(1996)108<0195:GCGBPT>2.3.CO;2 .
- ^ a b c d e f g h i j McElwain, JC, Punyasena, SW, Mass extinction events and the plant fossil record , in Trends in Ecology & Evolution , vol. 22, n. 10, 2007, pp. 548–557, DOI : 10.1016/j.tree.2007.09.003 , PMID 17919771 .
- ^ http://www.stratigraphy.org/ics%20chart/09_2010/StratChart2010.jpg
- ^ Erwin, DH, The Great Paleozoic Crisis: Life and Death in the Permian , New York, Columbia University Press, 1993, ISBN 0-231-07467-0 .
- ^ Magaritz, M, 13 C minima follow extinction events: a clue to faunal radiation , in Geology , vol. 17, 1989, pp. 337-340.
- ^ Krull, SJ, and Retallack, JR, 13 C depth profiles from paleosols across the Permian–Triassic boundary: Evidence for methane release , in GSA Bulletin , vol. 112, n. 9, 2000, pp. 1459-1472, DOI : 10.1130/0016-7606(2000)112%3C1459:CDPFPA%3E2.0.CO;2 .
- ^ a b Dolenec, T., Lojen, S., and Ramovs, A., The Permian-Triassic boundary in Western Slovenia (Idrijca Valley section): magnetostratigraphy, stable isotopes, and elemental variations , in Chemical Geology , vol. 175, 2001, pp. 175-190, DOI : 10.1016/S0009-2541(00)00368-5 .
- ^ Musashi, M., Isozaki, Y., Koike, T. and Kreulen, R., Stable carbon isotope signature in mid-Panthalassa shallow-water carbonates across the Permo–Triassic boundary: evidence for 13 C-depleted ocean , in Earth Planet. Sci. Lett. , vol. 193, 2001, pp. 9-20, DOI : 10.1016/S0012-821X(01)00398-3 .
- ^ a b H Visscher, H Brinkhuis, DL Dilcher, WC Elsik, Y Eshet, CV Looy, MR Rampino, and A Traverse, The terminal Paleozoic fungal event: Evidence of terrestrial ecosystem destabilization and collapse , in Proceedings of the National Academy of Sciences , vol. 93, n. 5, 1996, pp. 2155-2158. URL consultato il 20 settembre 2007 .
- ^ Foster, CB, Stephenson, MH; Marshall, C.; Logan, GA; Greenwood, PF, A Revision Of Reduviasporonites Wilson 1962: Description, Illustration, Comparison And Biological Affinities , in Palynology , vol. 26, n. 1, 2002, pp. 35-58, DOI : 10.2113/0260035 .
- ^ López-Gómez, J. and Taylor, EL, Permian-Triassic Transition in Spain: A multidisciplinary approach , in Palaeogeography, Palaeoclimatology, Palaeoecology , vol. 229, n. 1-2, 2005, pp. 1-2, DOI : 10.1016/j.palaeo.2005.06.028 .
- ^ Looy, CV, Twitchett, RJ; Dilcher, DL; Van Konijnenburg-van Cittert, JHA; Visscher, H., Life in the end-Permian dead zone , in Proceedings of the National Academy of Sciences , vol. 162, n. 4, 2005, pp. 653-659, DOI : 10.1073/pnas.131218098 , PMID 11427710 .
«See image 2» . - ^ a b Ward PD, Botha J, Buick R, De Kock MO, Erwin DH, Garrison GH, Kirschvink JL & Smith R, Abrupt and Gradual Extinction Among Late Permian Land Vertebrates in the Karoo Basin, South Africa , in Science , vol. 307, n. 5710, 2005, pp. 709-714, DOI : 10.1126/science.1107068 .
- ^ Retallack, GJ, Smith, RMH; Ward, PD, Vertebrate extinction across Permian-Triassic boundary in Karoo Basin, South Africa , in Bulletin of the Geological Society of America , vol. 115, n. 9, 2003, pp. 1133-1152, DOI : 10.1130/B25215.1 (archiviato dall' url originale il 5 maggio 2008) .
- ^ Sephton, Mark A., Visscher, Henk; Looy, Cindy V.; Verchovsky, Alexander B.; Watson, Jonathon S., Chemical constitution of a Permian-Triassic disaster species , in Geology , vol. 37, n. 10, 2009, pp. 875-878, DOI : 10.1130/G30096A.1 .
- ^ Rampino MR, Prokoph A & Adler A, Tempo of the end-Permian event: High-resolution cyclostratigraphy at the Permian–Triassic boundary , in Geology , vol. 28, n. 7, 2000, pp. 643-646, DOI : 10.1130%2F0091-7613%282000%2928%3C643%3ATOTEEH%3E2.0.CO%3B2 .
- ^ Wang, SC, Everson, PJ, Confidence intervals for pulsed mass extinction events , in Paleobiology , vol. 33, n. 2, 2007, pp. 324-336, DOI : 10.1666/06056.1 .
- ^ Twitchett RJ Looy CV Morante R Visscher H & Wignall PB, <0351:RASCOM>2.0.CO;2 Rapid and synchronous collapse of marine and terrestrial ecosystems during the end-Permian biotic crisis , in Geology , vol. 29, n. 4, 2001, pp. 351-354, DOI : 10.1130/0091-7613(2001)029<0351:RASCOM>2.0.CO;2 .
- ^ Retallack, GJ, Metzger, CA; Greaver, T.; Jahren, AH; Smith, RMH; Sheldon, ND, Middle-Late Permian mass extinction on land , in Bulletin of the Geological Society of America , vol. 118, n. 11-12, 2006, pp. 1398-1411, DOI : 10.1130/B26011.1 .
- ^ Stanley SM & Yang X, A Double Mass Extinction at the End of the Paleozoic Era , in Science , vol. 266, n. 5189, 1994, pp. 1340-1344, DOI : 10.1126/science.266.5189.1340 , PMID 17772839 .
- ^ Retallack, GJ, Metzger, CA, Jahren, AH, Greaver, T., Smith, RMH, and Sheldon, ND, Middle-Late Permian mass extinction on land , in GSA Bulletin , vol. 118, n. 11/12, novembre/dicembre 2006, pp. 1398-1411, DOI : 10.1130/B26011.1 .
- ^ Ota, A, and Isozaki, Y., Fusuline biotic turnover across the Guadalupian–Lopingian (Middle–Upper Permian) boundary in mid-oceanic carbonate buildups: Biostratigraphy of accreted limestone in Japan , in Journal of Asian Earth Sciences , vol. 26, n. 3-4, marzo 2006, pp. 353-368.
- ^ Shen, S., and Shi, GR, <0449:PEPOPB>2.0.CO;2 Paleobiogeographical extinction patterns of Permian brachiopods in the Asian-western Pacific region , in Paleobiology , vol. 28, 2002, pp. 449-463, DOI : 10.1666/0094-8373(2002)028<0449:PEPOPB>2.0.CO;2 .
- ^ Wang, XD, and Sugiyama, T., Diversity and extinction patterns of Permian coral faunas of China , in Lethaia , vol. 33, n. 4, dicembre 2000, pp. 285-294, DOI : 10.1080/002411600750053853 .
- ^ a b c d extinction
- ^ Racki G, Silica-secreting biota and mass extinctions: survival processes and patterns , vol. 154, 1–2, 1999, pp. 107-132, DOI : 10.1016/S0031-0182(99)00089-9 .
- ^ Bambach, RK, AH Knoll, SC Wang, <0522:OEAMDO>2.0.CO;2 Origination, extinction, and mass depletions of marine diversity , in Paleobiology , vol. 30, n. 4, 2004, pp. 522-542, DOI : 10.1666/0094-8373(2004)030<0522:OEAMDO>2.0.CO;2 .
- ^ Knoll, AH, Biomineralization and evolutionary history. In: PM Dove, JJ DeYoreo and S. Weiner (Eds), Reviews in Mineralogy and Geochemistry, ( PDF ), (2004) (archiviato dall' url originale il 20 giugno 2010) .
- ^ Stanley, SM, Predation defeats competition on the seafloor , in Paleobiology , vol. 34, n. 1, 2008, pp. 1-21, DOI : 10.1666/07026.1 . URL consultato il 13 maggio 2008 .
- ^ Stanley, SM, An Analysis of the History of Marine Animal Diversity , in Paleobiology , vol. 33, sp6, 2007, pp. 1-55, DOI : 10.1666/06020.1 .
- ^ McKinney, ML, Taxonomic selectivity and continuous variation in mass and background extinctions of marine taxa , in Nature , vol. 325, n. 6100, 1987, pp. 143-145, DOI : 10.1038/325143a0 .
- ^ a b Twitchett RJ, Looy CV, Morante R, Visscher H, Wignall PB, Rapid and synchronous collapse of marine and terrestrial ecosystems during the end-Permian biotic crisis , in Geology , vol. 29, n. 4, 2001, pp. 351-354, DOI : 10.1130/0091-7613(2001)029%3C0351:RASCOM%3E2.0.CO;2 .
- ^ a b c d e f g Payne, JL, Lehrmann, DJ; Wei, J.; Orchard, MJ; Schrag, DP; Knoll, AH, Large Perturbations of the Carbon Cycle During Recovery from the End-Permian Extinction , in Science , vol. 305, n. 5683, 2004, p. 506, DOI : 10.1126/science.1097023 , PMID 15273391 .
- ^ Knoll, AH, Bambach, RK; Canfield, DE; Grotzinger, JP, Comparative Earth history and Late Permian mass extinction , in Science(Washington) , vol. 273, n. 5274, 1996, pp. 452-456, DOI : 10.1126/science.273.5274.452 , PMID 8662528 .
- ^ Leighton, LR, Schneider, CL, Taxon characteristics that promote survivorship through the Permian–Triassic interval: transition from the Paleozoic to the Mesozoic brachiopod fauna , in Paleobiology , vol. 34, n. 1, 2008, pp. 65-79, DOI : 10.1666/06082.1 .
- ^ ( EN ) L. Villier, D. Korn, Morphological Disparity of Ammonoids and the Mark of Permian Mass Extinctions , in Science , vol. 306, n. 5694, 8 ottobre 2004, pp. 264-266, DOI : 10.1126/science.1102127 . URL consultato il 21 febbraio 2012 .
- ^ ( EN ) WB Saunders, E. Greenfest-Allen, DM Work, SV Nikolaeva, Morphologic and taxonomic history of Paleozoic ammonoids in time and morphospace , in Paleobiology , gennaio 2008, pp. 128-154, DOI : 10.1666/07053.1 . URL consultato il 21 febbraio 2012 .
- ^ a b Retallack, GJ, Permian–Triassic life crisis on land , in Science , vol. 267, n. 5194, 1995, pp. 77-80, DOI : 10.1126/science.267.5194.77 .
- ^ Looy, CV Brugman WA Dilcher DL & Visscher H, The delayed resurgence of equatorial forests after the Permian–Triassic ecologic crisis , in Proceedings National Academy of Sciences , vol. 96, 1999, pp. 13857-13862, PMID 10570163 .
- ^ CV Looy, Twitchett RJ, Dilcher DL, &Van Konijnenburg-Van Cittert JHA and Henk Visscher., Life in the end-Permian dead zone , in Proceedings of the National Academy of Sciences , vol. 14, n. 98, 3 luglio 2001, pp. 7879-7883, DOI : 10.1073/pnas.131218098 .
- ^ a b c d Retallack GJ, Veevers JJ, Morante R (1996). "Global coal gap between Permian–Triassic extinctions and middle Triassic recovery of peat forming plants". GSA Bulletin 108 (2): 195–207. DOI 10.1130/0016-7606(1996)108<0195:GCGBPT>2.3.CO;2
- ^ Michaelsen P, Mass extinction of peat-forming plants and the effect on fluvial styles across the Permian–Triassic boundary, northern Bowen Basin, Australia , in Palaeogeography, Palaeoclimatology, Palaeoecology , vol. 179, 3–4, 2002, pp. 173-188, DOI : 10.1016/S0031-0182(01)00413-8 .
- ^ Maxwell, WD, "Permian and Early Triassic extinction of non-marine tetrapods" , in Palaeontology , vol. 35, 1992, pp. 571-583.
- ^ Erwin DH, The End-Permian Mass Extinction , in Annual Review of Ecology and Systematics , vol. 21, 1990, pp. 69-91, DOI : 10.1146/annurev.es.21.110190.000441 .
- ^ a b c d e f g Knoll, AH, Bambach, RK, Payne, JL, Pruss, S., and Fischer, WW, Paleophysiology and end-Permian mass extinction ( PDF ), in Earth and Planetary Science Letters , vol. 256, 2007, pp. 295-313, DOI : 10.1016/j.epsl.2007.02.018 . URL consultato il 4 luglio 2008 (archiviato dall' url originale il 4 ottobre 2020) .
- ^ Lehrmann, DJ, Ramezan, J., Bowring, SA, et al. , Timing of recovery from the end-Permian extinction: Geochronologic and biostratigraphic constraints from south China , in Geology , vol. 34, n. 12, dicembre 2006, pp. 1053-1056, DOI : 10.1130/G22827A.1 .
- ^ Sahney, S. and Benton, MJ, Recovery from the most profound mass extinction of all time ( PDF ), in Proceedings of the Royal Society: Biological , vol. 275, 2008, p. 759, DOI : 10.1098/rspb.2007.1370 .
- ^ Ward PD, Montgomery DR, & Smith R, Altered river morphology in South Africa related to the Permian–Triassic extinction , in Science , vol. 289, n. 5485, 2000, pp. 1740-1743, DOI : 10.1126/science.289.5485.1740 .
- ^ a b Hallam A & Wignall PB, Mass Extinctions and their Aftermath , Oxford University Press, 1997, ISBN 978-0-19-854916-1 .
- ^ Rodland, DL & Bottjer, DJ, Biotic Recovery from the End-Permian Mass Extinction: Behavior of the Inarticulate Brachiopod Lingula as a Disaster Taxon , in Palaios , vol. 16, n. 1, 2001, pp. 95-101, DOI : 10.1669/0883-1351(2001)0163%C0095:BRFTEP3%E2.0.CO;2 .
- ^ Zi-qiang W, Recovery of vegetation from the terminal Permian mass extinction in North China , in Review of Palaeobotany and Palynology , vol. 91, 1–4, 1996, pp. 121-142, DOI : 10.1016/0034-6667(95)00069-0 .
- ^ a b c Y. Sun, MM Joachimski, PB Wignall, C. Yan, Y. Chen, H Jiang, L. Wang, X. Lai, Lethally Hot Temperatures During the Early Triassic Greenhouse , in Scienze , vol. 338, n. 6105, 19 ottobre 2012, pp. 366-370, DOI : 10.1126/science.1224126 .
- ^ Wagner PJ, Kosnik MA, & Lidgard S, Abundance Distributions Imply Elevated Complexity of Post-Paleozoic Marine Ecosystems , in Science , vol. 314, n. 5803, 2006, pp. 1289-1292, DOI : 10.1126/science.1133795 .
- ^ Clapham, ME, Bottjer, DJ and Shen, S., Decoupled diversity and ecology during the end-Guadalupian extinction (late Permian) , in Geological Society of America Abstracts with Programs , vol. 38, n. 7, 2006, p. 117. URL consultato il 28 marzo 2008 (archiviato dall' url originale l'8 dicembre 2015) .
- ^ Foote, M., Morphological diversity in the evolutionary radiation of Paleozoic and post-Paleozoic crinoids ( PDF ), in Paleobiology , vol. 25, sp1, 1999, pp. 1-116, DOI : 10.1666/0094-8373(1999)25[1:MDITER]2.0.CO;2 . URL consultato il 12 maggio 2008 .
- ^ Tomasz K. Baumiller, Crinoid Ecological Morphology , in Annual Review of Earth and Planetary Sciences , vol. 36, 2008, pp. 221-249, DOI : 10.1146/annurev.earth.36.031207.124116 . URL consultato il 15 febbraio 2012 .
- ^ Botha, J., and Smith, RMH, Lystrosaurus species composition across the Permo–Triassic boundary in the Karoo Basin of South Africa , in Lethaia , vol. 40, 2007, pp. 125-137, DOI : 10.1111/j.1502-3931.2007.00011.x . URL consultato il 2 luglio 2008 (archiviato dall' url originale il 16 dicembre 2012) . Full version online at Lystrosaurus species composition across the Permo–Triassic boundary in the Karoo Basin of South Africa ( PDF ), su nasmus.co.za . URL consultato il 2 luglio 2008 (archiviato dall' url originale il 10 settembre 2008) .
- ^ Benton, MJ, Vertebrate Paleontology , Blackwell Publishers, 2004, xii-452, ISBN 0-632-05614-2 .
- ^ Ruben, JA, and Jones, TD, Selective Factors Associated with the Origin of Fur and Feathers , in American Zoologist , vol. 40, n. 4, 2000, pp. 585-596, DOI : 10.1093/icb/40.4.585 .
- ^ Hans-Dieter Sues, Nicholas C. Fraser, Triassic life on land: the great transition , Columbia University Press, 30 giugno 2010, p. 84, ISBN 978-0-231-13522-1 . URL consultato il 21 febbraio 2012 .
- ^ Retallack GJ, Seyedolali A, Krull ES, Holser WT, Ambers CP, Kyte FT, Search for evidence of impact at the Permian–Triassic boundary in Antarctica and Australia , in Geology , vol. 26, n. 11, 1998, pp. 979-982.
- ^ a b Becker L, Poreda RJ, Basu AR, Pope KO, Harrison TM, Nicholson C, Iasky R, Bedout: a possible end-Permian impact crater offshore of northwestern Australia , in Science , vol. 304, n. 5676, 2004, pp. 1469-1476, DOI : 10.1126/science.1093925 .
- ^ Becker L, Poreda RJ, Hunt AG, Bunch TE, Rampino M, Impact event at the Permian–Triassic boundary: Evidence from extraterrestrial noble gases in fullerenes , in Science , vol. 291, n. 5508, 2001, pp. 1530-1533, DOI : 10.1126/science.1057243 .
- ^ Basu AR, Petaev MI, Poreda RJ, Jacobsen SB, Becker L, Chondritic meteorite fragments associated with the Permian–Triassic boundary in Antarctica , in Science , vol. 302, n. 5649, 2003, pp. 1388-1392, DOI : 10.1126/science.1090852 .
- ^ Kaiho K, Kajiwara Y, Nakano T, Miura Y, Kawahata H, Tazaki K, Ueshima M, Chen Z, Shi GR, End-Permian catastrophe by a bolide impact: Evidence of a gigantic release of sulfur from the mantle , in Geology , vol. 29, n. 9, 2001, pp. 815-818. URL consultato il 22 ottobre 2007 .
- ^ Farley KA, Mukhopadhyay S, Isozaki Y, Becker L, Poreda RJ, An extraterrestrial impact at the Permian–Triassic boundary? , in Science , vol. 293, n. 5539, 2001, p. 2343, DOI : 10.1126/science.293.5539.2343a .
- ^ Koeberl C, Gilmour I, Reimold WU, Philippe Claeys P, Ivanov B, End-Permian catastrophe by bolide impact: Evidence of a gigantic release of sulfur from the mantle: Comment and Reply , in Geology , vol. 30, n. 9, 2002, pp. 855-856, DOI : 10.1130/0091-7613(2002)030%3C0855:EPCBBI%3E2.0.CO;2 .
- ^ Isbell JL, Askin RA, Retallack GR, Search for evidence of impact at the Permian–Triassic boundary in Antarctica and Australia; discussion and reply , in Geology , vol. 27, n. 9, 1999, pp. 859-860, DOI : 10.1130/0091-7613(1999)027%3C0859:SFEOIA%3E2.3.CO;2 .
- ^ Koeberl K, Farley KA, Peucker-Ehrenbrink B, Sephton MA, Geochemistry of the end-Permian extinction event in Austria and Italy: No evidence for an extraterrestrial component , in Geology , vol. 32, n. 12, 2004, pp. 1053-1056, DOI : 10.1130/G20907.1 .
- ^ Langenhorst F, Kyte FT & Retallack GJ, Reexamination of quartz grains from the Permian–Triassic boundary section at Graphite Peak, Antarctica ( PDF ), Lunar and Planetary Science Conference XXXVI , 2005. URL consultato il 13 luglio 2007 .
- ^ von Frese RR, Potts L, Gaya-Pique L, Golynsky AV, Hernandez O, Kim J, Kim H & Hwang J, Permian–Triassic mascon in Antarctica , in Eos Trans. AGU, Jt. Assem. Suppl. , vol. 87, n. 36, 2006, pp. Abstract T41A-08. URL consultato il 22 ottobre 2007 (archiviato dall' url originale il 30 settembre 2007) .
- ^ RRB Von Frese, LV Potts, SB Wells, TE Leftwich, HR Kim, JW Kim, AV Golynsky, O. Hernandez, and LR Gaya-Piqué, GRACE gravity evidence for an impact basin in Wilkes Land, Antarctica , in Geochem. Geophys. Geosyst. , vol. 10, 2009, pp. Q02014, DOI : 10.1029/2008GC002149 .
- ^ MS Stanton, Is the Gulf's Origin Heaven Sent? , in Explorer , AAPG, American Association of Petroleum geologist, 2002, pp. 1-6. URL consultato il marzo 2012 (archiviato dall' url originale l'11 settembre 2008) .
- ^ Jones AP, Price GD, Price NJ, DeCarli PS, Clegg RA, Impact induced melting and the development of large igneous provinces , in Earth and Planetary Science Letters , vol. 202, n. 3, 2002, pp. 551-561, DOI : 10.1016/S0012-821X(02)00824-5 .
- ^ a b c d e White RV, Earth's biggest 'whodunnit': unravelling the clues in the case of the end-Permian mass extinction ( PDF ), in Phil. Trans. Royal Society of London , vol. 360, 2002, pp. 2963-2985, DOI : 10.1098/rsta.2002.1097 . URL consultato il 12 gennaio 2008 .
- ^ Ager, H. Bradford, Cambridge Conference, Giant Impact Craters Lead To Flood Basalts: A Viable Model , CCNet 33/2001: Abstract 50470 , 16 novembre 2000.
- ^ Hagstrum, Jonathan T, Large Oceanic Impacts As The Cause Of Antipodal Hotspots And Global Mass Extinctions , CCNet 33/2001: Abstract 50288 , 2001.
- ^ Zhou, MF., Malpas, J, Song, XY, Robinson, PT, Sun, M, Kennedy, AK, Lesher, CM & Keays, RR, A temporal link between the Emeishan large igneous province (SW China) and the end-Guadalupian mass extinction , in Earth and Planetary Science Letters , vol. 196, 3–4, 2002, pp. 113-122, DOI : 10.1016/S0012-821X(01)00608-2 .
- ^ Paul B. Wignall et al. , Volcanism, Mass Extinction, and Carbon Isotope Fluctuations in the Middle Permian of China , in Science , vol. 324, n. 5931, 2009, pp. 1179-1182, DOI : 10.1126/science.1171956 .
- ^ Andy Saunders, Marc Reichow, The Siberian Traps - Area and Volume , su le.ac.uk , 2009. URL consultato il 18 ottobre 2009 .
- ^ Mundil, R., Ludwig, KR, Metcalfe, I. & Renne, PR, Age and Timing of the Permian Mass Extinctions: U/Pb Dating of Closed-System Zircons , in Science , vol. 305, n. 5691, 2004, pp. 1760-1763, DOI : 10.1126/science.1101012 .
- ^ "Permian–Triassic Extinction - Volcanism" , su hoopermuseum.earthsci.carleton.ca . URL consultato il 15 marzo 2012 .
- ^ Palfy J, Demeny A, Haas J, Htenyi M, Orchard MJ, & Veto I, Carbon isotope anomaly at the Triassic– Jurassic boundary from a marine section in Hungary , in Geology , vol. 29, n. 11, 2001, pp. 1047-1050, DOI : 10.1130/0091-7613(2001)029%3C1047:CIAAOG%3E2.0.CO;2 .
- ^ a b c Berner, RA, Examination of hypotheses for the Permo-Triassic boundary extinction by carbon cycle modeling , in Proceedings of the National Academy of Sciences , vol. 99, 2002, p. 32095199, DOI : 10.1073/pnas.032095199 , PMID 11917102 .
- ^ a b Dickens GR, O'Neil JR, Rea DK & Owen RM, Dissociation of oceanic methane hydrate as a cause of the carbon isotope excursion at the end of the Paleocene , in Paleoceanography , vol. 10, n. 6, 1995, pp. 965-71, DOI : 10.1029/95PA02087 .
- ^ Schrag, DP, Berner, RA, Hoffman, PF, and Halverson, GP, On the initiation of a snowball Earth , in Geochemistry Geophysics Geosystems , vol. 3, 6 doi=10.1029/2001GC000219, 2002, p. 1036, DOI : 10.1029/2001GC000219 . URL consultato il 17 marzo 2010 (archiviato dall' url originale il 21 aprile 2008) . Preliminary abstract at Schrag, DP, On the initiation of a snowball Earth , su gsa.confex.com , Geological Society of America, giugno 2001. URL consultato il 17 marzo 2010 (archiviato dall' url originale il 25 aprile 2018) .
- ^ a b Benton, MJ, Twitchett, RJ, How to kill (almost) all life: the end-Permian extinction event , in Trends in Ecology & Evolution , vol. 18, n. 7, 2003, pp. 358-365, DOI : 10.1016/S0169-5347(03)00093-4 .
- ^ Dickens GR, The potential volume of oceanic methane hydrates with variable external conditions , in Organic Geochemistry , vol. 32, n. 10, 2001, pp. 1179-1193, DOI : 10.1016/S0146-6380(01)00086-9 .
- ^ Reichow MK, Saunders AD, White RV, Pringle MS, Al'Muhkhamedov AI, Medvedev AI & Kirda NP, 40 Ar/ 39 Ar Dates from the West Siberian Basin: Siberian Flood Basalt Province Doubled , in Science , vol. 296, n. 5574, 2002, pp. 1846-1849, DOI : 10.1126/science.1071671 .
- ^ Holser WT, Schoenlaub HP, Attrep Jr M, Boeckelmann K, Klein P, Magaritz M, Orth CJ, Fenninger A, Jenny C, Kralik M, Mauritsch H, Pak E, Schramm JF, Stattegger K & Schmoeller R, A unique geochemical record at the Permian/Triassic boundary , in Nature , vol. 337, n. 6202, 1989, pp. 39-44, DOI : 10.1038/337039a0 .
- ^ Dobruskina IA, Phytogeography of Eurasia during the early Triassic , in Palaeogeography, Palaeoclimatology, Palaeoecology , vol. 58, n. 1-2, 1987, pp. 75-86, DOI : 10.1016/0031-0182(87)90007-1 .
- ^ ( EN ) Newell ND, An Outline History of Tropical Organic Reefs ( PDF ), in American Museum novitates , vol. 2465, 1971, pp. 1-37. URL consultato il 3 novembre 2007 .
- ^ ( EN ) McRoberts, CA, Furrer, H., Jones, DS, Palaeoenvironmental interpretation of a Triassic– Jurassic boundary section from western Austria based on palaeoecological and geochemical data. , in Palaeogeography Palaeoclimatology Palaeoecology , vol. 136, n. 1-4, 1997, pp. 79-95, DOI : 10.1016/S0031-0182(97)00074-6 .
- ^ ( EN ) Wignall PB & Twitchett RJ, Permian–Triassic sedimentology of Jameson Land, East Greenland: Incised submarine channels in an anoxic basin , in Journal of the Geological Society , vol. 159, n. 6, 2002, pp. 691-703, DOI : 10.1144/0016-764900-120 .
- ^ a b ( EN ) R. Monastersky, Oxygen starvation decimated Permian oceans , in Science News , 25 maggio 1996. URL consultato il 20 febbraio 2010 .
- ^ a b Zhang R, Follows, MJ, Grotzinger, JP, & Marshall J, Could the Late Permian deep ocean have been anoxic? , in Paleoceanography , vol. 16, n. 3, 2001, pp. 317-329, DOI : 10.1029/2000PA000522 . URL consultato il 17 marzo 2010 (archiviato dall' url originale il 21 aprile 2008) .
- ^ Kump LR, Pavlov A, & Arthur MA, Massive release of hydrogen sulfide to the surface ocean and atmosphere during intervals of oceanic anoxia , in Geology , vol. 33, n. 5, 2005, pp. 397-400, DOI : 10.1130/G21295.1 .
- ^ Grice K, Cao C, Love GD, Bottcher ME, Twitchett RJ, Grosjean E, Summons RE, Turgeon SC, Dunning W & Yugan J, Photic Zone Euxinia During the Permian–Triassic Superanoxic Event , in Science , vol. 307, n. 5710, 2005, pp. 706-709, DOI : 10.1126/science.110432 .
- ^ Britannica Educational Publishing, The Paleozoic Era: Diversification of Plant and Animal Life , The Rosen Publishing Group, 1º agosto 2010, p. 307, ISBN 978-1-61530-196-6 . URL consultato il 20 febbraio 2012 .
{kind=link}
Bibliografia
- Michael J. Benton, When Life Nearly Died: The Greatest Mass Extinction of All Time , Thames and Hudson, 2003.
- Jess Over, (editore), Understanding Late Devonian and Permian–Triassic Biotic and Climatic Events , (Volume 20 nella serie Developments in Palaeontology and Stratigraphy (2006). The state of the inquiry into the extinction events.
- Walter C. Sweet, (editor), Permo–Triassic Events in the Eastern Tethys: Stratigraphy Classification and Relations with the Western Tethys (nella serie World and Regional Geology)
Voci correlate
Collegamenti esterni
- ( EN ) Estinzione di massa del Permiano-Triassico , su Enciclopedia Britannica , Encyclopædia Britannica, Inc.
- "The Permo–Triassic extinction" Una introduzione all'argomento.
- La causa della più grande catastrofe ambientale. Le Scienze settembre 2011 , su lescienze.it .
- Sito sui trappi siberiani. , su le.ac.uk .
- BBC2 'The Day the Earth Nearly Died' 'Il giorno che la terra quasi morì'. , su bbc.co.uk .
- PBS series Evolution : "Extinction!" video
- SpaceRef: "Big Bang in Antarctica: Killer Crater Found Under Ice" Immagini radar dalla of Ohio State University.
- Science Daily: Global warming led to atmospheric hydrogen sulfide and Permian extinction , su sciencedaily.com .
- Science Daily: Big Bang In Antarctica: Killer Crater Found Under Ice , su sciencedaily.com .
- David Morrison, "Did an Impact Trigger the Permian–Triassic Extinction?" , su nai.arc.nasa.gov (archiviato dall' url originale il 10 giugno 2011) .
- Giant Crater Found: Tied to Worst Mass Extinction Ever Robert Roy Britt (SPACE.com)
- Permian Extinction Event BBC News
- "Impact from the Deep" . Scientific American ottobre 2006.
| Controllo di autorità | LCCN ( EN ) sh2020001106 |
|---|