Insecta
| Insecte | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||
| Clasificare științifică | |||||||||||
| Domeniu | Eukaryota | ||||||||||
| Regatul | Animalia | ||||||||||
| Sub-regat | Eumetazoa | ||||||||||
| Ramură | Bilateria | ||||||||||
| Phylum | Arthropoda | ||||||||||
| Subfilum | Tracheata | ||||||||||
| Superclasă | Hexapoda | ||||||||||
| Clasă | Insecta Linnaeus , 1758 | ||||||||||
| Sinonime | |||||||||||
Euentomata | |||||||||||
| Subclase | |||||||||||
| |||||||||||
| ( EN ) „Pentru o aproximare aproximativă și lăsând deoparte șovinismul vertebratelor, se poate spune că, în esență, toate organismele sunt insecte” | ( IT ) „Aproximativ și lăsând deoparte șovinismul vertebratelor, putem spune că, în esență, toate organismele sunt insecte” |
| ( Robert May (1988). Câte specii există pe pământ? Science, 241 : 1441-9 ) | |
Insectele sau entomii (denumirea științifică Insecta Linnaeus , 1758 ), sunt o clasă de animale aparținând marelui filum al Arthropoda . Această clasă reprezintă cea mai mare dintre grupările de animale care populează Pământul , numărând peste un milion de specii, egale cu cinci șesimi din întregul regat animal .
Se crede că se numără printre cei mai vechi colonizatori ai țărilor emergente [1], deoarece fosilele de insecte găsite datează din Devonian . Eterogenitatea în morfologie , anatomie , biologie și etologie a conferit insectelor, de peste 300 de milioane de ani , un rol principal în colonizarea Pământului , în orice mediu în care există substanță organică , cu manifestări o capacitate remarcabilă de a concura [2] .
Insectele, în consecință, sunt organisme care, pozitiv sau negativ, au o relație strânsă cu Omul și activitățile sale, până la influențarea, mai mult sau mai puțin directă, a economiei , nutriției , obiceiurilor și sănătății.
Biodiversitatea insectelor este în scădere. [3] [4]
Morfologie
 Reprezentarea schematică a morfologiei unei insecte (femela din Orthoptera Caelifera ). |
|
|
Caracteristici generale

Insectele își datorează numele structurii metamerice a corpului [5] . Metameria insectelor este heteronomă: corpul, de fapt, este în general împărțit în trei regiuni morfologice distincte, numite cap , torace și abdomen , cărora li se asociază apendicele, cum ar fi antenele , picioarele și aripile . Regiunile morfologice sunt denumite generic tagmi .
Întregul corp este închis într-un exoschelet , format din substanțe organice care îi conferă o rezistență specifică. Există întinderi de discontinuitate în care tegumentul, din rigid, devine membranos. În acest fel corpul și apendicele sunt împărțite în unități, denumite generic segmente , sau somite (dacă se referă la corp) sau articole (dacă se referă la apendicele).
Forma, dimensiunea și aspectul general al corpului sunt extrem de variabile: multe himenoptere trichogrammatide au dimensiuni de ordinul a două zecimi de milimetru și, în orice caz, rareori depășesc un milimetru în lungime. În schimb, corpul lui Beetle Scarabeide Dynastes hercules poate ajunge la i Lungime de 17 cm [6] și molia saturnidă Attacus atlas , considerat cel mai mare fluture din lume, are o anvergură a aripilor care depășește 25 cm [7] . Formele variază foarte mult, inclusiv cele involute ale Rincoti Coccidi , unde diferențierea regiunilor morfologice și a anexelor corpului poate dispărea complet, cu excepția aparatului bucal .
Exoschelet

Susținerea și atașarea mușchilor la insecte este asigurată de un schelet extern care acționează și ca tegument . Exoscheletul este compus din mai multe straturi, organizate într-o membrană bazală , un epiteliu simplu ( epidermă ) și un strat inert, rigid și multistratificat ( cuticula ). Din acesta din urmă derivă principalele proprietăți ale exoscheletului; macromoleculele organice participă la compoziția sa care conferă rigiditate, rezistență, impermeabilitate, elasticitate, flexibilitate. Grosimea cuticulei nu este uniformă, ci este întreruptă la articulații, înlocuită de un țesut membranos și la suturi . Acestea sunt linii de rezistență mai mică care separă porțiunile exoscheletului real ( sclerite ). Zonele membranoase garantează flexibilitate și mișcare, suturile oferă o rezistență mai mică, ceea ce permite desfășurarea costumului de neopren .
Suprafața exoscheletului este adesea presărată cu fire de păr, peri, solzi și sculpturi, uneori esențiale în scopuri taxonomice .
Șefu
 Șef al ortopterelor . | a: antenă |
Capa este regiunea compusă din primele șase somite care se îmbină într-o structură în care se pierde metameria originală. Primii trei somite formează procefalia , următorii trei gnatocefalia [8] . Exoscheletul formează o capsulă care are două deschideri, peristomul și foramenul magnum . Primul definește deschiderea gurii, al doilea conectează capul la piept.
Structurile cefalice externe au două funcții principale: percepția stimulilor senzoriali și prima fază a hrănirii (luarea, prelucrarea și ingerarea alimentelor). Aceste structuri includ în general o pereche de antene , cei doi ochi compuși , 2 sau 3 ocelli și, în cele din urmă, apendicele aparatului bucal .
Antenele sunt organe cu o funcție senzorială predominantă, de formă și dezvoltare variate, constând dintr-un număr variabil de articole mai mult sau mai puțin diferențiate, numite antenomeri . Ochii compuși sunt structuri complexe atribuite percepției vizuale, formate prin agregarea unităților elementare numite ommatidia . Ocelli sunt ochi rudimentari, absenți în unele grupuri sistematice, esențiali pentru percepția orientării la insectele zburătoare [9] [10] .
Gnatitele sunt anexe care se articulează la peristom. Sunt reprezentate, în sens dorso-ventral, de o pereche de mandibule , o pereche de maxilare și buza inferioară ( labium ). Anterior, deschiderea bucală este delimitată de buza superioară ( labrum ) care, contribuind la formarea aparatului bucal, este de fapt derivată din primul somit al procefalului. Cavitatea bucală, delimitată de apendicele bucale, este împărțită în două părți, una dorsală și una ventrală, de prefaringe . Palpi , organele multi-articulate responsabile de percepția stimulilor senzoriali, sunt în general inserate pe maxilare și pe buza inferioară.
Aparatul tipic de gură este cel de mestecat, potrivit pentru a lua alimente solide și a le mărunți. Cu toate acestea, multe grupuri sistematice prezintă modificări anatomice, morfologice și funcționale care îl adaptează la alte funcții. Vorbim apoi despre un aparat de supt înțepător, de supt, de lins.
Cufăr
 Vedere laterală a toracelui unui ortopter . | a: gât |
Toracele derivă din combinația a trei somite și este în principal sediul organelor motorii. Este împărțit în pro- , mezo- și metatorax . Exoscheletul fiecărui segment poate fi urmărit înapoi la un inel format prin organizarea a patru zone:
- o dorsală, numită tergo, tergite sau Noto;
- două laterale, numite pleură ;
- o ventrală, numită stern sau sternită .
În terminologia este folosit pentru a specifica în același timp , segmentul toracic și zona la care se referă: de exemplu, tergites celor trei segmente sunt numite respectiv pronotum, mesonotum, metanotum; denumiri similare sunt folosite pentru a indica sternitele, pleura și, adesea, apendicele și scleritele individuale.
Morfologia tergitei, pleurei și sternului poate fi relativ simplă sau complicată de prezența suturilor, în special în al doilea și al treilea segment al formelor înaripate. Diferențe marcate există și în dezvoltarea relativă a celor trei segmente: formele apterice au un protorax destul de dezvoltat și o omogenitate substanțială a celor trei segmente; formele înaripate, în schimb, au un protorax slab dezvoltat, iar celelalte segmente toracice foarte dezvoltate în raport cu rolul jucat de aripile respective. Structurile mai complexe se găsesc în Diptera , în care mezotoraxul are o dezvoltare preponderentă în comparație cu celelalte segmente, și în Himenoptera Apocritică , unde o parte din uritul I participă, de asemenea, la constituirea toracelui, formând al patrulea segment toracic. ( propodeum ).
În generalitatea insectelor există o pereche de picioare pentru fiecare segment toracic, care în mod secundar se poate reduce sau dispărea. Prezența a trei perechi de picioare toracice se găsește în general și în formele juvenile. La unele larve există și organe abdominale de locomoție ( pseudo-labe ), la altele pot fi atrofice sau lipsesc cu totul. Sunt anexe libere și multi-articulate, formate din coxa (sau șold ), trohanter , femur , tars , pretars și unghii . Tarsul este la rândul său împărțit în mai multe articole numite tarsomer și pretarul este ascuns în ultimul tarsomer. Unghiile se articulează pe pretar, ceea ce permite aderența la suprafețe aspre și la alte organe cu forme și funcții specifice. Funcția primară a picioarelor este cea locomotorie; în acest caz picioarele sunt numite cursoare sau ambulatorii . Adaptările morfologice și anatomice vizează adesea funcții precum înotul, prădarea, săpatul, săriturile.
Aripile sunt prezente, în stadiul adult și ca personaj primar, la toate insectele, cu excepția Apterygota . Meiotterismul secundar este frecvent, cu reducerea evolutivă a aripilor până la dispariția completă ( apoteza ). Sunt purtate de mezotorax și metatorax. Din punct de vedere morfologic, acestea sunt expansiuni laterale ale exoscheletului toracelui și constau structural din două lamine suprapuse: una dorsală, în continuitate cu tergitul, una ventrală, în continuitate cu pleura. La majoritatea insectelor continuitatea este întreruptă de un sistem de articulații compus din trei sclerite pterice și două sclerite toracice diferențiate de pleure. Aripile insectelor sunt lipsite de mușchi intrinseci și mișcarea se datorează acțiunii mușchilor localizați în torace. Aripa este traversată de conducte, numite coaste sau vene , în care trec hemolimfa , nervii și traheele . Cursul nervurilor este un element important al determinării taxonomice în unele ordine.
Morfologia aripilor variază foarte mult în funcție de grupul sistematic. În general sunt membranosi, uneori sunt mai mult sau mai puțin sclerificate. Anteriorul are adesea o dezvoltare mai mare și pot fi prezente organe de conectare specifice pentru sincronizarea mișcării. Suprafața aripii este împărțită în trei regiuni, respectiv numite remiges , anal și jugal ; primul are cea mai mare dezvoltare și cea mai mare forță. Funcția principală este aceea de zbor, cu toate acestea adaptările pot modifica această funcție până când se pierde complet în favoarea altor scopuri, transformând aripile în organe de protecție, reglare a echilibrului etc.
Abdomen
 Abdomenul unei ortoptere feminine. | la: tars |
Abdomenul este regiunea morfologică în care se află organele de reproducere . De o consistență mai delicată decât toracele, este compus din 11 segmente, numite urite , dar urita XI [11] este atrofică sau rudimentară. În orice caz, în grupurile sistematice mai evoluate, numărul de urite tinde să scadă datorită involuției sau modificării acestora.
Fiecare urită are o formă care poate fi urmărită înapoi la cea a unui inel în care se disting patru zone: un dorsal ( urotergus ), un ventral ( urosterno ) și două laterale membranoase, uneori sclerificate prin expansiunea scleritei ventrale sau dorsale. Uritele sunt articulate între ele prin membrana intersegmentală , o structură elastică și flexibilă, care permite regiunii să se dilate și să aibă o anumită mobilitate.
Apendicele abdominale nu sunt în general foarte evidente în comparație cu cele toracice și cefalice. La majoritatea insectelor sunt prezente organele genitale externe ( gonapofiză sau armături genitale): la bărbați este prezent organul copulator , format din expansiunile urosternului IX; la femele ovipozitorul , format din expansiunile urosternului VIII și IX. De asemenea, ovipozitorul poate atinge o dezvoltare remarcabilă în unele grupuri sistematice; uneori se transformă într-o înțepătură și devine un instrument de ofensă și apărare; la altele poate fi complet absent și funcția este îndeplinită de alte structuri care iau numele de ovipozitor de substituție .
Alte anexe obișnuite la multe insecte sunt cercii , introduși pe laturile ultimului urit. Rudimentare sau absente în ordinele mai puțin evoluate, în unele grupuri sistematice pot ajunge la o dezvoltare notabilă. În larve există adesea procese care îndeplinesc diverse funcții. Cele mai frecvente sunt pseudo-picioarele larvelor eruciforme .
Anatomie
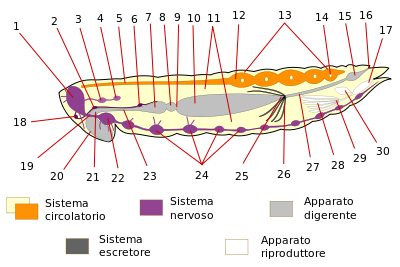 Reprezentarea schematică a anatomiei unei insecte feminine (sistemele respiratorii, secretorii și musculare nu sunt prezentate) |
|
|
Comparativ cu vertebratele , insectele și artropodele inversează luxația sistemelor axiale: sistemul circulator și sistemul nervos periferic își asumă respectiv o poziție dorsală și ventrală, în timp ce sistemul digestiv este deplasat într-o poziție centrală. În timp ce menținem unele analogii, structura anatomică diferă semnificativ de cea a vertebratelor, chiar dacă avem tendința de a împrumuta terminologia specifică a organelor și sistemelor pentru relevanță.
Endoschelet
Endoscheletul este o extensie internă a exoscheletului care favorizează atașarea mușchilor și sprijinul organelor. Este compus dintr-un set de procese interne, de diverse forme și dezvoltare. În șef este localizat tentoriul, cea mai complexă formațiune, prezentă în aproape toate insectele. Susține creierul și esofagul și mușchii extrinseci ai antenelor și cei ai gurii sunt conectați la acesta. În torace există, pentru fiecare segment, câte o pereche de fragme (dorsale), una de apodeme (laterale) și furche (neuniformă și ventrală). În cele din urmă, în abdomen există apodeme dorsale și ventrale de dezvoltare redusă.
Sistem muscular
Mușchii insectelor sunt aproape toți de tip striat; acestea sunt atașate la cuticula exoscheletului sau endoscheletului prin intermediul fibrilelor necontractile ( tonofibrile ), asemănătoare cu tendoanele vertebratelor . Numărul lor este mare, de ordinul a mii [12] . Acestea sunt împărțite în schelet și visceral : primele au nume derivate din calea lor sau funcția lor și sunt în general uniforme și simetrice. Ei sunt responsabili de mișcările anexelor și regiunilor morfologice ale corpului, prin urmare sunt numite și somatice . Musculatura viscerală este asociată cu organele interne și este formată doar parțial din fibre musculare netede.
Sistem digestiv
Sistemul digestiv al insectelor este un tub care începe de la deschiderea gurii și se termină în cel anal. Este împărțit în trei secțiuni distincte: stomodeo , mezenter , proctodeum , respectiv numit și intestin anterior, mediu și posterior. Calea poate fi mai mult sau mai puțin sinuoasă în funcție de complexitatea anatomo-fiziologică a speciei. În cele mai primitive forme este un canal longitudinal care traversează întregul corp, în timp ce specializările particulare sunt la originea complicațiilor structurale. Trece cele trei regiuni morfologice plasându-se deasupra lanțului ganglionar ventral și sub vasul dorsal. Terminologia adoptată pentru a distinge diferitele secțiuni este adesea împrumutată de cea a vertebratelor, dar diferențele structurale și fiziologice profunde nu implică nicio omologie.
Stomodeo îndeplinește funcțiile de ingestie și prima prelucrare a alimentelor și, la unele insecte, de depozitare temporară. Începe de la deschiderea gurii și se termină la valva inimii unde curge în mezenter. Procedând în direcția cefalo-caudală, este împărțit în următoarele secțiuni: cavitate bucală , faringe , esofag , gizzard , acesta din urmă dezvoltându-se mai presus de toate la insecte cu piese bucale de mestecat. Stomodeo are uneori un diverticul, ingluvie , care poate atinge dimensiuni precum să ocupe o parte a abdomenului. Funcțiile ingluvie au specificități, cum ar fi justificarea unor denumiri particulare (de exemplu, pungă de miere la albine , stomac social la furnici , pungă de sânge la țânțari ).
Mezenterul este locul unde are loc digestia efectivă, cu demolarea enzimatică a macromoleculelor și absorbția nutrienților. Bolusul alimentar este înconjurat de o membrană ( membrană peritrofică ) produsă de celulele epiteliale ale mezenterului și care formează o veziculă în cadrul căreia au loc procesele de demolare. Membrana în sine acționează ca un filtru de dializă , prevenind contactul direct al bolusului și al enzimelor digestive cu peretele mezenterului.
Proctodeo este partea din spate și este responsabil în primul rând de reabsorbția apei și de eliminarea părților nedigerate și a cataboliților. În secțiunea inițială este legată de aparatul excretor, deoarece tuburile Malpighiene converg acolo. Începe de la valva pilorică , dar în unele grupuri sistematice mezenterul este orb și, prin urmare, există o ruptură în continuitatea tractului digestiv. În sens antero-posterior, proctodeo este împărțit în secțiuni care au nume împrumutate din sistemul digestiv al vertebratelor; în ordinea distingem ileonul , colonul , rectul .
Natura histologică a stomodului și a proctodeului este ectodermică , cea a mezenterului este probabil entodermică [13] [14] .
Pereții stomodeului și proctodeului sunt compuși, din interior spre exterior, de un strat cuticular, numit intima , de un strat epitelial, de o membrană bazală, de o tunică musculară formată dintr-un fascicul intern de mușchi longitudinali și unul extern a mușchilor circulari și, în cele din urmă, a unei tunici peritoneale de țesut conjunctiv . Tunica musculară include, în stomodeo, un pachet intern de mușchi longitudinali și unul extern de mușchi circulari. În proctodeu există trei fascicule musculare: unul intermediar de mușchi longitudinali între două straturi de fibre circulare.
Mezenterul, care se desfășoară din interior spre exterior, este compus din membrana peritrofică, un epiteliu simplu, compus din celule care au o dezvoltare diferită, o membrană bazală, o tunică musculară compusă dintr-un pachet intern de mușchi circulari și unul extern de mușchii longitudinali și, în cele din urmă, tunica intestinală.
Structura tipică a canalului digestiv poate suferi modificări profunde în funcție de grupurile sistematice. Una dintre cele mai semnificative este prezența camerei de filtrare în aproape toate Rincoti Omotteri . Acest organ are funcții de dializă și bypass : pentru a evita diluarea excesivă a sucurilor digestive ale mezenterului, camera filtrantă separă o parte substanțială din lichid și zaharuri de macromoleculele lipidice și proteice . Prima fază este deviată în proctodeum, în timp ce macromoleculele vor urma calea normală, intrând în mezenter. Această adaptare este tipică pentru insectele producătoare de miere , un excrement lichid cu un conținut ridicat de zahăr, de mare importanță ecologică [15] .
Sistemul respirator

Transportul oxigenului până la celule nu este încredințat circulației sanguine [16] , prin urmare, insectele prezintă diferențe marcate în ceea ce privește sistemele circulator și respirator al vertebratelor.
Sistemul respirator al insectelor preia parțial structura metamerică a corpului lor, dar în interior se rezolvă într-o rețea complexă și densă de conducte care transportă oxigenul către celule. Stigmele , traheele și traheolul formează aparatul. Stigmele sau spiraculele traheale sunt deschideri, în număr de 1-10 perechi, situate pe laturile unuia sau mai multor segmente toracice și abdominale. Traheele sunt tubuli derivati dintr-o invaginare a tegumentului, din care mentin aceeasi structura. Trei ramuri traheale pornesc de la fiecare stigmat, conectate între ele și cu cele ale segmentelor adiacente, formând o rețea anastomozată în ansamblu. Traheele se ramifică la rândul lor în traheole , de calibru inferior, care formează o rețea de transport capilar. Traheolele mai mici au un diametru de ordinul câtorva microni .
La multe insecte traheele se pot dilata formând saci de aer , a căror funcție este de a constitui rezerve de aer sau de a acționa ca organe hidrostatice sau aerostatice [17] [18] .
Sistem circulator
Sistemul circulator este deschis: circulația umorală nu are loc în interiorul vaselor, ci într-o cavitate, numită hemocel sau lacunom , în contact cu organele și țesuturile din toate regiunile corpului, inclusiv coastele aripilor. Are loc doar parțial în interiorul unui vas, care are funcțiile unei inimi . Umorul care curge în hemocel, hemolimfa , îndeplinește simultan funcțiile limfei și sângelui vertebratelor , cu excepția transportului de oxigen , care la insecte este încredințat sistemului respirator [16] . Hemolimfa, pe de altă parte, este responsabilă pentru transportul substanțelor nutritive și cataboliți și pentru apărarea imună .
Hemocelul este împărțit de două diafragme longitudinale în trei cavități intercomunicante. Formate de țesut conjunctiv și muscular, au o mișcare peristaltică care favorizează circulația spre zona caudală și intrarea în partea posterioară a vasului dorsal pulsatoriu. Vasul dorsal diferă în două secțiuni, una posterioară ( inimă ) și una anterioară ( aorta ). Inima este împărțită în camere intercomunicante ( ventriculită ), fiecare prevăzută cu două valve ( ostioli ) care reglează continuitatea cu hemocelul. Sistemul muscular asociat vasului dorsal impresionează pulsațiile care împing sângele spre regiunea cefalică. Alte organe pulsatorii accesorii se află în anexe și favorizează circulația în organe care ar oferi o rezistență mai mare.
Hemolimfa insectelor diferă considerabil de sângele vertebratelor ; este compus dintr-o fracție lichidă ( plasmă ) și o fracțiune celulară, formată din hemocite . Orice pigmenți , de obicei galbeni sau verzi, nu au funcții respiratorii.
Sistem nervos

P: protocerebro
D: deutocerebro
T: polizor
G: gnatocerebro
CV: lanț ganglionar ventral
În analogie cu sistemul nervos al vertebratelor, insectele disting un sistem central, periferic și visceral, acesta din urmă cu funcții neurovegetative.
Neuronii sunt de trei tipuri: senzoriale , motorii și asociative . Primii sunt asociați cu receptori senzoriali, sunt bipolari și transmit impulsuri de la periferie la ganglioni. Acestea din urmă au corpul localizat în ganglioni, sunt unipolare și transmit impulsuri către sistemul muscular. Terțele, situate și în ganglioni, sunt multipolare și îndeplinesc funcții asociative. Transmiterea semnalelor urmează aceleași mecanisme ca și la vertebrate: de-a lungul axonilor sub forma unui impuls electric, prin alterarea potențialului membranei și în sinapsele cu emisia unui mediator chimic , acetilcolina . Multe insecticide , care acționează ca inhibitori ai acetilcolinesterazei , au, de asemenea, un efect neurotoxic asupra vertebratelor.
Sistemul nervos central are o organizare metameră: este format dintr-un lanț dublu de ganglioni, localizați ventral sub canalul digestiv, cu o pereche de ganglioni pentru fiecare segment. Ganglionii sunt conectați între ei prin fibre nervoase transversale și longitudinale, care dispar odată cu eventuala fuziune. Structura metamerică dispare în cap, odată cu fuziunea ganglionilor cefalici în două mase. Primul, numit cerebro , cerebr sau ganglion supraesofagian , este situat deasupra stomodului. Al doilea, numit gnatocerebro sau ganglion subesofagian , este poziționat sub stomodiu. Cele două conexiuni care leagă cerebro de gnatocerebro formează un inel ( brâu parastomodeal sau periesofagian ) traversat de esofag. Cele două mase ganglionare inervează organele și apendicele capului. Dal cerebro partono, inoltre, il simpatico dorsale e il sistema neuroendocrino cardio-aortico .
Il sistema nervoso viscerale costituisce il complesso dell'apparato neurovegetativo e mantiene una propria autonomia rispetto al centrale, pur essendovi collegato. Ad esso compete l'innervazione di organi interni ed è composto da tre distinti sistemi: il simpatico dorsale o stomatogastrico , il simpatico ventrale , ed il simpatico caudale . Il primo, impari, si localizza sotto il cerebro e nella parte dorsale dello stomodeo e innerva il tratto iniziale dell'apparato digerente. Il secondo parte dai gangli della catena ventrale e innerva le trachee e gli stigmi. Il terzo, infine, parte dall'ultima coppia della catena ventrale e innerva il proctodeo e gli organi genitali.
Il sistema nervoso periferico comprende gli assoni dei neuroni motori, che innervano la muscolatura striata, ei neuroni sensoriali associati ai recettori. Si sviluppa in tutte le parti del corpo ed è associato ai gangli del sistema nervoso centrale.
Apparato escretore
L' apparato escretore provvede all'eliminazione delle sostanze di rifiuto (cataboliti). È composto da organi localizzati ( tubi malpighiani ) e diffusi ( nefriti o nefrociti ). A questi si aggiungono tessuti e altri organi che svolgono anche funzioni escretorie secondarie.
I tubi malpighiani sono reni costituiti da tubuli sottili, a fondo cieco, che confluiscono nel proctodeo , subito dopo la valvola pilorica. Assenti negli Afidi , negli altri insetti sono presenti in numero variabile da poche unità ad oltre 200. Fluttuano nell' emocele oppure aderiscono al proctodeo. La funzione è quella di filtrare i prodotti del metabolismo proteico ( ammoniaca , urea , acido urico ) e regolare gli equilibri salini , sottraendo le sostanze di rifiuto dall' emolinfa [19] . Secondariamente possono ospitare microrganismi simbionti , produrre seta , sostanze collanti, ecc. [19] .
I nefriti sono gruppi sparsi di cellule, a volte isolate, associati allo stomodeo , alle ghiandole salivari e al vaso dorsale. La loro funzione è quella di regolare il pH dell'emolinfa e agire come organi escretori intermedi, trasformando i cataboliti in forme che saranno poi eliminate dai tubi malpighiani [20] .
Apparato secretore

Gli Insetti dispongono di un apparato ghiandolare complesso, distinto in un sistema esocrino, composto da ghiandole a secrezione esterna, e in uno endocrino, composto da ghiandole a secrezione interna.
Il sistema endocrino è formato da cellule e organi che hanno relazione con il sistema nervoso. Regola la biologia dell'Insetto e raggiunge livelli di complessità organizzativa paragonabili a quello degli animali superiori. Una particolarità che differenzia gli Insetti dai Vertebrati consiste nell'assenza di funzioni di secrezione endocrina nell'apparato riproduttore. Le ghiandole endocrine d'importanza basilare si identificano in quattro strutture: il sistema endocrino cerebrale , i corpi cardiaci e allati e le ghiandole protoraciche . La funzione primaria svolta da queste ghiandole è la regolazione dellosviluppo postembrionale , ma singolarmente queste strutture controllano anche altri processi biologici.
Il sistema endocrino cerebrale è costituito da cellule nervose del cerebro, che hanno perso la capacità di trasmettere impulsi nervosi diventando ghiandole neuroendocrine. L'attività principale si ha nello stadio giovanile, con la produzione dell' ormone cerebrale , trasportato dagli assoni ai corpi cardiaci . Questi accumulano l'ormone, lo modificano e lo mettono in circolo nell'emolinfa. Funzioni secondarie dei corpi cardiaci consistono nel rilascio di ormoni che regolano il funzionamento di alcuni organi e del sistema muscolare. I corpi allati sono a loro volta deputati, dietro stimolo dell'ormone cerebrale, alla produzione della neotenina , responsabile della persistenza dei caratteri giovanili. Le ghiandole protoraciche sono localizzate ventralmente, in genere fra il capo e il torace. La loro funzione primaria consiste nel rilascio, dietro stimolo dell'ormone cerebrale, dell' ecdisone , che induce la muta .
Il sistema esocrino è composto da ghiandole uni- o pluricellulari. Queste ultime sono a loro volta semplici o composte, le prime con un unico dotto escretore comune a tutte le cellule, le altre con un dotto escretore per ogni cellula confluente un serbatoio comune. Altre classificazioni possono riguardare la posizione e la distribuzione nel corpo.
Per quanto riguarda la natura del secreto, nella classe vi è una grande varietà di ghiandole, molte delle quali sono tuttavia specifiche di uno o più gruppi sistematici. Ghiandole che svolgono la stessa funzione possono inoltre differire per localizzazione, struttura e sviluppo in funzione delle varie categorie sistematiche. Una trattazione dell'anatomia e della fisiologia del sistema esocrino non può prescindere da una semplificazione riduttiva, data l'estrema eterogeneità. Per gli approfondimenti si rimanda perciò ai singoli gruppi sistematici.
Fra i sistemi ghiandolari di maggiore frequenza si citano i seguenti:
- Ghiandole accessorie dell'apparato genitale . Presenti in entrambi i sessi, secernono sostanze che in genere sono coadiuvanti della riproduzione: il secreto dei maschi ha funzioni trofiche nei confronti degli spermatozoi , quello delle femmine interviene, in genere, nella dinamica dell'ovideposizione. Non mancano funzioni specifiche estranee alla riproduzione (es. le ghiandole velenifere degli Imenotteri Aculeati ).
- Ghiandole a feromoni . Presenti in vari ordini e, in genere, localizzate nell'addome, secernono sostanze volatili attive a basse concentrazioni, percepite quasi sempre solo da individui della stessa specie. I feromoni hanno per lo più una funzione di comunicazione chimica nella vita relazionale e sono coinvolti nella riproduzione, nelle interrelazioni all'interno di comunità, nelle dinamiche di competizione intraspecifica.
- Ghiandole ceripare . Frequenti fra gli Imenotteri , i Rincoti Omotteri e alcuni Coleotteri , sono in genere diffuse nel tegumento e secernono la cera . Il secreto è usato per vari scopi, fra cui è frequente la protezione del corpo.
- Ghiandole odorifere . Presenti in genere nella parte dorsale del corpo, si rinvengono in vari ordini ( Rincoti , Coleotteri, Ditteri , Imenotteri, Lepidotteri , Neurotteri ). Il secreto è composto da sostanze volatili che possono avere un effetto repulsivo o, al contrario, attrattivo. Nel primo caso ha funzioni difensive, nel secondo induce spesso l'instaurazione di rapporti di simbiosi .
- Ghiandole salivari . Presenti nella zona cefalica della generalità degli Insetti, sono associate alle appendici boccali. Emettono succhi digestivi contenenti spesso anche sostanze con azione specifica in determinati contesti (anticoagulanti, declorofillizzanti, ecc.)
- Ghiandole sericipare . Presenti in molti ordini, mostrano una notevole varietà nella localizzazione e nella struttura. Secernono la seta , utilizzata, secondo i casi, per la costruzione di bozzoli , nidi, ooteche , capsule protettive del corpo, ecc.
Apparato riproduttore

Negli Insetti, l' ermafroditismo è stato accertato solo in Icerya purchasi e in alcuni Ditteri [21] , perciò l'apparato riproduttore si differenzia in due sessi distinti a partire da strutture embrionali omologhe.
Le gonadi , di origine mesodermica, sono la sede di svolgimento della gametogenesi . Si differenziano nei testicoli (maschio) e negli ovari (femmina) e sono composte da elementi tubulari ( testicoliti e ovarioli ) confluenti nei gonodotti pari; in entrambi i sessi le fasi della gametogenesi si succedono lungo il tubulo in ordine distale-prossimale. Alle gonadi seguono i gonodotti pari , condotti di origine ectodermica che confluiscono in un gonodotto comune , impari. Il gonodotto comune, nel maschio, è detto dotto eiaculatore e mantiene la fisionomia di un condotto escretore che comunica con l'organo copulatore. Nella femmina si differenzia nella vagina e comunica in genere con l'ovopositore; in alcuni gruppi sistematici può presentare un diverticolo dotato di apertura indipendente, la borsa copulatrice , usata per la copula. Un altro organo differenziato nella femmina è la spermateca , una vescicola a fondo cieco collegata alla vagina, in cui vengono mantenuti gli spermatozoi nell'intervallo di tempo che intercorre fra l'accoppiamento e la fecondazione.
In entrambi i sessi, l'apparato dispone di ghiandole accessorie , che riversano i secreti nel tratto prossimale del gonodotto comune. I secreti hanno funzioni diverse, secondo i sessi e le specie, e possono coesistere differenti tipi ghiandolari nello stesso individuo. Il secreto del maschio svolge in genere la funzione di lubrificante e di nutrizione degli spermatozoi. Nella femmina la natura e la funzione del secreto dipendono essenzialmente dal tipo ghiandolare. Le ghiandole più comuni sono dette colleteriche e il loro secreto è usato come coadiuvante dell'ovideposizione, come collante per le uova, per la costruzione di ooteche, ecc. Altre ghiandole, dette spermofile , secernono un fluido nutritivo per gli spermatozoi conservati nella spermateca. Negli Imenotteri Apocriti sono presenti anche le ghiandole velenifere : il loro secreto, emesso con l'ovopositore o con l'aculeo, ha un'azione associata all'ovideposizione oppure integra la funzione secondaria di strumento di offesa e difesa nelle femmine sterili.
Gli organi genitali esterni si differenziano in genere come processi degli urosterni costituendo l'armatura genitale, importante elemento di determinazione tassonomica. Nel maschio è differenziata dal IX urosterno e forma l' organo copulatore o edeago , impiegato per l'accoppiamento. L'edeago può mancare negli insetti primitivi: in questo caso gli spermatozoi sono rilasciati all'esterno in capsule, dette spermatofori , che saranno poi prelevate e introdotte dalla femmina. Nella femmina, l'armatura genitale, detta ovopositore , è differenziata dagli urosterni VIII e IX ed è generalmente composta da tre coppie di valve, talvolta di lunghezza eccezionale. È utilizzato come organo per la deposizione delle uova e può anche perforare tessuti vegetali o animali. In diversi Insetti, l'ovopositore può mancare del tutto oppure è sostituito funzionalmente da adattamenti degli ultimi uriti, che prendono il nome di ovopositore di sostituzione . Negli Imenotteri Aculeati l'ovopositore subisce un adattamento anatomico e funzionale che lo trasforma in strumento di offesa e difesa; in questo caso prende il nome di aculeo o pungiglione .
Biologia

La lunga storia evolutiva, l'elevato numero di specie, la capacità di adattamento a condizioni di vita anche estreme, fanno degli Insetti un raggruppamento sistematico dalla biologia eterogenea. Una trattazione organica che copra i variegati aspetti, dalla fisiologia al ciclo biologico, dall'alimentazione alla riproduzione, dalle relazioni all'etologia in generale, non può che essere sommaria e generica. Si rimanda perciò alle specifiche sezioni e alla trattazione dei singoli gruppi sistematici per gli opportuni approfondimenti.
Evoluzione
L' evoluzione degli insetti data almeno dal periodo Devoniano , epoca a cui risale il più antico fossile di insetto rinvenuto, Rhyniognatha hirsti , la cui età è stimata in circa 400 milioni di anni [22] . Durante la storia della terra, il clima è cambiato numerose volte, influenzando la diversità degli insetti. Gli Pterigoti subirono una massiccia estinzione durante il Carbonifero , mentre gli Endopterigoti subirono un'altra grave estinzione nel Permiano . I sopravvissuti a queste estinzioni di massa sono evoluti durante il Triassico in quelli che sono essenzialmente gli insetti moderni. Molte famiglie di insetti attuali apparvero nel Giurassico e durante il Cretaceo numerosi generi si svilupparono o apparvero. Si pensa che nel Terziario già esistesse la maggior parte dei generi di insetti moderni; molti degli insetti racchiusi nell' ambra fossile appartengono di fatto a generi contemporanei.
Ciclo biologico
Il ciclo biologico di un Insetto può durare, secondo la specie, da pochi giorni a diversi anni e, spesso, il suo svolgimento è in stretta relazione con fattori ambientali di natura climatica (in particolare la temperatura ) e nutrizionali. Ciò fa sì che gli Insetti possano adattarsi a svariati ambienti, comprese le regioni più fredde della Terra, ricorrendo ad accorgimenti biologici quali lo svernamento in stato di diapausa e la migrazione stagionale.
Il ciclo biologico si dice monovoltino o univoltino se compie una sola generazione l'anno, polivoltino se compie più generazioni. Il numero delle generazioni non è correlato alla lunghezza del ciclo, in quanto in una popolazione possono coesistere differenti stadi di sviluppo con sovrapposizione di più generazioni (es. Afidi ). Condizioni di elevata specificità possono sincronizzare il ciclo con quello di una specie vegetale o animale da cui l'Insetto dipende per la sua nutrizione. In questi casi si può avere la concentrazione e la sincronizzazione degli eventi riproduttivi e dello sviluppo, con generazioni ben definite (es. la Tignola dell'olivo ).
Riproduzione

La riproduzione avviene in genere per via sessuale ( anfigonia ), con l'accoppiamento fra sessi distinti e la fecondazione delle uova da parte degli spermatozoi . I meccanismi che regolano l'anfigonia variano da specie a specie e, talvolta, hanno livelli di complessità tale da condizionare la dinamica di un'intera comunità ( Insetti sociali ). All'anfigonia si accompagnano altre forme di riproduzione non sessuale, che spesso si riflettono sulla dinamica di popolazione e, in qualche specie, prevalgono sulla riproduzione sessuale. In questi casi ricorre spesso la partenogenesi , riscontrata in quasi tutti gli ordini in varie forme. Un caso particolare di partenogenesi è la pedogenesi , accertata in alcuni Ditteri Cecidomiidi e nel Coleottero Micromalthus debilis : la larva porta a maturazione gli ovari e genera individui simili che si sviluppano a sue spese, uccidendola [23] [24] . Un altro tipo di riproduzione non sessuale è la poliembrionia , che consiste nell'origine di un numero indefinito di larve dalla segmentazione di un solo uovo, riscontrata in alcuni Imenotteri Terebranti [24] .
La determinazione del sesso non segue un unico schema. Nella generalità delle specie, il sesso è determinato dalla presenza di un cromosoma sessuale X in doppia dose in un sesso e in singola dose, o abbinato ad un cromosoma Y, nel sesso opposto. Il sesso eterogametico è quello maschile, ad eccezione dei Lepidotteri e dei Tricotteri . In alcuni ordini il sesso è invece determinato dal rapporto fra anfigonia e partenogenesi aploide: dalla fecondazione si originano le femmine, dalla partenogenesi i maschi.
La maggior parte degli Insetti è ovipara , ma si riscontrano anche la viviparità e l' ovoviviparità . Questi differenti comportamenti possono coesistere anche nell'ambito di una stessa specie o, addirittura, di uno stesso individuo [25] .
Sviluppo postembrionale

Lo sviluppo postembrionale degli Insetti si fonda sulla metamorfosi , sequenza discontinua di trasformazioni fisiche che, a partire dall'uovo, porta alla formazione dell'insetto adulto.
Negli Insetti e, più in generale, negli Artropodi, si deve distinguere l'accrescimento ponderale (incremento del peso), che procede in modo continuo, dall'accrescimento lineare (incremento delle dimensioni), che è invece discontinuo. A causa della rigidità dell'esoscheletro, l'aumento di peso in un insetto giovane non è accompagnato da un aumento di dimensioni: ad un certo punto, la vecchia cuticola ( exuvia ) si lacera e l'individuo ne fuoriesce con una nuova, in grado di contenere l'aumento di dimensione. Il fenomeno è detto muta .
Alla muta può accompagnarsi anche la metamorfosi, ossia una trasformazione fisica che non si limita solo all'aumento delle dimensioni. Accrescimento e metamorfosi sono regolati da meccanismi neurormonali che si riassumono nell'interazione tra due ormoni , l' ecdisone e la neotenina . L'ecdisone stimola la genesi della nuova cuticola, perciò è il segnale biologico che avvia una nuova muta. La neotenina, invece, inibisce la comparsa dei caratteri dell'adulto, che restano latenti sotto forma di abbozzi. L'ecdisone stimola la muta e la metamorfosi, ma in presenza di neotenina la metamorfosi viene inibita.
La classificazione dei tipi di metamorfosi verte su schemi per i quali non c'è uniformità di orientamento fra gli Autori. Le fasi dello sviluppo ontogenetico possono infatti essere interpretate sotto differenti aspetti e la terminologia non sempre è usata in modo inequivocabile. Per ulteriori approfondimenti si rimanda ai dettagli descritti nella voce di approfondimento.
Ad una prima analisi, si distinguono due tipi fondamentali di metamorfosi, indicati con i termini di metamorfosi completa e incompleta . L'elemento morfologico che distingue i due tipi è il modo in cui si formano, eventualmente, gli abbozzi alari.
Nella metamorfosi completa, che si identifica con l' olometabolia , lo stadio giovanile è detto larva e differisce notevolmente dall'adulto (detto anche immagine ). Questa differenza è alla base di profonde modificazioni, che si attuano durante la ninfosi , nello stadio di pupa , spesso preceduto da una fase brevissima ( eopupa ): l'insetto subisce una drastica ricostruzione della sua struttura morfologica e anatomica, ad eccezione del sistema nervoso e dell'apparato circolatorio. L'entità delle modifiche è tale da richiedere, in questa fase, lo stato di immobilità dell'insetto. Gli abbozzi alari sono interni e compaiono solo nello stadio di pupa. Questo tipo di metamorfosi è proprio degli Insetti più evoluti, alati o secondariamente atteri, i cui ordini sono compresi nella coorte degli Endopterigoti . Nelle forme più primitive di metamorfosi completa, la pupa è mobile. In realtà la pupa mobile è un adulto in fase farata , stadio di transizione, successivo all'ultima muta, durante il quale l'adulto, pur essendo completamente formato, non ha ancora abbandonato l' exuvia [26] .
Nella metamorfosi incompleta, che si identifica con l' eterometabolia , proprio degli Insetti alati, o secondariamente atteri, meno evoluti, appartenenti alla coorte degli Esopterigoti , lo stadio giovanile è detto neanide e differisce dall'adulto solo per le minori dimensioni, l'assenza dei caratteri sessuali e, nelle forme alate, l'assenza delle ali. Il passaggio dallo stadio giovanile a quello adulto si svolge in uno o più stadi di ninfa , durante i quali si verifica la comparsa degli abbozzi alari esterni. La ninfa è mobile ad eccezione della neometabolia , un tipo di metamorfosi intermedio fra l'eterometabolia propriamente detta e l'olometabolia.
Ai due descritti in precedenza si aggiunge un terzo tipo, detto ametabolia , in cui, in realtà, vi è assenza di metamorfosi. L'ametabolia si verifica negli insetti primitivamente atteri ( Tisanuri ): in questo caso gli stadi giovanili non presentano alcuna differenza morfologica rispetto agli adulti, se non nelle dimensioni, e lo sviluppo si attua con mute senza alcuna metamorfosi. L'ametabolia non va confusa con la pseudoametabolia , una forma di eterometabolia tipica di specie secondariamente attere.
Lo schema descritto si complica se si considera la neometabolia una metamorfosi distinta e intermedia fra quella incompleta e quella completa e se si prendono in esame le varianti che ricorrono nell'ambito dell'eterometabolia e dell'olometabolia. Infine, si distingue un quinto tipo di metamorfosi, detto catametabolia , che vede una regressione anatomica e morfologica dell'insetto nel corso dello sviluppo ontogenetico. In altri termini, l'adulto ha una forma e una struttura più semplice di quella dello stadio giovanile. La catametabolia in realtà è una variante dovuta all'adattamento della specie ad una particolare etologia ed è riscontrabile sia nell'olometabolia sia nell'eterometabolia.
Alimentazione

I regimi dietetici degli Insetti sono tra i più svariati e possono cambiare drasticamente anche nel corso dello sviluppo di un individuo ( olometaboli e paurometaboli ). La maggior parte degli Insetti ha rapporti trofici con i vegetali o con gli animali , oppure manifesta un regime onnivoro . Sono frequenti anche i detritivori o, più in generale, saprofagi ; non mancano, infine, casi di insetti micetofagi . Una classificazione rigorosa, tuttavia, non è possibile in quanto i limiti che definiscono i regimi dietetici sono spesso labili.
Fra gli insetti che si nutrono a spese dei vegetali vivi si distinguono due categorie: i fitofagi ei fitomizi . Sono spesso indicati, impropriamente, come parassiti , ma il rapporto trofico che li lega alle piante si identifica in una forma di predazione riconducibile all' erbivoria . I rapporti possono essere più o meno specifici e si distinguono specie monofaghe, oligofaghe e polifaghe. I fitofagi hanno apparato boccale masticatore ed erodono i tessuti vegetali attaccando gli organi dall'esterno o dall'interno ( minatori ). Una terminologia informale identifica i fitofagi in base all'organo o al tessuto attaccato: i fillofagi (a spese delle foglie), gli antofagi (fiori), i carpofagi (frutti), gli xilofagi (legno), ecc.
I fitomizi sono invece insetti ad apparato boccale pungente-succhiante e si nutrono a spese della linfa elaborata o dei succhi cellulari di cellule epidermiche. Un gruppo particolare è rappresentato dagli insetti galligeni , che si sviluppano all'interno di un'alterazione istologica dell'organo colpito ( galla ). In questo caso la fitofagia si avvicina meglio al concetto di parassitismo.
Per quanto concerne la zoofagia, si può fare una distinzione, per il differente ambito d'interesse, fra specie che attaccano i Vertebrati e le altre che si nutrono a spese di Invertebrati. Anche in questo caso si possono individuare forme di monofagia, oligofagia e polifagia. La prima categoria annovera un limitato numero di specie, spesso d'interesse medico o veterinario . I rapporti trofici si collocano a metà strada fra il parassitismo e la predazione , con vari livelli di affinità verso il primo o il secondo. Si nutrono a spese dell' epidermide o dei suoi annessi oppure di sangue ( ematofagi ). Sono rari i casi di predazione vera e propria (es. i Ditiscidi e alcuni Rincoti Eterotteri predano anche piccoli Vertebrati acquatici). Le forme assimilabili ai parassiti sono in genere epizoiche, tuttavia non mancano casi di endoparassiti (es. alcune famiglie di Ditteri). Fra i carnivori si annoverano anche le specie prettamente saprofaghe che si nutrono a spese delle spoglie di Vertebrati morti ( sarcofaghe ).
La maggior parte delle specie a regime zoofago ha però rapporti trofici con altri artropodi e, in particolare, con altri insetti. In questi casi i rapporti si identificano con la predazione vera e propria o con il parassitoidismo , mentre i casi di parassitismo vero e proprio sono alquanto rari. Fra gli artropofagi si annoverano anche gli oofagi che predano o parassitizzano le uova. In generale si tratta di specie di grande interesse perché possono essere sfruttate nella lotta biologica e integrata . Non è raro il cannibalismo , in genere fra predatori ma, talvolta, anche fra i fitofagi; questo fenomeno si verifica frequentemente in condizioni di scarsa disponibilità alimentare o in cattività [27] [28] [29] [30] . Meno frequente è invece il parassitoidismo fra individui della stessa specie, fenomeno noto con il termine di autoparassitismo e che si riscontra soprattutto negli Imenotteri Afelinidi , nei quali si riscontrano comportamenti di particolare interesse naturalistico [31] , e nelle già citate specie con riproduzione pedogenetica.
Un regime dietetico particolare è quello dei glicifagi . Questi insetti si nutrono a spese di sostanze zuccherine e in generale usano come substrato alimentare il nettare dei fiori e la melata emessa dai Rincoti oi succhi che fuoriescono da ferite. Le fonti proteiche utilizzate sono rappresentate, secondo i casi, dal polline , da escrementi di animali o da altri materiali. L'apparato boccale è di tipo succhiante o lambente-succhiante oppure di tipo masticatore più o meno adattato. Molti glicifagi sono importanti come insetti pronubi , in quanto provvedono al trasporto del polline e, quindi, favoriscono l'impollinazione incrociata.
Alcuni regimi dietetici sono particolarmente complessi e non hanno una collocazione ben definita: ad esempio, alcuni Imenotteri parassitoidi di insetti galligeni possono diventare fitofagi quando, una volta esaurita la vittima, completano il loro ciclo di sviluppo nutrendosi a spese dei tessuti della galla. Anche fra gli Insetti si annoverano, comunque, specie onnivore, in grado di nutrirsi indifferentemente con alimenti di origine sia vegetale sia animale.
Etologia

Per i motivi esposti in precedenza, gli aspetti comportamentali di questa classe sono eterogenei e svariati, talvolta con livelli di complessità non irrilevanti. I comportamenti possono rappresentare delle peculiarità proprie di una o poche specie oppure estese a gruppi sistematici più ampi.
Uno degli aspetti più interessanti dell'etologia è la vita degli insetti sociali , specie che hanno sviluppato un livello complesso di gregarismo organizzato per caste e basato sulla costruzione di nidi complessi, la trasmissione di stimoli sensoriali, la cura della prole, la ricerca, l'approvvigionamento e la conservazione degli alimenti, la regolazione della riproduzione, ecc. Questi fenomeni etologici, presi nel complesso, determinano il successo ecologico della comunità sacrificando il singolo individuo. Agli esempi più noti di aggregazione sociale, che interessano per lo più gli Isotteri e gli Imenotteri , si aggiunge una vasta gamma di fenomeni di aggregazione non sociale, rappresentati nella maggior parte degli ordini, con differenti livelli di complessità in merito alle relazioni interne. All'aggregazione intraspecifica si associano spesso relazioni interspecifiche che rientrano nel novero dei fenomeni di commensalismo e simbiosi .
A prescindere dalle aggregazioni sociali, l'etologia degli Insetti mostra interessanti e variegati comportamenti per quanto concerne l'alimentazione, la costruzione di nidi, la riproduzione, la migrazione, la comunicazione fra individui della stessa specie, la relazione con organismi di specie differenti, la locomozione, il mimetismo , la difesa, l'emissione di luci o suoni, ecc. Per una trattazione di questi aspetti si rimanda alle voci relative ai singoli gruppi sistematici.
Dinamiche di popolazione

Anche per quanto concerne la dinamica delle popolazioni, si verificano modelli differenti, sui quali interferisce spesso l'Uomo alterando o semplificando gli ecosistemi . In generale, gli Insetti hanno - come la maggior parte degli Invertebrati - una strategia di vita basata sul potenziale riproduttivo ( strategia r ), perciò manifestano una straordinaria rapidità di colonizzazione degli ecosistemi che perdono le loro condizioni di equilibrio. Questo problema si presenta in modo particolare negli agrosistemi , in cui la riduzione della biodiversità crea squilibri da cui traggono beneficio proprio le specie dotate di grande potenziale riproduttivo [32] .
In condizioni naturali molte specie hanno dinamiche basate sulle fluttuazioni : il livello della popolazione si mantiene, per periodi più o meno lunghi, in una fase di latenza, in equilibrio con la disponibilità alimentare e la cenosi degli antagonisti. Periodicamente, a intervalli più o meno regolari, si manifestano le gradazioni , repentini cambiamenti della densità di popolazione rispetto alla fase di latenza. Nel corso di una gradazione si hanno tre fasi [32] [33] :
- progradazione : la popolazione mostra ritmi di crescita esponenziale, in virtù del forte potenziale biotico, della disponibilità di risorse nutritive e della scarsa incidenza dei fattori naturali di controllo;
- culmine : la popolazione aumenta con incrementi decrescenti, fino a raggiungere un massimo; in questa fase diventano incisivi i fattori di controllo naturali della popolazione: competizione intraspecifica, riduzione della fertilità, insorgenza di malattie e incremento della popolazione degli antagonisti;
- retrogradazione : a causa della forte incidenza dei fattori naturali di controllo, la mortalità aumenta notevolmente e la popolazione crolla più o meno rapidamente fino a riportarsi sui valori di equilibrio della fase di latenza.
Il fenomeno delle gradazioni si riscontra in genere negli ecosistemi forestali e interessa in particolare i Lepidotteri ei Coleotteri defogliatori. L'entità di una gradazione è tale che può portare alla completa spogliazione di un'intera foresta. Lo studio della dinamica di popolazione, il monitoraggio e l'elaborazione di modelli previsionali hanno lo scopo di prevenire le gradazioni che, quando si protraggono per due o tre anni consecutivi, possono causare danni di grande entità sul patrimonio forestale.
Importanza degli Insetti
L'ampia diffusione della classe e la frequente interazione con l' Uomo coinvolgono gli Insetti in vari ambiti d'interesse, oltre a quello strettamente zoologico e naturalistico . L'elevato numero di specie presenti in questa classe ha portato all'evoluzione di una branca specifica della Zoologia, detta Entomologia , che a sua volta si suddivide in differenti rami di specializzazione. La complessità della materia è tale che, in genere, gli ambiti di studio dei singoli esperti si concentrano su un solo ordine o, talvolta, su gruppi sistematici più ristretti. Ciò ha portato alla costituzione di associazioni accademiche di livello internazionale il cui interesse verte su singoli raggruppamenti tassonomici ("Emitteristi", "Coleotteristi", ecc.)
Dannosità

La dannosità degli Insetti è forse l'aspetto più evidente che si associa a questa classe; in realtà il numero di specie significativamente dannose è irrisorio a fronte del numero di specie utili o indifferenti. Queste poche specie possono però causare danni di tale entità da costituire, alcune, dei veri e propri flagelli. Lo studio degli Insetti come fattori di danno si colloca in quattro ambiti d'interesse:
- Agroforestale. Sono potenzialmente dannosi gli Insetti che attaccano le specie vegetali da cui l'uomo trae utilità diretta o indiretta. Il grado d'importanza è rapportato all'entità dei danni, perciò il numero delle specie realmente dannose si riduce sensibilmente. D'altra parte ogni anno ci sono insetti che da uno status di specie indifferente passano ad uno di specie dannosa a causa di emergenze scaturite da particolari contesti (importazione da altre regioni, degradazione degli equilibri ecologici, ecc.). Le specie fitofaghe di maggiore importanza rientrano generalmente negli ordini dei Rincoti , degli Ortotteri , dei Lepidotteri , dei Ditteri , degli Imenotteri e dei Coleotteri .
- Agroalimentare. Sono generalmente dannosi tutti gli insetti che attaccano le derrate, in quanto possono causare la distruzione totale o l'inservibilità di intere partite stoccate nei magazzini o accelerare la decomposizione dei prodotti deperibili. Le specie dannose alle derrate si annoverano, in genere, fra i Coleotteri, i Lepidotteri, i Ditteri ed i Blattoidei . Oltre ai danni diretti va considerato anche il rischio igienico-sanitario dovuto alla possibile trasmissione di agenti patogeni per l'uomo, frequente nel caso di Blattoidei e Ditteri.
- Tecnologico. In questo ambito sono particolarmente dannosi gli insetti xilofagi , che attaccano il legno da opera e tutti i suoi manufatti. Sotto questo aspetto gli ordini di maggiore interesse sono gli Isotteri (termiti), particolarmente dannosi nelle aree tropicali, ei Coleotteri (tarli). Altri insetti d'interesse tecnologico sono le tarme , microlepidotteri saprofagi che attaccano i manufatti tessili.
- Igienico-sanitario. Sono d'interesse medico o veterinario gli insetti che attaccano i vertebrati superiori, in particolare i parassiti epizoici, gli endoparassiti ei predatori ematofagi che attaccano i Mammiferi e gli Uccelli . Questi insetti sono responsabili di affezioni dirette oppure microbiche in quanto vettori di Virus e microrganismi patogeni. D'interesse igienico-sanitario sono anche quegli insetti prettamente saprofagi possibili vettori di patogeni. Gli insetti d'interesse igienico-sanitario fanno capo soprattutto agli ordini dei Mallofagi , degli Anopluri , dei Ditteri e dei Sifonatteri .
Utilità

Ai danni causati da molte specie di Insetti si contrappone l'utilità di molte altre specie, soprattutto come organismi inseriti negli ecosistemi e coinvolti nelle reti alimentari. Sono moltissimi, tuttavia, i casi in cui gli insetti sono direttamente utili all'Uomo.
L'aspetto più importante, per gli studi dedicati e la sempre più diffusa applicazione, riguarda il ruolo degli insetti come organismi ausiliari. Gli artropofagi sono utili, se non essenziali, come fattori di controllo naturali di artropodi dannosi. Questo ruolo è oggi messo in primo piano nelle varie forme di agricoltura sostenibile ( agricoltura biologica , coltivazioni condotte con i criteri della lotta biologica ed integrata , ecc.) e in ambito forestale. Sono utili anche alcuni fitofagi, quando la loro azione biologica si esercita su piante infestanti, come ad esempio si è verificato nel controllo delle Opuntia in Australia , impiegando il Cactoblastis cactorum e il Dactylopius opuntiae . Non meno importante è il ruolo degli insetti pronubi , compresi soprattutto negli ordini dei Ditteri e degli Imenotteri , essenziali per la fruttificazione di molte specie agrarie, che si avvalgono dell'impollinazione entomofila.
L'importanza degli insetti ausiliari è tale che ormai sono una realtà economica le cosiddette biofabbriche , allevamenti industriali di Insetti da impiegare nella lotta biologica o nella lotta biotecnica (es. maschi sterili). Questo settore d'impiego ha finora occupato un ruolo marginale, soprattutto per l'elevato costo, poco competitivo nei confronti del settore della Industria chimica , tuttavia è in espansione per la domanda crescente di salubrità ambientale e alimentare.

Gli impieghi economici erano di grande importanza soprattutto in passato. Prima dell'avvento della chimica industriale, molte specie erano sfruttate per ricavarne sostanze utilizzate per vari scopi, come coloranti, lacca, seta, cera. Emblematica è l'importanza che ha avuto il bombice del gelso , il cui prodotto, la seta , è stato per secoli il protagonista degli scambi commerciali tra l' Asia e l' Europa . L'impiego economico per eccellenza, attualmente, riguarda le Api , utilizzate per la produzione del miele e della cera e, secondariamente, della pappa reale e della propoli .
Di importanza marginale e locale, ma solo in apparenza, è l'uso degli insetti nell'alimentazione umana, presso varie popolazioni nell' America centrale , in Africa e in Asia [34] . È stato accertato che il numero di specie sfruttate come cibo dall'Uomo ammonta ad oltre 1400 [35] . Alcune specie sono considerate vere e proprie leccornie ricercate sul mercato come, ad esempio, l' axayácatl in Messico , dalle cui uova si ottiene l' ahuautle o caviale messicano [36] [37] , ed i Belostomatidi in Asia [38] [39] . Tuttavia l'aspetto più importante, a cui la stessa FAO dedica una particolare attenzione, è la potenziale risorsa alimentare che gli insetti possono rappresentare per arginare il problema della sottoalimentazione, per l'alto valore nutritivo, superiore a quello della carne e del pesce : i bruchi essiccati contengono, infatti, oltre il 50% in proteine , il 17% in glucidi e il 15% in lipidi [40] .
Va infine ricordato l'interesse riscosso dagli Insetti nel settore dell' hobbistica in quanto sono soggetti ricercati sia per l'allestimento delle collezioni entomologiche sia per la fotografia naturalistica.
Inquadramento tassonomico
Nelle vecchie classificazioni la Classe Insecta , sinonimo di Hexapoda , comprendeva anche alcuni ordini di Artropodi primitivamente atteri. Revisioni più recenti attribuiscono la denominazione Hexapoda ad un raggruppamento sistematico superiore, elevato al rango di Superclasse, al cui interno si fa distinzione fra due classi: la prima, Paraentoma o Entognatha si identifica in un gruppo monofiletico [41] [42] [43] comprendente quelli che in passato erano considerati gli ordini più primitivi degli insetti, la seconda, Euentomata o Ectognatha , comprende gli Insetti propriamente detti, corrispondente ad un altro raggruppamento monofiletico.
L'inquadramento sistematico della classe si può riassumere nel seguente schema:
- Superclasse Hexapoda . Compresa nel subphylum Tracheata , a sua volta facente parte del phylum Arthropoda . Si suddivide in due raggruppamenti sistematici:
- Paraentoma o Entognatha . Raggruppamento parallelo agli insetti, racchiude gli esapodi dotati di mascelle e mandibole interne al capo e si suddivide in tre sottoclassi: Oligoentomata (comprendente i Collemboli ), Mirientomata (comprendente i Proturi ), Polientomata (comprendente i Dipluri ).
- Euentomata o Ectognatha o Insecta propriamente detti. Raggruppamento monofiletico elevato al rango sistematico di classe che include gli esapodi con un apparato boccale esterno alla testa. Si suddivide tassonomicamente in due sottoclassi in base alla presenza delle ali come carattere apomorfo :
- Apterygota . Sottoclasse comprendente gli Insetti primitivamente atteri e ametaboli.
- Pterygota . Sottoclasse comprendente gli Insetti alati o secondariamente atteri e che subiscono una metamorfosi nel corso dello sviluppo postembrionale.
La classificazione all'interno della classe si basa fondamentalmente sulla metamorfosi e sulla morfologia alare. Va in ogni caso ribadito che la sistematica degli Insetti è in continua evoluzione a causa dell'elevato numero di specie e della continua acquisizione di nuove conoscenze, soprattutto nel campo della filogenesi . La maggior parte degli Autori considera di particolare importanza la ripartizione in ordini che, storicamente, non ha subito sostanziali modifiche. Gli ordini compresi nello schema tradizionale classico sono 28:
Invece gli ordini dello schema più moderno comprende 27 ordini:
Questo schema si differenzia dallo schema di suddivisione indicato in precedenza, nei seguenti punti:
- Sulla base delle relazioni filogenetiche, Blattodea, Mantodea e Isoptera sono attualmente inclusi in un unico ordine che prende il nome di Dictyoptera.
- Analogamente, Mallophaga e Anoplura sono inglobati in un unico ordine che prende il nome di Phthiraptera.
- I Neuroptera sensu lato sono attualmente considerati un clade filogenetico (da alcuni un superordine) comprendente tre ordini: Raphidioptera, Megaloptera e Neuroptera sensu stricto (corrispondenti al sottordine Planipennia dei Neuroptera sensu lato ).
Queste revisioni sono attualmente adottate nella generalità delle pubblicazioni scientifiche, per quanto sui manuali tecnici, più o meno recenti, e in alcuni settori dell'Entomologia applicata, come l'Entomologia agraria, si continui ad usare sovente la vecchia suddivisione.
Note
- ^ Tremblay , p. 11 .
- ^ Tremblay , pp. 12-13 .
- ^ Worldwide decline of the entomofauna: A review of its drivers , DOI : 10.1016/j.biocon.2019.01.020 .
- ^ ( EN ) Ed Yong, Insects Are In Serious Trouble , su The Atlantic , 19 ottobre 2017. URL consultato il 6 aprile 2019 ( archiviato il 18 febbraio 2021) .
- ^ Insetto deriva dal latino insecta , Entomo dal greco éntoma . In entrambi i casi l'etimologia fa riferimento alla segmentazione del corpo.
- ^ Staněk , p. 270 .
- ^ Staněk , p. 377 .
- ^ Servadei et al. , pp. 18-19 .
- ^ Tremblay , pp. 31-32 .
- ^ Gert Stange, Jonathon Howard, An ocellar dorsal light response in a dragonfly ( PDF ), in J. Exp. Biol. , vol. 83, 1979, pp. 351-355. URL consultato il 28 dicembre 2007 .
- ^ Per convenzione, gli uriti sono indicati con un numero romano progressivo, partendo da quello anteriore
- ^ Tremblay , p. 24 .
- ^ Servadei et al. , p. 109 .
- ^ Tremblay , p. 51 .
- ^ La melata, insieme al nettare , costituisce la base alimentare degli adulti di diversi gruppi sistematici, per la maggior parte compresi negli ordini dei Ditteri e degli Imenotteri . Nelle foreste di Conifere è la principale materia prima utilizzata dalle api per la produzione del miele .
- ^ a b Fanno eccezione gli Insetti apneustici, privi di spiracoli tracheali, nei quali il trasporto dei gas è affidato all'emolinfa.
- ^ Servadei et al. , p. 71 .
- ^ Tremblay , p. 39 .
- ^ a b Tremblay , pp. 41-42 .
- ^ Servadei et al , p. 80 .
- ^ Servadei et al. , p. 89 .
- ^ ( EN ) Michael S. Engel, David A. Grimaldi, New light shed on the oldest insect , in Nature , vol. 427, 2004, pp. 627-630. URL consultato il 18 gennaio 2015 ( archiviato il 20 settembre 2011) .
- ^ Tremblay , p. 50 .
- ^ a b Servadei et al. , p. 105 .
- ^ Servadei et al. , p. 100 .
- ^ Tremblay , p. 55 .
- ^ Luca Conte, Controllo biologico: come impiegare gli insetti utili? , su Controllo biologico (Traduzione e adattamento dalla versione originale "Biological Control: Application of Natural Enemies" di D. Orr e J. Baker, Department of Entomology of North Carolina State University) , Gruppo di ricerca Entomologia, Università di Padova. URL consultato il 5 luglio 2008 (archiviato dall' url originale il 6 ottobre 2008) .
- ^ Tremblay. , p. 74 .
- ^ Servadei et al. , pp. 256, 313, 562 .
- ^ Giorgio Nicoli e Paolo Radenghieri (a cura di). Gli ausiliari nell'agricoltura sostenibile . Bologna, Calderini Edagricole, 2000, pp. 40, 92. ISBN 88-206-4504-1 .
- ^ Gennaro Viggiani. Lotta biologica e integrata . Napoli, Liguori Editore, 1977, pp- 422-423. ISBN 88-207-0706-3
- ^ a b Tremblay , pp. 73-81 .
- ^ Servadei et al. , pp. 165-168 .
- ^ ( EN ) Gene R. De Foliart, The Human Use of Insects as a Food Resource: A Bibliographic Account in Progress . URL consultato il 6 luglio 2008 ( archiviato l'11 marzo 2003) .
- ^ Fonte FAO, Forest insects as food: humans bite back. A Workshop focused on Asia-Pacific resources and their potential for development , in Non-Wood News , n. 16, 2008, p. 71.
- ^ Julieta Ramos-Elorduy, Threatened edible insects in Hidalgo, Mexico and some measures to preserve them , in Journal of Ethnobiology and Ethnomedicine , vol. 2, 2006, p. 51, DOI : 10.1186/1746-4269-2-51 . URL consultato il 26 aprile 2008 .
- ^ ( EN ) Just when thought a worm in Tequila was weird... Mexican Insects as Food! [ collegamento interrotto ] , su In Mexico Guide . URL consultato il 26 aprile 2008 .
- ^ Staněk , p. 111 .
- ^ ( EN ) Gene R. De Foliart, Chapter 25. Other Countries in Southeastern Asia , in The Human Use of Insects as a Food Resource: A Bibliographic Account in Progress (archiviato dall' url originale l'11 marzo 2003) .
- ^ ( EN ) Edible insects, important source of protein in central Africa , su FAO Newsroom , Food and Agriculture Organization of the United Nations, 2004. URL consultato il 7 luglio 2008 ( archiviato il 25 giugno 2008) .
- ^ C. Bitsch e J. Bitsch., Internal anatomy and phylogenetic relationships among apterygote insect clades (Hexapoda) , in Annales de la Société Entomologique de France , vol. 3, 1998.
- ^ A. Carapelli, F. Frati, F. Nardi, C. Simon, Molecular phylogeny of apterygotan insects based on nuclear and mitochondrial genes , in Proceedings of the 5th International Symposium on Apterygota , 1998.
- ^ D'Haese, CA, Were the first springtails semi-aquatic? A phylogenetic approach by means of 28S rDNA and optimization alignment , in Proceedings of the Royal Society of London, Series B, Biological Sciences , vol. 269, 2002.
Bibliografia
- Ermenegildo Tremblay. Entomologia applicata . Volume I. 3ª ed. Napoli, Liguori Editore, 1985. ISBN 88-207-0681-4
- Antonio Servadei; Sergio Zangheri; Luigi Masutti. Entomologia generale ed applicata . Padova, CEDAM, 1972.
- Guido Grandi. Istituzioni di entomologia generale . Bologna, Calderini, 1966. ISBN 88-7019-084-6 .
- Aldo Pollini. Manuale di entomologia applicata . Bologna, Edagricole, 2002. ISBN 88-506-3954-6 .
- Vincent B. Wigglesworth. Insect Physiology . London, Meuthuen & Co., 1934. In inglese.
- Václav Jan Staněk. Enciclopedia illustrata degli insetti . Elisabetta Ghisotti Steinman (Trad. it.). Praga, Artia (Ed. it. Librerie Accademia) [1970], 1978. ISBN 0-600-03085-7 .
- Ângelo Moreira da Costa Lima. Insetos do Brasil . Escola Nacional de Agronomia, 1940. In portoghese.
- Chris Durden. Learn about... Texas Insects. A Learning and Activity Book . Elena T. Ivy (Art Director). Austin, Texas Parks and Wildlife Press, 1999. ISBN 1-885696-27-2 . In inglese.
- Michael Chinery. Field Guide to Insects of Britain and Northern Europe . Volume 3. Auflage, Harper Collins, 1993. ISBN 0-00219-918-1 . In inglese.
- Michael Chinery. Pareys Buch der Insekten . Franckh-Kosmos, 2004. ISBN 3-440-09969-5 . In tedesco.
Voci correlate
Altri progetti
-
 Wikiquote contiene citazioni sugli insetto
Wikiquote contiene citazioni sugli insetto -
 Wikizionario contiene il lemma di dizionario « insetto »
Wikizionario contiene il lemma di dizionario « insetto » -
 Wikimedia Commons contiene immagini o altri file sugli insetto
Wikimedia Commons contiene immagini o altri file sugli insetto -
 Wikispecies contiene informazioni sugli insetto
Wikispecies contiene informazioni sugli insetto
Collegamenti esterni
- ( EN ) Insecta / Insecta (altra versione) , su Enciclopedia Britannica , Encyclopædia Britannica, Inc.
- ( EN ) Insecta , in Encyclopædia Iranica , Ehsan Yarshater Center, Columbia University.
- ( EN , FR ) Insecta , su Enciclopedia canadese .
- ( EN ) Opere riguardanti Insecta , su Open Library , Internet Archive .
- ( EN ) Insecta , su Fossilworks.org .
- ( EN ) Insects , su The Visual Novel Database .
- Siti tematici contenenti trattazioni relative alla classe degli Insetti
- Leonardo Melchionda, Insetti , su webalice.it . URL consultato il 27 dicembre 2008 (archiviato dall' url originale il 6 gennaio 2010) .
- Leonardo Melchionda, Insetti , su Google Sites.
- Canale difesa contro i parassiti delle colture agrarie, ornamentali, forestali , su Agrolinker. Il portale per lo studente e lo studioso delle scienze agrarie e forestali . URL consultato il 2 luglio 2008 .
- Forum Entomologi Italiani , su entomologiitaliani.net . URL consultato il 16 novembre 2009 .
- ( EN ) Systematic Entomology Laboratory (SEL) , su ars.usda.gov , Agricultural Service Research, US Department of Agriculture. URL consultato il 2 luglio 2008 (archiviato dall' url originale il 21 maggio 2008) .
- ( EN ) Australian Faunal Directory: Main Checklist [ collegamento interrotto ] , su environment.gov.au , Department of Australian Government of Environment, Water, Heritage and the Arts. URL consultato il 2 luglio 2008 .
- ( EN ) John Meyer, General Entomology , su cals.ncsu.edu , North Carolina State University. URL consultato il 4 luglio 2008 .
- ( EN ) Gordon John Larkman Ramel, The Insects Home Page , su The Earth Life Web . URL consultato il 4 luglio 2008 .
- ( EN )BugGuide , su bugguide.net . URL consultato il 4 luglio 2008 .
- ( EN ) Entomology Image Gallery , su ent.iastate.edu , Department of Entomology, Iowa State University. URL consultato il 4 luglio 2008 .
- ( EN ) North American Insects and Spiders , su cirrusimage.com , Red Planet Inc.. URL consultato il 4 luglio 2008 .
- ( EN ) Research Projects: Department of Entomology , su The Natural History Museum, London . URL consultato il 4 luglio 2008 .
- ( EN ) Insecta. Insects , su The Tree of Life Web Project , 2002. URL consultato il 4 luglio 2008 .
- ( DE ) Arthropoda - Insekten und Spinnen Mitteleuropas , su arthropods.de . URL consultato il 28 luglio 2008 .
- ( DE ) Insekten - Box: Steckbriefe , su insektenbox.de . URL consultato il 28 luglio 2008 .
- ( EN ) Bugbios - Insects on the Web , su bugbios.com . URL consultato il 28 luglio 2008 (archiviato dall' url originale il 9 maggio 2008) .
- ( DE ) Bilder fliegender Insekten , su insektenflug.de . URL consultato il 28 luglio 2008 .
- Siti tematici contenenti trattazioni relative a specifici ordini di insetti
- ( EN , RU ) Beetles (Coleoptera) and Coleopterists , su zin.ru , Zoological Institute, Russian Academy of Sciences, Saint Petersburg. URL consultato il 2 luglio 2008 .
- ( EN ) Dragonflies and Damselfies (Odonata Information Network) , su iodonata.net , International Odonata Research Institute. URL consultato il 4 luglio 2008 .
- ( EN ) The Orthopterist's Society , su 140.247.119.138 . URL consultato il 4 luglio 2008 (archiviato dall' url originale il 5 giugno 2008) .
- ( EN ) P. Mazzei, D. Reggianti; I. Pimpinelli,Moths and Butterflies of Europe and North Africa , su leps.it . URL consultato il 4 luglio 2008 .
- Checklist e banche dati tassonomia
- ( EN ) Taxon details: Insecta , in Fauna Europaea version 2.6.2 , Fauna Europaea Web Service, 2013. URL consultato il 04-07-2008 .
- ( EN ) ITIS Standard Report Page: Insecta , in Integrated Taxonomic Information System . URL consultato il 04-07-2008 .
- Fabio Stoch, Fauna Italia , su faunaitalia.it . URL consultato il 2 luglio 2008 .
Per siti tematici relativi a gruppi sistematici inferiori si rimanda alle specifiche sezioni.
| Controllo di autorità | Thesaurus BNCF 634 · LCCN ( EN ) sh85066670 · GND ( DE ) 4027110-9 · BNF ( FR ) cb11932119n (data) · BNE ( ES ) XX524421 (data) · NDL ( EN , JA ) 00566605 |
|---|