Fotoreceptor
Această intrare sau secțiune despre anatomie nu citează sursele necesare sau cei prezenți sunt insuficienți . |
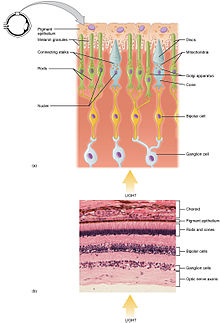
Fotoreceptorii sunt neuroni specializați care se găsesc pe retină . Lumina care ajunge în partea din spate a ochiului este „tradusă” în semnale bioelectrice care ajung la creier prin nervul optic .
Fotoreceptorii sunt de două tipuri: conuri și tije . Conurile sunt concentrate în zona centrală a retinei ( foveea ) și sunt responsabile pentru viziunea culorilor ( fotopică ) și viziunea distinctă; există cel puțin trei tipuri diferite, respectiv pentru roșu, verde și albastru (există aproximativ 6 milioane pe ochi). Lansetele, pe de altă parte, sunt mai sensibile la mișcare, sunt utilizate pentru vedere în întuneric ( scotopic ) și sunt concentrate în zona periferică a retinei. Conurile și tijele au o sensibilitate diferită la lumină datorită organizării respective a muncii. Lucrarea conurilor este individuală în sensul că fiecare dintre ele generează un impuls care este trimis la creier independent. Cu toate acestea, în cazul tijelor, câteva mii de elemente converg către un singur interneuron și impulsul care este trimis către creier iese din suma tuturor impulsurilor individuale. Tijele sunt astfel de aproximativ 4000 de ori mai sensibile la lumină decât conurile.
Trei părți pot fi identificate în structura fotoreceptorilor:
1) un segment extern: caracterizat prin structuri membrane (denumite „discuri”), pe care sunt poziționați pigmenții care reacționează la stimulul fotonilor (lumina care ajunge în „pachetele” numite cuante). Această parte este în contact cu epiteliul pigmentar , stratul cel mai exterior al retinei care conține o cantitate mare de melanină pentru a absorbi lumina care nu a fost reținută de retină. În plus, are funcția de a resinteza pigmenții vizuali și de a facilita înlocuirea discurilor.
2) segment intern: caracterizat prin prezența de organite interne precum mitocondriile , aparatul Golgi etc., esențiale pentru metabolismul celular și nucleul.
3) terminație sinaptică: permite transmiterea semnalelor de la fotoreceptor la celulele bipolare prin sinapse, adică prin transmisie biochimică între celulele nervoase (datorită moleculelor numite neurotransmițători ).
Mecanism de traducere
În celulele cu tije absorbția fotonilor are loc datorită rodopsinei , o moleculă compusă dintr-o structură proteică, opsină și un cromofor retinal. Opsina este o proteină care leagă molecula retiniană de ultimul segment transmembranar. Sosirea fotonului schimbă structura retinei prin rotația lanțului terminal conectat la opsin, trecând astfel de la forma retiniană 11-cis la forma all-trans. Molecula de rodopsină este astfel transformată mai întâi în metarodopsină I și apoi în metarodopsină II. Acesta din urmă activează subunitatea alfa a proteinei Transducin G care va activa o anumită PDE fosfodiesterază; Fosfodiesteraza va hidroliza GMPc în GMP și acest lucru va determina închiderea anumitor canale prezente în membrana fotoreceptorilor numite canale CNG a căror probabilitate de deschidere depinde, tocmai de concentrația nucleotidelor ciclice. Canalele de GNC sunt permeabile la ionii de sodiu și calciu (acesta din urmă constituie aproximativ o șeptime din curentul care intră în aceste canale); închiderea acestor canale va determina o hiperpolarizare a fotoreceptorilor care, printr-un potențial gradat, va limita exocitoza neurotransmițătorilor în terminația sinaptică.
Caracteristica receptorilor vertebratelor este că aceștia nu răspund la stimul printr-o depolarizare, ci printr-o hiperpolarizare. De fapt, în condiții normale, membrana segmentului exterior are un potențial de membrană de -40 mV mai mic decât cel al altor neuroni, ceea ce determină prezența curenților electrotoni care depolarizează și membrana presinaptică.
linkuri externe
- ( EN ) Photoreceptor , în Encyclopedia Britannica , Encyclopædia Britannica, Inc.
| Controlul autorității | Tezaur BNCF 36922 · LCCN (EN) sh85101407 · GND (DE) 4045934-2 · BNF (FR) cb16755836n (data) |
|---|