Araneae
| Păianjen | |
|---|---|
 | |
| Gama geologică | |
| Clasificare științifică | |
| Domeniu | Eukaryota |
| Regatul | Animalia |
| Sub-regat | Eumetazoa |
| Superphylum | Protostomie Ecdysozoa |
| Phylum | Arthropoda |
| Subfilum | Chelicerata |
| Clasă | Arachnida |
| Ordin | Araneae Clerck , 1757 |
| Subordine și Infraordine | |

Araneidele ( Araneae Clerck , 1757 ) sunt un ordin de arahnide , împărțit, în noiembrie 2019, în 120 de familii care includ 48 349 de specii. [1]
Sunt artropode terestre echipate cu chelicere și au corpul împărțit în două segmente, cefalotorax și opistosom , și opt picioare. Cele două segmente sunt unite de un pedicel cilindric mic. [2] La fel ca în toate artropodele, celomul (cavitatea mezodermului pentru transportul fluidelor), deși foarte mic, permite trecerea hemolimfei care oxigenează și hrănește țesuturile și elimină deșeurile. Intestinul este atât de îngust încât păianjenii nu pot mânca bucăți de material solid, oricât de mici ar fi, și sunt forțați să-și lichideze alimentele cu diferite enzime digestive și să le macine fin cu aparatul de mestecat.
Cu excepția celui mai primordial subordine, cel al Mesotelelor , păianjenii au cel mai centralizat sistem nervos dintre toate artropodele și, ca acestea, au ganglionii cefalici fuzionați într-o singură masă din cefalotorax. Spre deosebire de majoritatea artropodelor, păianjenii nu au mușchi extensori în corp; mișcările picioarelor și ale corpului se obțin prin variații de presiune ale sistemului lor hidraulic. [3] În partea terminală a abdomenului există filiere care extrudă mătasea, utilizate pentru a înfășura prada și a construi pânzele de păianjen.
Caracteristici generale

Singura specie vegetariană de păianjeni a fost descrisă în 2007 [4] ; toți ceilalți sunt prădători, atât ai altor păianjeni, cât și ai insectelor; speciile mai mari vin să prădeze chiar și păsări mici și șopârle fără dificultate. Din diverse studii și observații s-a constatat că exemplarele tinere își completează dieta cu nectar , în timp ce exemplarele adulte preferă să completeze cu polen . La mai multe specii de păianjeni, chelicerele au devenit capabile să secrete o otravă pentru a imobiliza prada, uneori atât de toxică încât să fie periculoasă chiar și pentru oameni. Recent, doze mici din aceste otrăvuri sunt testate în scopuri terapeutice sau ca insecticide nepoluante. Majoritatea păianjenilor își captează prada prinzându-i în pânze de păianjen lipicioase, ascunzându-se în apropiere și apoi injectându-i otravă. Speciile care folosesc această tactică de vânătoare sunt foarte sensibile la cea mai mică vibrație care scutură firele rețelei. Pe de altă parte, speciile de urmărire sunt prevăzute cu o vedere excelentă, de până la zece ori mai clară decât cea a unei libelule . [5]
Unii păianjeni vânători au abilități și inteligență [ fără sursă ] în utilizarea diferitelor tactici pentru a copleși prada, arătând, de asemenea, că poate învăța altele noi, dacă este necesar. De exemplu, unele specii și-au schimbat aspectul luând aspectul exterior al furnicilor până la punctul de a fi ușor confundat cu una dintre ele. [6] Pânzele de păianjen sunt foarte variabile ca formă, cantitatea de pânză folosită și dimensiune. Se pare că primele forme care au fost adoptate au fost cele orbiculare; speciile care încă le folosesc sunt puține: majoritatea păianjenilor existenți preferă să-și extindă pânza și să o facă cât mai încâlcită pentru a maximiza volumul de aer în care se poate întâlni o insectă.
La păianjenii masculi, pedipalpii (apendicele situate sub sau în partea laterală a deschiderii bucale) au fost modificați ca o seringă pentru a injecta sperma în tractul genital feminin. Unii dintre ei trebuie să efectueze ritualuri de curte complicate înainte de a se putea apropia de femeie pentru a o fertiliza, pentru a evita să fie devorați. [7] Masculii majorității speciilor scapă de această soartă, iar femela le permite să se poziționeze în părțile marginale ale pânzelor chiar și pentru o perioadă de timp după împerechere. Pe de altă parte, există masculi din unele specii care, după ce au fecundat femela, i se oferă ei, probabil pentru a furniza mai mulți nutrienți descendenților care se vor naște. La unele specii femelele construiesc un fel de ovatură mătăsoasă în care cel puțin o sută de ouă găsesc spațiu; tinerii, odată născuți, sunt în orice caz incapabili să obțină singuri hrană, până când nu vor finaliza prima lor năpârlire. [8] În aceste cazuri, femelele / mamele sunt cele care hrănesc descendenții și împărtășesc prada capturată cu aceasta.

Doar câteva specii de păianjeni au un comportament social, adică construiesc pânze în cooperare cu alți păianjeni, până la câteva mii; [9] comportamentul social este variat: de la o simplă toleranță de vecinătate, ca la văduva neagră agresivă, la o vânătoare reală coordonată între mai multe exemplare și subdiviziune ulterioară a prăzii capturate. În ceea ce privește longevitatea, majoritatea păianjenilor nu depășesc doi ani, se știe că doar câteva exemplare de tarantule și diferite migalomorfe captive au supraviețuit până la douăzeci și cinci de ani. [7]
Din studiul rămășițelor fosile găsite, primele animale capabile să producă mătase apar în Devonian , în urmă cu aproximativ 386 de milioane de ani [10] , însă le lipsește orice urmă de lanțuri de aprovizionare . Primii păianjeni fosili adevărați au fost găsiți în roci carbonifere , databile prin urmare între 299 și 318 milioane de ani în urmă, și sunt foarte asemănători cu cei actuali aparținând ordinului Mesotelelor . Principalele grupuri de păianjeni moderni, Araneomorphae și Mygalomorphae , au apărut în Triasicul inferior , acum aproximativ 200 de milioane de ani. [10] Doar în fosilele datate de acum 130 de milioane de ani există păianjeni păstrați în chihlimbar , având o bogăție mai mare de detalii vizibile și analizabile, în special detalii anatomice. Chihlimbarul a fost găsit, de asemenea, cu o îmbrățișare mortală între păianjen și prada acestuia, în timp ce îl coconează și altele în care au fost păstrate și ouăle din sacii ovigeroși .
Descriere
Structura corpului
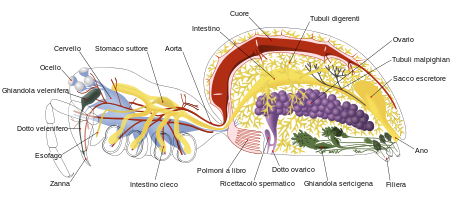 |  |
Păianjenii sunt chelicerați și ca artropode au un corp împărțit în segmente cu margini articulate, acoperite de o cuticulă compusă din chitină și proteine. [11] Capul este format din diferite segmente care se îmbină între ele în timpul dezvoltării embrionului . [12] Pe măsură ce chelicerează, corpul lor este format din două tagmate , segmentare tipică a artropodelor, un set de segmente care stau la baza unei funcții similare. Prima dintre acestea, care poartă numele de cefalotorax sau prosom, este fuziunea completă a două tagmate care într-o insectă ar fi separate: capul și toracele; tagma posterioară ia numele de abdomen sau opistosom . [11] Modelul fuziunii segmentelor care alcătuiesc capul cheliceratelor este unic printre artropode: ceea ce ar fi primul segment al capului dispare de obicei la începutul dezvoltării, astfel încât cheliceratelor le lipsesc antenele tipice ale multe artropode. De fapt, cheliceratele din fața gurii au doar două anexe care funcționează cel mult doar pentru a însoți mâncarea, dar le lipsește complet structura care ar putea avea o funcție maxilară . [12] [13] Primele anexe de pe partea din față a aparatului bucal se numesc pedipalpi și au funcții diferite în cadrul diferitelor grupuri de chelicerate. [11]
![Structura de bază a corpului artropodelor [14]](http://upload.wikimedia.org/wikipedia/commons/6/66/Arthropod_body_struct_01.png)

Păianjenii și scorpionii aparțin unuia dintre grupurile de chelicerate, numite arahnide . [13] În timp ce chelicerele scorpionilor sunt o pereche modestă de gheare care au funcția de a însoți mâncarea spre gură sau de a o menține nemișcată, [15] cele ale păianjenilor se termină sub formă de colți ascuțiți, care sunt în general otrăvitori; atunci când nu sunt în uz ele sunt îndoite înapoi în partea superioară, în care sunt alocate mustăți groase care au sarcina de a filtra și păstrarea orice bulgări solide, deoarece păianjeni hrănesc numai cu alimente lichide sau transformate în formă lichidă. [7] Pedipalpii din celelalte scorpionide iau forma unor gheare ascuțite capabile să capteze și să țină prada [15] , în timp ce în păianjeni acționează ca senzori de dimensiuni destul de mici, ale căror părți bazale sunt o extensie a gurii . În plus, păianjenii masculi au mărit aceste secțiuni din urmă folosindu-le pentru transferul spermei. [7]
La păianjeni, cefalotoraxul și abdomenul sunt unite de un mic peduncul cilindric care permite abdomenului să se deplaseze independent de cap în timp ce secretă mătase. Partea superioară a cefalotoraxului este acoperită cu o singură carapace convexă, în timp ce partea inferioară este acoperită cu două bucăți de carapace oarecum aplatizate. Abdomenul este moale și are forma ovoidă, nu prezintă niciun semn de subdiviziune sau fracționare, cu excepția Mesotelelor primitive, ai căror reprezentanți actuali, Liphistiidae , au carapace formată din diferite segmente în partea posterioară. [7]
Problema segmentării capului artropodului
![Problema liderului artropodului [16]](http://upload.wikimedia.org/wikipedia/commons/0/07/Arthropod_head_problem_01.png)
O problemă supusă dezbaterii aprinse în rândul experților din sector este aceea referitoare la segmentarea capului artropodelor, care diferă de la un grup la altul; în special, se discută dacă și cum pot fi încadrate aceste diferențe într-un proces evolutiv care se extinde de la trilobiți la crustacee și chelicerate din care păianjenii fac parte. Originile acestor segmentări și prima lor diferențiere se regăsesc în fosilele Burgess Shale , din perioada cambriană , când s-au răspândit o pluralitate de forme de viață foarte diferite, dintre care multe au dispărut în scurt timp. [17] După cum se poate vedea din tabelul din lateral, segmentarea începe de la o structură simplă și liniară, cum ar fi cea a trilobiților, și apoi crește în complexitate, atât în poziționarea diferitelor organe (picioare, pedipalpi, chelicerae), și în creșterea cantității de segmente care, formând inițial parte a capului, în crustacee și trahee, merg să alcătuiască corpul. În sute de milioane de ani, partea de sus, primul segment sau acron , care conține diferitele tipuri de ochi compuși, a rămas aproape neschimbat, la fel și segmentul capului responsabil pentru creșterea antenelor , absent doar în chelicerate. În cele din urmă, este singular cum în Chelicerate și Trahee se pierde un segment posterior acronului în timpul dezvoltării embrionare , respectiv primul și al doilea segment, ca și cum nu ar fi funcționale, continuând să fie latent în aceste grupuri de animale, ca și în cel al omului. apendicele sau coccixul coloanei vertebrale .
Sistemul circulator și respirator
Sistem circulator
La fel ca alți artropode, păianjenii sunt echipați cu celom structurat în zone mici situate în vecinătatea sistemului reproductiv și a sistemului excretor, dar care nu au o funcție circulatorie. Această funcție este îndeplinită de hemocel , o cavitate extinsă pe cea mai mare parte a lungimii corpului și în care circulă fluxul sanguin. Inima are o formă tubulară și este situată în partea din spate a corpului, cu ostii subțiri multiple în interior care permit fluxul de sânge într-o singură direcție, împiedicându-l să curgă înapoi; acest lucru face ca sângele să pătrundă în inimă din hemocel, dar nu se întoarce până nu a trecut prin întreaga parte anterioară; [18] O arteră se ramifică din inimă și se deschide în partea anterioară a abdomenului și care, trecând prin pedicel , se ramifică și se extinde în multe părți ale cefalotoraxului. Deci păianjenii au un sistem circulator deschis. [7]
Sistemul respirator
Păianjenii au dezvoltat mai multe tipuri de organe respiratorii, inclusiv plămâni de carte , trahee sau ambele. Sângele tuturor păianjenilor care au plămâni de carte conține pigmentul respirator hemocianină, ceea ce face transportul oxigenului mai eficient. [13]
Migalomorphic și Mesothelae păianjeni au două perechi de plămâni de carte , care umplu cu hemolimfa , și au două deschideri externe pe suprafața ventrală a abdomenului , care permite aerului să intre și să circule oxigen . Acesta este și cazul uneia dintre cele mai vechi familii de păianjeni araneomorfi , familia Hypochilidae ; familiile rămase din acest grup au doar perechea anterioară de plămâni de carte intacte, în timp ce cea posterioară a sistemului respirator este parțial sau total transformată într-o trahee, prin care oxigenul este difuzat în hemolimfă sau direct în țesuturi și organe. [7] Traheele, acolo unde sunt prezente, sunt foarte asemănătoare ca formă, dimensiune și funcție cu traheele insectelor , dar la păianjeni sunt situate în opistosom. [19] Sistemul respirator cu traheea s-a răspândit probabil de la păianjeni mici pentru a contracara deshidratarea și uscarea. [13] Traheea a fost inițial conectată la organele înconjurătoare printr-o pereche de deschideri numite spiracule traheale , dar astăzi în majoritatea păianjenilor aceste spiracule s-au contopit într-una în mijloc, deplasându-se spre spate lângă filiere . [7] Păianjenii care posedă trahee au, în general, o rată metabolică mai mare și o rezistență mai bună la deshidratare. [20]
Nutriție, digestie și excreție
Nutriție

La păianjeni, chelicerele sunt esențiale pentru nutriție: constau dintr-un segment bazal și un segment terminal în formă de gheară sau colț. O caracteristică unică printre chelicerate este partea terminală a chelicerelor, vârful colțului, care este folosit de majoritatea speciilor pentru a inocula veninul secretat de glandele veninului. [7] Mărimea acestor glande variază în funcție de diferitele familii: în unele lipsesc complet, în altele implică secțiuni mari ale părții anterioare a prosomului. Glandele sunt în general alcătuite dintr-o parte epitelială cu funcție de excreție și sunt înfășurate într-o acoperire ale cărei contracții permit stropirea picăturilor de otravă în interiorul chelicerelor până la baza colților. [21] Singura familie de păianjeni complet lipsită de glande veninoase este cea a Uloboridelor . [22] Cele două pedipalpi sunt mai scurte decât picioarele și formate din șase segmente; la baza au gnathocoxae folosite ca fălci pentru a ține alimentele strâns, ajutând la măcinarea, lichefierea și în final asimilarea acestora. [23]
Digestie
În contextul arahnidelor, [13] păianjenii au un intestin destul de îngust, care poate primi hrană doar sub formă lichidă și două seturi de filtre concepute pentru a păstra orice particulă de hrană solidă, oricât de mică ar fi. [7] Folosesc, unul câte unul, două sisteme diferite pentru a digera alimentele în exterior, înainte de a le ingera.

Unele pompează enzime digestive din mezenter în pradă și apoi aspiră țesuturile lichefiate ale prăzii în intestin; prin urmare, abandonează exuvia goală a prăzii compusă în general din materiale prea dure și nedigerabile. Alte specii de păianjeni macină mai întâi prada pulpând-o cu partea terminală a pedipalpilor și chelicerelor, folosind întotdeauna enzime digestive pentru a facilita asimilarea. La aceste specii atât chelicerele, cât și baza pedipalpilor sunt poziționate astfel încât să formeze o cavitate înaintea gurii propriu-zise. [7]
Stomacul, situat în cefalotorax, funcționează ca o pompă care trimite alimentele tot mai mult în sistemul digestiv. Mezenterul este plin de puncte oarbe pentru digestie, compartimente fără fund, unde substanțele nutritive sunt extrase din alimentele ingerate. Cele mai multe dintre ele sunt situate în abdomen, unde sistemul digestiv este predominant, dar unele sunt situate și în cefalotorax. [7]
Excreţie
Majoritatea păianjenilor transformă deșeurile diverselor activități fiziologice în acid uric , care poate fi excretat ca material uscat. Tubii malpighieni sunt responsabili pentru extragerea acestor produse reziduale din sângele din hemocel și apoi descărcarea lor în bursa cloacală , din care sunt expulzați prin anus . [7] Producția de acid uric și îndepărtarea deșeurilor prin tubuli sunt caracteristici care au evoluat independent în diferite ramuri ale artropodelor pentru a conserva apa și a le permite să supraviețuiască în medii în care este o marfă rară și prețioasă; [24] Dintre diferitele structuri responsabile de excreție, tubulii malpighieni de insecte și arahnide se dezvoltă din părți complet diferite ale embrionului . [13] Cu toate acestea, unii păianjeni primitivi din ordinul Mesothelae și din subordinea Mygalomorphae , păstrează nefridia ( rinichii mici ) ai presupusului artropod ancestral ancestral, [7] ale cărui funcții sunt similare cu cele ale rinichilor umani: o anumită cantitate de apă pentru a expulza deșeurile azotate, cum ar fi amoniacul . [24]
Sistem nervos central
Sistemul nervos central de bază al unui artropod constă dintr-o pereche de mănunchiuri de nervi care rulează de-a lungul intestinului cu grupuri de ganglioni ca centre locale de control în diferitele segmente ale corpului și un creier format prin fuziunea ganglionilor segmentelor a capului plasat în fața și în spatele gurii, astfel încât esofagul să fie înconjurat de acești conglomerate de ganglioni. [25] În special putem distinge un ganglion supraesofagian mai mic , care primește informații de la ocelli și alte organe de simț, și are funcția de a prelucra și integra stimuli externi și apoi, prin neuronii cu care este conectat la celălalt ganglion, coordonează un răspuns. Celălalt ganglion, care ia numele de ganglion subesofagian , situat în partea inferioară a cefalotoraxului , este ansamblul ganglionilor diferitelor anexe și are forma unei stele cu mai multe vârfuri; acest ganglion își continuă drumul în opistosom printr-un mic cordon, inervându-l, iar funcția sa principală este coordonarea motorie a animalului. [26] [27] Cu excepția Mesotelelor primitive, din care Liphistiidae este singura familie care a supraviețuit, păianjenii au sistemul nervos centralizat tipic arahnidelor : ganglionii tuturor segmentelor din partea posterioară a esofagului sunt fuzionați, astfel încât cefalotoraxul să fie în mare parte umplut cu țesut nervos și să nu existe ganglioni în abdomen. [7] [13] [25] Cu toate acestea, în mezotele, ganglionii abdomenului și partea posterioară a cefalotoraxului sunt separate și distincte. [20]
Organele senzoriale
Ocelli

Majoritatea păianjenilor au patru perechi de ochi pe partea din față a cefalotoraxului, poziționate în modele care variază de la o familie la alta. [7] Perechea poziționată pe frunte este de tipul numit ocelli pigment-cup (recipiente de culoare), care în majoritatea artropodelor servesc doar la identificarea direcției din care ajunge lumina, folosind umbra aruncată de suprafețele din ocelli înșiși . În orice caz, ochii principali ai părții frontale a capetelor de păianjen au capacitatea de a forma imagini destul de clare și distincte. [28] [29]
Se crede că ocelii laterali au derivat din ochii compuși ai cheliceratelor mai primitive, dar nu au subdiviziunile fațetelor tipice celor ale altor nevertebrate. La mulți păianjeni acești ocelli secundari detectează lumina reflectată diferit de ochii principali prin intermediul unui tapetum lucidum reflectorizant; de exemplu, acest lucru înseamnă că în întuneric ochii păianjenilor de lup sunt detectați de lumina reflectată a unei torțe datorită tapetelor . Pe de altă parte, ochii secundari ai păianjenilor săritori nu au nicio urmă de tapetum. [7]

Acuitatea vizuală a păianjenilor săritori o depășește cu un factor zece pe cea a libelulelor care sunt, printre insecte, cei care văd cel mai bine. De fapt, ochiul uman, oricât de complex este, este doar de cinci ori mai ascuțit decât cel al unui păianjen sărit. Acești păianjeni pot avea o vedere atât de ascuțită datorită unei serii de lentile, unei retine formate din patru straturi și capacității de a-și întoarce ochii peste tot, integrând imaginile tuturor într-una singură. Singurul dezavantaj al întregului proces de scanare și integrare a imaginii este că este relativ lent. [5]
Cuticula și părul
Așa cum se întâmplă cu alți artropode, cuticula , prin însăși natura sa, ar bloca majoritatea stimulilor senzoriali care vin din exterior, cu excepția cazului în care cuticula în sine este pătrunsă cu organe de simț mici, care conectează exteriorul corpului cu sistemul. De fapt, păianjenii și alte tipuri de artropode au modificat structura cuticulei pentru a putea primi stimuli din exterior și pentru a putea reacționa la ei. Diversi senzori tactili, în special firele de păr de diferite forme numite seturi , sunt sensibili la diferite niveluri de presiune, de la briza ușoară a curenților de aer până la contactul de presiune cu o anumită forță. Alți senzori chimici ne permit să asigurăm funcțiile noastre de recunoaștere a mirosului și gustului , adesea din partea mediană a setelor. [28] Păianjenii au, de asemenea, senzori speciali în articulațiile membrelor lor capabili să perceapă chiar vibrații mici. În construcția pânzelor de păianjen, toate informațiile susținute de acești senzori mecanici și chimici sunt mai importante decât vederea însăși, în timp ce viziunea acută predomină asupra celorlalte organe de simț la vânătoarea activă de păianjeni. [7]
La fel ca în majoritatea artropodelor, păianjenii nu au organe responsabile de echilibru și accelerație și se bazează exclusiv pe vedere pentru a determina din când în când unde este sus și unde este jos. Propioceptorii artropodelor, adică acele organe de simț care poartă informații despre gradul de forță aplicat de mușchi și despre gradul de curbură al diferitelor articulații ale picioarelor, sunt activi și funcționali. Pe de altă parte, se știe puțin despre alte tipuri de senzori interni pe care ar putea avea păianjenii și artropodele. [28]
Locomoţie
Picioare
Păianjenii au patru perechi de picioare funcționale la ambulație, fiecare dintre ele fiind formată din șapte segmente; începând din interior spre exterior sunt: coxa, trohanterul, femurul, rotula, tibia, metatarsul și tarsul. Toate aceste segmente sunt acoperite de o cuticulă prevăzută cu păr, dintre care unele pot fi considerate organe reale ale simțului. [23] Deși toți artropodii folosesc mușchii legați în exoscheletul lor pentru locomoția de a-și flexa membrele, păianjenii și unele grupuri de artropode folosesc un sistem hidraulic de presiune pentru a-i întinde și contracta, un sistem moștenit la rândul său de la formele vieții strămoșilor artropodelor. [3] Ca rezultat, un păianjen cu un cefalotorax străpuns nu mai poate să-și extindă picioarele, iar picioarele păianjenilor morți se contractă singuri, îndoindu-se și agățându-se. [7] Se pare că din unele experimente, păianjenii pot genera presiuni, în interior, de până la opt ori forța necesară pentru a-și extinde complet picioarele, [30] și păianjenii salticidi pot sări în sus până la cincizeci de ori lungimea lor crescând brusc sângele presiune în a treia și a patra pereche de picioare. [7]
Scopule
Majoritatea păianjenilor care vânează activ, mai degrabă decât să se bazeze pe pânze de păianjen, au smocuri dense de păr foarte fin între ghearele lor împerecheate pe vârfurile picioarelor. Aceste smocuri, cunoscute sub numele de scopule , constau din peri ale căror părți terminale sunt împărțite în aproximativ o mie de ramuri, care permit păianjenilor care le posedă să meargă ușor pe suprafețe netede precum sticla și cu capul în jos pe orice tip de tavan. S-a constatat că scopulele obțin acest contact puternic datorită straturilor de apă extrem de subțiri de pe suprafețe. [7] Păianjenii, ca și alți arahnide, atât pe jos, cât și pe alergare, odihnesc întotdeauna cel puțin patru picioare pe pământ în același timp. [31]
Producția de mătase
Lanțurile de aprovizionare

Abdomenul păianjenilor nu are fanere cu excepția celor care au fost modificate pentru a forma unul până la patru ( de obicei trei) perechi de scurt, mobile filierelor , care emit mătase. Ogni filiera ha molti fusuli , sorta di tubicini microscopici, ognuno dei quali è collegato ad una ghiandola sericigena . Sono conosciuti almeno sei tipi di ghiandole setifere, ognuna delle quali produce un tipo leggermente diverso di seta. [7]
La seta è composta principalmente da una proteina molto simile a quella adoperata nelle sete secrete da insetti. Nella fase iniziale di produzione è un liquido che non indurisce subito per esposizione all'aria e ciò consente ai ragni di intrecciarla esternamente. In seguito, dopo che si è indurita, la proteina riesce a cambiare la struttura interna. [32] Una volta completata la filatura, la seta risultante per quanto concerne la resistenza alla trazione , a parità di diametro, è simile al nylon ea materiali biologici quali la chitina , la cellulosa e il collagene , ma è molto più elastica, in altre parole si può stirare e allungare molto di più prima che si spezzi o perda la forma originaria. [7]

Cribellum
Alcuni ragni hanno un cribellum , una filiera modificata con oltre 40.000 fusuli , ognuno dei quali produce una sola fibra eccezionalmente sottile. Tali fibre vengono estratte fuori dal calamistro , una sorta di appendice anatomica dalla forma di pettine con un insieme di peli sulla punta articolata del cribellum, e unite insieme in un solo filo lanoso, utilizzata dal ragno proprio per separare e rendere più voluminosi i fili vischiosi distesi dal cribellum; [33] così distese le fibre sono molto efficienti nell'afferrare e trattenere le setole degli insetti. I ragni più primitivi avevano cribellum che producevano seta in grado di catturare le prede, prima che sviluppassero la seta cosparsa di goccioline appiccicose, più efficiente nel trattenere gli insetti. La maggior parte delle famiglie di ragni ha perso il cribellum e vengono per questo chiamati anche ecribellati . [7]
Alcune specie di ragni che non costruiscono ragnatele per afferrare le prede, utilizzano la seta in diversi altri modi: come involucri per lo sperma e per le uova fertilizzate, come corde di sicurezza nell'affidarsi alle correnti di vento, per costruire tane e ricoveri, e persino come paracadute dagli esemplari più giovani e più leggeri. [7]
Riproduzione e ciclo vitale
Rituali della riproduzione

I ragni si riproducono sessualmente e la fertilizzazione è interna ma indiretta, in pratica lo sperma non è inserito nel corpo della femmina dai genitali del maschio ma da una struttura intermedia. Diversamente da altri artropodi terrestri, [34] i ragni maschi non producono spermatofore, involucri pieni di sperma, all'interno del loro corpo, ma roteano la piccola tela che lo contiene fino all' eiaculazione e poi lo trasferiscono su alcune strutture a forma di pipetta situate sulle punte dei loro pedipalpi . Nel momento in cui un maschio scopre nei suoi pressi segnali della presenza di una femmina della sua stessa specie, controlla se è pronta per l'accoppiamento: ad esempio, in alcune specie che costruiscono ragnatele o corde di sicurezza , il maschio può identificare la specie e il sesso del ragno che ha prodotto questi oggetti attraverso l' odorato . [7]

Corteggiamento
I ragni adoperano generalmente rituali di corteggiamento elaborati per impedire alle femmine, spesso di dimensioni maggiori, di mangiare i maschi prima di portare a termine la fertilizzazione, con le poche eccezioni dove il maschio è di dimensioni tanto più piccole da non essere considerato minimamente appetibile. I maschi si fanno così riconoscere e le femmine, che appaiono fondamentalmente passive non mostrando aggressività, si rendono disponibili all'accoppiamento. Sebbene ogni specie abbia le sue peculiari caratteristiche nel corteggiamento, possiamo però distinguerne tre diversi tipi a seconda della strategia messa in atto da parte del maschio:
- Richiede contatto diretto fra maschio e femmina.
- Richiede feromoni femminili che stimolino il maschio.
- Richiede un riconoscimento visivo da parte della femmina.
Primo tipo
I ragni delle famiglie Thomisidae e Clubionidae hanno un corteggiamento molto modesto; di solito il maschio semplicemente striscia sulla femmina, tiene fermo il suo addome e inserisce l'organo copulatore nell' apertura genitale femminile . Ancora più fugace è l'incontro fra i due sessi di Micrommata virescens , uno Sparassidae , dove il maschio colorato piomba con un salto accanto alla femmina più grande di lui e di colore verde intenso, l'afferra con i cheliceri e, senza tante cerimonie, si posiziona per l'accoppiamento e lo completa con la totale passività della femmina.
Anche molti Haplogynae e migalomorfi mostrano pochi preliminari [35] [36] ; dopo il contatto diretto dei corpi, i maschi e le femmine si scambiano un breve intergioco con le zampe frontali ei pedipalpi prima dell' accoppiamento . Nel caso del terafoside Aphonopelma hentzi , ad esempio, il maschio entra in contatto con la femmina tamburellando con le zampe anteriori sul corpo di lei e collocandosi frontalmente. La reazione della femmina è di difesa disponendosi in posizione di attacco con le zampe anteriori sollevate. Questa particolare posizione consente però di proseguire il corteggiamento al maschio che, tamburellando e tirando, la trasforma in una posizione di sottomissione, in modo da consentirgli l'accoppiamento senza correre pericoli. Curioso è inoltre il fatto che la femmina offra i propri cheliceri aperti al maschio affinché possa tenerli bloccati per il tempo necessario all'accoppiamento, all'incirca un minuto o poco più.
Alcuni ragni muniti di cribellum invece hanno sviluppato un'altra particolare modalità di corteggiamento: i maschi legano le femmine con la seta prima dell'accoppiamento: fatto che sembra avere solo un significato simbolico dato che dopo l'accoppiamento la femmina è perfettamente in grado di liberarsi da sola [37] .

Secondo tipo
Nel caso si renda necessaria l'emissione di feromoni da parte della femmina, il corteggiamento è molto più complesso e diversificato.
Spesso i feromoni sono già secreti dalla femmina sulla ragnatela (riscontrato nei generi Filistata , Segestria e Amaurobius ); in questo caso il maschio cerca di farsi riconoscere tirando o pizzicando fili della ragnatela stessa. A volte il contatto fisico è necessario per scatenare il corteggiamento come in Amaurobius ferox ; invece in Amaurobius simili ed in Amaurobius fenestralis è sufficiente la presenza del feromone. In molte specie di Amaurobius i segnali vibratori generati da zampe, palpi e addome assumono caratteristiche peculiari a seconda della specie.
In alcuni Clubionidae e Gnaphosidae i maschi si sistemano vicino a femmine immature [38] ed aspettano la loro ultima muta per accoppiarsi immediatamente dopo, approfittando del momento in cui queste sono ancora inoffensive in quanto la cuticola non si è ancora completamente sclerotizzata. Questo comportamento è stato riscontrato anche nei maschi del genere Argiope .
Nel caso specifico di Philodromus rufus , un ragno della famiglia dei Thomisidae che vive due anni, la femmina è, occasionalmente, capace di essere fecondata durante l'ultima muta addirittura anche in presenza dei fluidi della muta stessa; inoltre essa si accoppia ripetutamente con lo stesso maschio o anche con altri per più giorni consecutivi; il feromone, in questo caso, è concentrato sulle zampe : infatti se queste vengono lavate con appropriati solventi, non si ha più risposta da parte del maschio [39] . Negli Araneidae i maschi pizzicano i fili delle tele delle femmine in maniera da attirarle sul filo dell'accoppiamento da essi creato.
In Linyphia triangularis , della famiglia Linyphiidae , invece il maschio è sempre dominante rispetto alla femmina e non viene mai da essa attaccato [40] , anzi si nutre sfruttando la tela di lei e poi al momento del corteggiamento la distrugge quasi completamente affinché nessun altro maschio possa raggiungerla e nessuna preda possa distrarre la femmina stessa dalla sua presenza.
Nel caso invece dei Lycosidae e dei Pisauridae le femmine lasciano una traccia di feromoni sul substrato sul quale camminano insieme al filo di seta; infatti occludendo le filiere della femmina ricettiva si è visto che non vi è alcuna reazione da parte dei maschi collocati sullo stesso substrato sul quale è appena passata, mentre si ha una immediata risposta di corteggiamento quando alla stessa femmina si liberano di nuovo le filiere [41] . Sono comunque necessari al corteggiamento anche segnali tattili, vibratori, visivi e acustici. Spesso la femmina non è passiva, risponde attivamente ai segnali maschili. I segnali vibratori ed acustici sono particolarmente necessari in caso di incontri notturni. Il repertorio di segnali del corteggiamento dei Lycosidae è veramente molto complesso e vario.

Nei Pisauridae invece il corteggiamento è curioso per l'approccio da parte del maschio alla femmina tramite un dono, di solito una mosca avvolta nella seta, ma nei più evoluti rimane solo il simbolico involucro vuoto; l'accoppiamento avviene grazie al fatto che la femmina è distratta e pone la sua attenzione principalmente sul dono.
Terzo tipo
Nei Salticidae e negli Oxyopidae i segnali visivi sono i più importanti nel corteggiamento sebbene vi siano anche segnali tattili e chimici [42] . I maschi compiono una vera e propria danza davanti alla femmina e spesso le estremità delle zampe più usate a questo scopo sono addirittura colorate [43] [44] [45] .
Se la femmina rimane immobile o risponde con i segnali adeguati alla specie il maschio la tocca con le zampe anteriori e procede con l'accoppiamento. Pare che le femmine più giovani che abbiano appena mutato siano preferite dai maschi a quelle più vecchie; forse il segnale dei feromoni è più debole in queste ultime. Al riguardo, è interessante il caso di Phidippus johnsoni che cambia completamente sistema di corteggiamento se la femmina è fuori o è nascosta dentro un nido [42] .
Se il corteggiamento ha successo il maschio inietta il suo sperma dai pedipalpi nella femmina attraverso l'apertura genitale della stessa, nota come epigino , posta sulla parte inferiore del suo addome. Gli apparati riproduttivi delle femmine variano da semplici tubi a sistemi che includono ricettacoli seminali nei quali le femmine immagazzinano lo sperma ricevuto e lo rilasciano quando sono fertili. [7]
Accoppiamento

Le molte variazioni delle posizioni dell'accoppiamento si possono riassumere in tre tipi fondamentali ed uno inusuale:
- Caratteristico dei ragni erranti più primitivi Mesothelae , Ortognatha e Haplogynae : il maschio si avvicina alla femmina di fronte, alza il suo prosoma e vi inserisce uno o entrambi i palpi nell'apertura genitale femminile.
- Diffuso fra i ragni che tessono una ragnatela: l'incontro è sempre frontale ma avviene mentre i due partner sono appesi alla tela capovolti. Il maschio inserisce nella femmina i due palpi consecutivamente.
- Riscontrato nei ragni erranti più evoluti: Clubionidae , Lycosidae , Salticidae e Thomisidae : il maschio si arrampica sul prosoma della femmina e si sporge dalla parte in cui vuole inserire il palpo provocando una torsione volontaria della femmina dalla stessa parte.
- In Cheriacanthium , genere dei Clubionidae, viene posta in opera una quarta posizione: i ragni si accoppiano con le superfici ventrali a contatto.
All'interno di una stessa famiglia la posizione dell'accoppiamento non sempre è fissa. Ad esempio negli Agelenidae la posizione n. 3 è quella più comune ma in certe specie di Tegenaria adottano la n. 1.
I sacrifici del maschio
I ragni maschi del genere Tidarren amputano uno dei loro palpi durante la maturazione ed entrano nella vita adulta con un solo palpo. I palpi costituiscono il 20% della massa del corpo degli appartenenti a questo genere, e liberandosi di uno dei due palpi migliorano di molto la mobilità. Nella specie yemenita Tidarren argo, il palpo rimanente è strappato via dalla femmina dopo l'accoppiamento. Il resto di palpo strappato rimane nell' epigino della femmina per le successive quattro ore, a quanto pare continuando a funzionare indipendentemente. Nel frattempo la femmina si alimenta del maschio rimasto senza palpi. [46]
In oltre il 60% dei casi la femmina del ragno australiano Latrodectus hasselti uccide e mangia il maschio dopo che questi ha inserito il secondo palpo nell'apertura genitale della femmina; in questo caso si osserva addirittura che i maschi cooperano cercando di impalarsi sulle zanne della femmina, offrendosi come cibo. Alcune ricerche effettuate hanno mostrato che la maggior parte dei maschi non trova mai un'occasione per accoppiarsi ei pochi fortunati che vi riescono aumentano le probabilità di avere una discendenza assicurandosi che la femmina sia ben nutrita. [47]

Nella norma dei casi, comunque, i maschi di parecchie specie scampano a più accoppiamenti, limitati principalmente dalla loro vita di breve durata. Alcuni riescono a vivere senza correre pericoli anche per qualche tempo nelle ragnatele insieme con le femmine. [48] Come abbiamo visto i maschi che eseguono un rituale di corteggiamento, in alcuni casi (ad esempio la vedova nera ) rischiano comunque di essere divorati dopo l'accoppiamento, mentre in altri i maschi cercano di tenere a bada la femmina eseguendo il rituale di legare e di sottomettere la partner (più che altro simbolico visto che la femmina è ampiamente in grado di liberarsi in qualunque momento). [49]
La deposizione delle uova nel sacco ovigero
Buona parte delle specie della famiglia Pholcidae depone le uova nel terreno in piccole masserelle collegate fra loro, ma possono considerarsi un'eccezione rispetto alla totalità dei ragni. [50]

Infatti la grande maggioranza delle femmine depone fino a 3.000 uova alla volta in uno o più sacchi ovigeri avvolti dalla seta, [7] e si adoperano per mantenerli ad un livello abbastanza costante di umidità . [48]

Il sacco ovigero , ( cocoon è il termine inglese tecnico), viene intessuto dalla femmina in varie forme a seconda della specie: può essere da sferico a emisferico, da cilindrico a fusiforme; alcuni ragni lo ricoprono di terriccio o di foglioline per mimetizzarlo con l'ambiente. Il colore della seta che avvolge il sacco ovigero può essere bianco, rosa, o anche giallo paglierino. Alcuni ragni costruiscono un solo bozzolo in cui più femmine depongono le uova. [50] In alcune specie le femmine muoiono poco dopo la deposizione, ma altre specie proteggono i sacchi ovigeri legandoli alla loro ragnatela o nascondendoli in tane apposite, portandoli nei cheliceri o legandoli alle filiere e trascinandoli con sé. [7]
I piccoli di ragno ( spiderlings )
I piccoli di ragno passano tutti i loro stadi larvali nelle uova e ne escono fuori come ragnetti, molto piccoli e sessualmente immaturi ma di aspetto e forma del tutto simile a quella degli adulti. Alcuni ragni mostrano delle cure parentali per i loro giovani, come ad esempio i ragni-lupo che, rendendo ruvidi i peli del loro dorso, vi covano sopra i piccoli ragnetti che riescono a restare abbarbicati alla madre, [7] mentre le femmine di alcune specie rispondono al comportamento mendicante dei loro giovani dandogli la propria preda o rigurgitando del cibo già ingerito. [48]
Muta dei ragni
Preparativi

Come gli altri artropodi, i ragni effettuano la muta nel crescere in quanto le loro cuticole non sono estensibili. [51] In alcune specie i maschi approfittano di una femmina che ha fatto da poco la muta per accoppiarsi con meno rischio di essere divorati in quanto essa è indebolita per lo sforzo fatto. [48] Nei ragni di dimensioni medio-grandi ci si può accorgere dell'inizio della muta ponendo attenzione ad alcuni particolari: l'animale smette di alimentarsi per un periodo che va da due settimane a qualche mese (dipende dalla specie). Questo prolungato digiuno è segno che il ragno ha accumulato tutte le sostanze necessarie per avviare una nuova fase di accrescimento che culmina con la muta. Inoltre diventa meno aggressivo del solito e le specie che sono solite produrre tela costruiscono una struttura apposita dove riporsi in attesa di questo cambiamento. I dettagli fisici esterni da cui possiamo intuire che è prossima la muta comprendono un' opistosoma più grosso del solito, in evidenza soprattutto se il ragno è giovane, e la perdita dei peli urticanti che lo fa divenire glabro a chiazze. Proprio la colorazione di queste zone ci aiuta a comprendere quando il ragno è prossimo alla muta: quando la colorazione passa dal colore rosa chiaro al grigio scuro, ciò è segno di una forte idratazione della pelle , necessaria perché si possa staccare dall' epidermide sottostante. Appena poco prima della muta alcuni ragni costruiscono con le filiere un tappetino di seta abbastanza fitto; ad esempio, la Lasiodora parahybana vi depone all'intorno i peli urticanti che va perdendo dall'opistosoma al fine di scoraggiare eventuali predatori che volessero attaccarla in questo momento di alta vulnerabilità. [52]
La muta
A questo punto il ragno si sdraia supino in modo da avere la possibilità di muovere più agevolmente le zampe. Poi, grazie ad alcuni enzimi, la parte superiore del cefalotorace si spacca consentendo al ragno di sgusciare fuori dalla vecchia pelle un poco alla volta con movimenti lenti. La pelle dell'opistosoma si spacca lateralmente e quindi vengono rinnovati anche organi esterni come le filiere ei polmoni a libro . Nel caso abbia subito mutilazioni in passato, il ragno è in grado di ricostruire l'arto mancante, anche se inizialmente sarà di dimensioni più piccole degli altri, dimensioni che recupererà una muta dopo l'altra. [53] La frequenza con cui i ragni compiono la muta varia a seconda dell'età, della specie e delle condizioni alimentari: ragni giovani effettuano le mute più di frequente, a volte anche a distanza di un mese, in modo da raggiungere dimensioni che rendano più difficile la predazione da parte di altre specie. Gli esemplari adulti, invece, ne effettuano una o due all'anno, soprattutto a seconda delle condizioni alimentari. I ragni arboricoli e terrestri delle zone tropicali sono più veloci nella crescita e nelle mute avendo una maggiore costanza di alimenti disponibili lungo tutto l'arco dell'anno. [8]
Dimensioni

I ragni sono presenti in natura in una grande varietà di dimensioni. I ragni più piccoli appartengono alla sottofamiglia degli Erigoninae, detti anche ragni nani , la cui lunghezza del corpo è inferiore al millimetro; il Patu marplesi , appartenente alla famiglia Symphytognathidae , e comune nella Samoa Occidentale , ha una dimensione di mezzo millimetro.

I ragni più grandi e più pesanti, invece, sono annoverati fra le tarantole che possono raggiungere un bodylength , cioè una lunghezza del solo corpo, di 90 millimetri ed una legspan , cioè la distanza massima fra le punte delle zampe distese, di oltre 250 millimetri. [54] I ragni più grossi conosciuti appartengono al genere Theraphosa ( Theraphosa blondi e Theraphosa apophysis ) delle foreste della Guyana : sono stati ritrovati degli esemplari che raggiungevano i 30 cm di legspan . Il primato effettivo sembra appartenere ad un esemplare di Theraphosa apophysis (negli anni novanta del secolo scorso era denominata Pseudotheraphosa apophysis , poi il genere dal 2001 è confluito in Theraphosa [55] ) che misurava ben 33 centimetri di legspan . [56]
Per quanto riguarda i ragni fossili alcuni esemplari del genere Megarachne studiati nel 1980 e risalenti al periodo Carbonifero raggiungevano una legspan di ben 80 centimetri; [57] nel 2005, uno studio più accurato ed approfondito su questi esemplari ha portato a classificarli come Eurypterida , classe di artropodi chelicerati aventi alcune affinità con gli Araneae. [58] [59]
Colorazione
Sono state identificate finora solo tre classi di pigmenti nei ragni: gli ommocromi , responsabili di una grande varietà di colori negli occhi, le biline e la guanina , anche se altri pigmenti sono stati scoperti ma non ancora caratterizzati. Le melanine , i carotenoidi e le pterine , che sono molto comuni in altri animali, nei ragni sono completamente assenti. In alcune specie l' esocuticola delle zampe e il prosoma vengono modificate da un processo simile a quello della concia , dando luogo ad una colorazione marrone. [60] Le biline sono state ad esempio rinvenute in Micrommata virescens e sono la causa della sua colorazione verde acceso. La guanina è responsabile per le chiazze bianche del ragno comunissimo Araneus diadematus e in molte specie viene accumulata in cellule specializzate chiamate guanociti . In generi quali Tetragnatha , Leucauge , Argyrodes o Theridiosoma , la guanina è responsabile del loro aspetto argenteo. La guanina, originariamente, è un prodotto finale del metabolismo di una proteina e quindi diviene prodotto di scarto per la maggior parte degli animali. Alcuni ragni, invece, hanno imparato a conservarla ea depositarla nell'organismo per poterla poi utilizzare all'occorrenza. [60] I colori strutturali che si manifestano nelle varie specie, così importanti per il riconoscimento tassonomico di molte di esse, sono il risultato della diffrazione , diffusione e interferenza della luce, ad esempio, nella modifica delle setae e del corpo. Il prosoma bianco dell' Argiope è dovuto a peli che riflettono la luce del Sole; sia il genere Lycosa che Josa hanno aree di cuticola modificata che fungono da piccoli riflettori della luce naturale. [60]
Orifizi
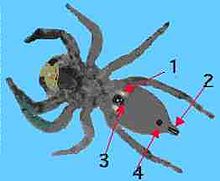
Nei ragni possiamo distinguere i seguenti orifizi:
- La bocca, di piccole dimensioni, è l'apertura fra le gnathocoxae ed è l'unico orifizio che non è posto ventralmente.
- L'orifizio genitale si apre nella parte ventrale dell' opistosoma più vicina al pedicello ; nelle femmine è ricoperto dalla piastra detta epigino .
- Gli ingressi a forma di fessura o lamellare dei polmoni a libro , da due a quattro, sono posti ai lati dell'epigino.
- La parte terminale della trachea , detta stigma tracheale , è posta nella parte centrale dell'opistosoma, poco più in basso dell'orifizio genitale.
- Gli orifizi di forma conica, detti filiere , dai quali viene secreta la seta sono situati nella parte terminale dell'opistosoma.
- L' ano , per l'escrezione dei residui non digeriti, è situato poco sopra le filiere. [19]
Ecologia e comportamento
Nutrizione senza predazione
Normalmente i ragni sono considerati predatori di altri animali, tuttavia esistono alcune eccezioni: ad esempio, il ragno salticida Bagheera kiplingi ottiene il 90% del suo cibo da materiale vegetale abbastanza solido prodotto dalle acacie in mutua relazione simbiotica con una specie di formica del genere Pseudomyrmex . [4]
Le forme giovani (dette juvenilia ) di alcuni ragni delle famiglie Anyphaenidae , Corinnidae , Clubionidae , Thomisidae e Salticidae si alimentano col nettare secreto dalle piante. Studi di laboratorio mostrano che si comportano così intenzionalmente, per lunghi periodi e, mentre si alimentano, provvedono anche ad una pulizia periodica del loro corpo. Questi ragni preferiscono anche soluzioni zuccherine ad acqua semplice, il che indica che sono alla continua ricerca di nutrienti. Siccome molti ragni hanno abitudini notturne è probabile che sia stato sottostimato il loro ricorso al consumo di nettare. Il nettare contiene amminoacidi , lipidi , vitamine , sali minerali oltre a zucchero e vari studi hanno dimostrato che, quando queste integrazioni alimentari sono disponibili, i ragni riescono a vivere più a lungo. Dal punto di vista strettamente ergonomico il ragno, alimentandosi di nettare, evita i rischi di lotte con le prede e il dispendio energetico nel produrre veleno ed enzimi digestivi. [61]
Diverse specie di ragni sono note per alimentarsi di artropodi morti ( saprofagia ), della seta delle ragnatele ed anche delle loro cuticole esterne dopo la muta. Il polline intrappolato nella ragnatela può essere mangiato e vari studi hanno dimostrato che i giovani ragni che hanno potuto nutrirsi di polline hanno migliori possibilità di sopravvivenza. In cattività si hanno diverse notizie attendibili di ragni che si cibano anche di banane , marmellate , salsicce e latte . [61] Tutto ciò è meno insolito di quanto si pensi: fra gli insetti, gli esemplari giovani di una specie di mantide religiosa , si nutrono attivamente di polline, proprio come si comportano alcuni ragni adulti che catturano insetti all'interno dei fiori, ( Thomisidae , ad esempio), cibandosi anche del polline per integrare l'alimentazione. [62]
Tecniche predatorie
- In agguato sulle ragnatele
Il metodo più classico e più famoso di un ragno per catturare la preda è la ragnatela appiccicosa. La grande varietà di disposizioni e strutture delle ragnatele permette a diversi tipi di ragni di intrappolare specie di insetti diversi nella stessa area; ad esempio ragnatele costruite orizzontalmente fra le foglie intrappolano insetti che volano dalla vegetazione interna verso l'alto, mentre ragnatele verticali o poco inclinate consentono di catturare insetti che volano orizzontalmente. I ragni costruttori di ragnatele hanno la vista poco acuta, ma in compenso una sensibilità eccezionale alle vibrazioni della ragnatela stessa. [7]

Le femmine del ragno d'acqua Argyroneta aquatica costruiscono una ragnatela a forma di campana di immersione subacquea che riempiono d'aria ed usano per digerire le prede, mentre fanno la muta, quando si accoppiano e allevano i piccoli. In pratica vivono quasi completamente all'interno di queste campane ; ne escono solamente per cacciare le prede che hanno la sfortuna di impattare con la campana stessa o coi fili che l'ancorano. [63] Alcuni ragni usano le superfici placide di laghi e stagni a guisa di ragnatele : quando due insetti lottando tra loro finiscono sul pelo dell'acqua, il ragno sente le vibrazioni che causano agitandosi e accorre per assalirli. [7]
- Ragnatele mobili e semi-mobili
Vi sono inoltre ragni che tessono piccole ragnatele, aderenti ad una o più zampe, mobili o semimobili, che manipolano per catturare le prede: in pratica distendendo le zampe, la ragnatela si tende in tutta la sua ampiezza; ripiegando le zampe, la tela si affloscia. Quelli del genere Hyptiotes ei ragni della famiglia Theridiosomatidae tendono fortemente le loro tele per poi rilasciarle quando una preda le colpisce, tuttavia non trasportano attivamente la tela con loro, fabbricandola nuova di volta in volta. Le specie della famiglia Deinopidae tessono tele molto piccole, le tengono distese fra le loro due prime paia di zampe e le spingono in avanti fino a due volte la lunghezza del proprio corpo per intrappolare la preda; questa tecnica può aumentare la superficie occupata dalle tele stesse anche di un fattore dieci. Alcuni esperimenti hanno dimostrato che il Deinopis spinosus ha due diverse tecniche per intrappolare la preda: zampate all'indietro per prendere insetti volatori che individua attraverso le vibrazioni e zampate in avanti per afferrare prede che camminano sul terreno. Queste due tecniche sono state osservate anche in altri deinopidi . Gli insetti che camminano sul terreno costituiscono la maggior parte delle prede dei Deinopidae, ma popolazioni di Deinopis subrufus si cibano principalmente di ditteri tipulidi che afferrano menando zampate all'indietro. [64]

Le femmine mature del genere Mastophora costruiscono ragnatele che constano di una singola linea trapezoidale che pattugliano da un lato all'altro. Costruiscono anche una bolas di forma sferica utilizzando un singolo filo che ha la forma di una punta cui è legata una grande palla di seta appiccicaticcia e bagnata al tatto. Emettono composti chimici che sono molto simili ai feromoni delle falene , e poi appendono le bolas alla tela. Anche se approssimativamente il 50% delle loro zampate va a vuoto, riescono a prendere in una notte la stessa quantità di prede di un ragno che tesse una tela orbicolare di uguali dimensioni. Se in circa trenta minuti non hanno catturato una preda con questo sistema, reingeriscono la bolas per poi secernerla di nuovo, dopo essere rimasti per un po' inattivi. [65] [66] Gli esemplari maschi adulti e giovani sono troppo piccoli per costruire le bolas , ma rilasciano feromoni di diverso tipo in grado di attirare falene volanti e le afferrano con le due paia di zampe anteriori. [67]

I primitivi Liphistiidae , i ragni che costruiscono porte-trappola come la famiglia Ctenizidae e molte tarantole, predano tendendo un'imboscata appostandosi in cunicoli spesso chiusi da porte-trappola che impediscono alla preda di fuggire e circondati da reti di fili sottili il cui minimo movimento è percepito dal ragno che le ha costruite proprio per non dare scampo alla malcapitata preda. [20]
- Nascosti in agguato
Altri predatori in agguato, inclusi i Thomisidae detti ragni-granchio , fanno a meno di tali aiuti; [7] alcune specie per predare le api , che hanno un'ottima visione nell' ultravioletto , riescono ad adeguare l'indice di riflessione nell'ultravioletto per mimetizzarsi completamente nei fiori in cui si sono appostate. [60] Essi sono in grado di cambiare colore in pochi giorni per adeguarsi al colore del fiore sul quale si trovano. Lycosidae , Salticidae e Thomisidae catturano la preda inseguendola e fanno principalmente affidamento sulla vista per localizzarla con precisione. [7] Alcuni ragni semi-acquatici, che solitamente vivono vicino a fiumi e stagni nel Nord America, specialmente nelle paludi della Florida , e di dimensioni piuttosto grandi, si ancorano con le zampe posteriori ad una pietra o una pianta, e tengono quelle anteriori a filo d'acqua in attesa della preda talvolta costituita da piccoli pesci d'acqua dolce, come il Gambusia affinis , che riescono a catturare, portare fuori dall'acqua e tramortire con le neurotossine prima di mangiarli. [68]
- Tattiche predatorie istintive

Fra i ragni salticidi quelli del genere Portia sembrano cacciare usando una certa intelligenza, [5] aggirando le loro vittime o adescandole fuori dalle loro ragnatele. Studi di laboratorio hanno mostrato che le tattiche istintive adoperate da questi ragni sono punti di partenza per approcci del tipo prova e sbaglia , che consentono loro di imparare molto rapidamente come affrontare prede sempre diverse. [69] Il loro atteggiamento porta comunque a ritenerli pensatori relativamente lenti, cosa che non deve certo sorprendere in quanto i loro cervelli sono enormemente più piccoli di quelli dei mammiferi che adottano tecniche di caccia non molto più sofisticate. [5]
- Mimetizzazione

I ragni del genere Myrmarachne , detti anche ragni imitatori di formiche , per adattarsi a questo ruolo hanno subìto modifiche decisive: sviluppano addomi più magri e pseudo-vite nel cefalotorace per imitare nel modo più preciso possibile le tre regioni distinte del corpo di una formica ; tengono sempre puntato verso l'alto il primo paio di zampe per imitare l'aspetto delle antenne di questi insetti, particolare anatomico che nei ragni manca del tutto, e nello stesso tempo nascondono alla formica incontrata di avere otto zampe invece di sei; inoltre sviluppano un paio di occhi rotondi di colore uniforme per nascondere di averne otto o sei semplici, mentre le formiche ne hanno due composti; infine ricoprono il loro corpo con peli che riflettono la luce per assomigliare ai corpi generalmente lucidi delle formiche. In alcune specie di ragni i maschi e le femmine imitano specie diverse di formiche, anche perché usualmente le femmine dei ragni sono di dimensioni maggiori dei maschi. Queste specie di ragni cambiano sovente anche il loro comportamento per adeguarlo maggiormente a quello delle formiche e farsi accettare come uno di loro; ad esempio molti adottano un modello di movimento a zig-zag, se hanno la capacità di saltare evitano di farlo in presenza delle formiche; ei ragni del genere Synemosyna camminano sull'orlo esterno delle foglie allo stesso modo delle formiche del genere Pseudomyrmex . In molti ragni e artropodi l'adattamento ad imitare le formiche può anche servire da deterrente visivo nei confronti di vari predatori quali uccelli, lucertole e altre specie di ragni. Buona parte di queste specie comunque preda direttamente le formiche o le parassita come se fossero afidi . Ancora da menzionare il comportamento dei ragni del genere Amyciaea , anch'essi mirmecofagi , che, per quanto dissimili dalle formiche Oecophylla, mentre sono in caccia sanno imitare perfettamente la posa morente di quelle formiche, attirando così l'attenzione delle formiche operaie che appena si avvicinano vengono predate. Dopo l'uccisione di una formica alcuni di questi ragni la tengono fra loro e le altre formiche per evitare di essere riconosciuti come estranei e quindi attaccati. [70]
Tattiche difensive
Colorazione mimetica

Si ritiene che la colorazione dei ragni sia un camuffamento che li aiuti ad evitare i loro predatori più pericolosi quali uccelli e vespe parassite, i quali hanno una buona visione dei colori. La maggior parte delle specie dei ragni è colorata in modo da mascherarsi bene nel fogliame o nel terreno e attirare poco l'attenzione; alcune specie, al contrario, hanno una colorazione accesa con strisce e macchie che tendono a separare i contorni dell'animale e darne una visione non conforme. In alcune specie come il ragno Theridion grallator , endemico delle isole Hawaii , molti schemi di colorazione sono presenti in un rapporto che sembra rimanere costante, e questo costituisce senz'altro una difficoltà per il predatore nel riconoscere chiaramente l'animale. La maggior parte dei ragni non sono pericolosi o di sapore sgradevole o repellente, però la colorazione sgargiante che esprimono offre loro un certo beneficio. Comunque le specie in possesso di veleno mortale, mascelle potenti e peli urticanti hanno colori alquanto vistosi e, se minacciati, non esitano a metterli in mostra. [60] [71]
Peli urticanti
Vari ragni della famiglia Theraphosidae , comprese le tarantole e la sottofamiglia Harpactirinae , hanno peli urticanti sui loro addomi ed usano le zampe per dare un leggero colpetto e scagliarli verso gli assalitori. Questi peli sono composti da setole molto fini con basi fragili e una fila di piccole barbe sulla punta. Queste barbe provocano irritazione intensa soprattutto se colpiscono parti delicate quali occhi o mucose ma non vi è alcuna prova che contengano veleno. [72] Alcuni ragni si difendono dalle vespe includendo nelle loro ragnatele dei fili molto robusti, consentendo al ragno stesso di fuggire mentre la vespa è impegnata a strappare questi fili più tenaci. [73] Il ragno di colore dorato Carparachne aureoflava , originario del Deserto del Namib , mette in fuga le vespe parassite muovendosi a scatti dal posto in cui si trova e facendo la ruota sulle dune di sabbia circostanti. [74]
Longevità nei ragni
La maggior parte dei ragni vive solamente per uno o due anni, anche se alcune tarantole in cattività hanno raggiunto i venticinque anni [7] . Per quanto concerne i ragni di una certa dimensione, che sono anche quelli più tenuti sotto controllo in quanto vengono allevati dagli appassionati oltre che studiati dai ricercatori, si è appurato che di norma i maschi sono meno longevi delle femmine: appena giunti alla maturità sessuale non possono più effettuare mute e il loro ciclo vitale giunge a compimento. Le femmine invece, dopo la maturità sessuale, continuano a crescere e ad effettuare mute. La morte sopraggiunge o per problemi metabolici o per eccessivi sforzi profusi per la muta. In termini numerici il record documentato di un ragno maschio è di massimo cinque anni, mentre le femmine di norma vivono circa quindici anni: anche qui i record documentati portano un massimo di oltre quarant'anni [75] . Altro segno distintivo notato è che i ragni mediamente più longevi sono quelli terrestri che abitano le zone con maggiore escursione termica, il che li obbligherebbe ad alternare periodi di attività con periodi di latenza; mentre quelli meno longevi sono i ragni arboricoli delle zone tropicali, calde e umide tutto l'anno. [76]
Socialità nei ragni
Alcune specie di ragni che tessono tele vivono insieme in grandi colonie e mostrano comportamento sociale, anche se non nel complesso, tutti insieme, come accade fra gli insetti sociali . Il ragno Anelosimus eximius , della famiglia Theridiidae può costituire colonie di oltre 50.000 individui. [77] Il genere Anelosimus ha proprio una forte tendenza verso la socialità : tutte le specie americane note sono sociali e una specie del Madagascar ha un comportamento che è avvicinabile a quello sociale. [78] Membri di altre specie, appartenenti alla stessa famiglia, ma di generi diversi, hanno sviluppato indipendentemente comportamento sociale, in casi di convergenza evolutiva . Ad esempio, anche se il Theridion nigroannulatum appartiene ad un genere che non ha altre specie sociali affini, costruisce delle colonie che possono contenere molte migliaia di individui che cooperano nella cattura delle prede e nella spartizione del cibo. [79] Fra gli altri ragni sociali possiamo annoverare la maggior parte delle specie di Philoponella , appartenente alla famiglia Uloboridae ; la Agelena consociata , della famiglia Agelenidae e Mallos gregalis della famiglia Dictynidae . [80] I ragni predatori sociali hanno bisogno di difendere le loro prede dai cleptoparassiti [81] , in pratica da altri ragni ladri , e le colonie quanto più sono grandi, tanto più hanno successo in questo. [82] Il ragno vegetariano Bagheera kiplingi vive in piccole colonie in cui si aiutano reciprocamente nella protezione delle uova e dei piccoli. [4] Persino i ragni del genere Latrodectus , fra i quali si annovera la temibile vedova nera, notoriamente aggressivi e cannibali, in cattività formano piccole colonie, condividendo la ragnatele e il cibo predato. [83]
Tipi e forme di ragnatele
Caratteri generali

Gli studi finora condotti non hanno mostrato correlazioni fra la classificazione dei ragni e le tipologie di ragnatele che essi costruiscono: nell'ambito dello stesso genere, specie diverse possono costruire ragnatele dello stesso tipo o alquanto diverse. Non vi è neppure corrispondenza fra la classificazione dei ragni e la composizione chimica delle loro tele. La convergenza evolutiva nella costruzione delle ragnatele, in altre parole l'uso di tecniche di costruzione simili da parte di specie fra loro geneticamente lontane, è alquanto frequente. Le ragnatele costruite in forma orbicolare ei comportamenti ripetitivi e spiraliformi che occorrono per edificarle non hanno finora ricevuto la giusta attenzione da parte degli aracnologi , nonostante il fatto che questo tipo di ragnatela rappresenti il costrutto della maggior parte dei ragni. La sequenza di costruzione basilare raggio-spirale presente nelle ragnatele di forma orbicolare e spiraliforme, e l'elevato senso della direzione necessario per edificarle, è stato sicuramente ereditato da più famiglie di ragni attraverso antenati comuni. [84] Vari studiosi ritenevano che le ragnatele orbicolari appiccicose fossero un'innovazione evolutiva che ha dato luogo alla differenziazione degli Orbiculariae. Da ricerche più recenti sembra invece che i ragni che costruiscono tele di forma tubolare siano un sottogruppo che si è evoluto da quelli che la edificano di forma orbicolare e che i primi contano il 40% di specie in più e sono quattro volte più numerosi dei secondi. Il loro maggiore successo evolutivo potrebbe essere dovuto al fatto che le vespe sfecidi , che sono i principali predatori dei ragni, di frequente preferiscono attaccare i ragni che costruiscono ragnatele orbicolari o spiraliformi. [85]
Ragnatele orbicolari o spiraliformi

Le ragnatele di tipo orbicolare sono le più diffuse; hanno forma pressoché circolare e, partendo dai punti di appoggio da dove il ragno comincia a costruirle, si sviluppano a spirale verso il centro. Una ragnatela deve assolvere queste tre funzioni: intercettare la preda; assorbire l'urto della preda stessa senza rompersi; intrappolare l'animale, lasciandolo impigliato e invischiato nella tela, trattenendolo fino all'arrivo del ragno. Non vi è una forma o disegno di ragnatela ottimale per ogni preda. Ad esempio, una spaziatura più larga fra le linee circolari della tela ne aumenterà certamente l'area e la sua abilità nell'intercettare la preda, ma a scapito della capacità di trattenerla per il tempo necessario; nel caso di una spaziatura più serrata fra le linee, le gocce appiccicose migliorano senz'altro la trattenuta della preda, ma ne rendono più facile l'individuazione a distanza e quindi la possibilità di evitarla da parte dei malcapitati animali, almeno durante il giorno e comunque l'area occupata diviene molto minore. All'incirca la metà delle potenziali prede che incappano in una ragnatela orbicolare riesce a scappare. Non vi sono sostanziali differenze fra la struttura delle ragnatele costruite durante il giorno o durante la notte. In effetti non esiste alcuna correlazione semplice fra il tipo di ragnatela costruita e la tipologia di preda che vi rimane intrappolata, in quanto ogni tipo di ragnatela orbicolare è in grado di catturare una varietà cospicua di prede. [84]
I punti di snodo ( hubs ) delle ragnatele orbicolari, dove in genere si appostano i ragni, solitamente sono posti verso il centro, in modo che il ragno possa spostarsi velocemente in qualunque direzione e afferrare la preda nel minor tempo possibile. Se nella tela vi è una direzione preferenziale verso la quale il ragno può scappare dai suoi predatori, il punto di snodo dove si apposta è collocato nei pressi di questa. [84]
Le ragnatele orbicolari orizzontali sono le più diffuse nonostante siano meno efficienti nell'intercettare e nel trattenere la preda e più vulnerabili all'azione della pioggia battente o di ramoscelli e foglie che vi cadano sopra. Alcuni ricercatori sostengono che queste ragnatele orizzontali offrano dei benefici compensativi quali: vulnerabilità ridotta ai danni che può causare il vento ; la visibilità ridotta per le prede che volano verso l'alto e vi incappano, in quanto rivolgono la schiena al cielo; la possibilità di farle oscillare per catturare insetti in volo orizzontale lento. Non vi è comunque un singolo motivo che giustifichi la grande diffusione di questo tipo di ragnatela rispetto a quella verticale. [84]

I ragni, soprattutto quelli della sottofamiglia Argiopinae , spesso intrecciano bande e nastri di seta in varie parti della ragnatela, rendendola di fatto più visibile e appariscente, che prendono il nome di decorazioni o stabilimenta . Ricerche sul campo hanno dimostrato che queste ragnatele con varie bande decorative catturano un maggior numero di preda per ogni ora, anche se non ne è ben chiaro il motivo. [86] In ogni caso successivi studi condotti in laboratorio hanno dimostrato che i ragni riducono la costruzione di queste decorazioni se sono consapevoli della vicina presenza di potenziali predatori. [87]
Vi sono molte varianti insolite di ragnatele orbicolari, alcune delle quali fenomeno di convergenza evolutiva : ragnatele costruite a pelo d'acqua, allo scopo di intrappolare gli insetti che volano a poca distanza dalla superficie; oppure quelle con ramoscelli passanti per il loro centro, presumibilmente per nascondere i ragni dai predatori; infine ragnatele a forma di scala , che sembrano molto efficienti per cacciare falene. In ogni caso il senso di molte di queste varianti a volte appare poco chiaro. [84]
Ragnatele dalla forma tubolare e cobweb

I ragni delle famiglie Ctenizidae , Liphistiidae , Barychelidae , Cyrtaucheniidae , Nemesiidae e Idiopidae costruiscono delle particolari tele tubolari a porta-trappola , ossia con un'apertura più o meno mimetizzata nel terreno o fra le foglie, dalla quale il ragno può sbucare all'improvviso per catturare la preda e trascinarla all'interno. In genere estendono tutt'intorno alla porta-trappola filamenti di seta, al fine di poter controllare, standosene ben nascosti, la direzione e la velocità della preda che sta sopraggiungendo, tramite le vibrazioni che essa produce nel camminare sopra questi filamenti.
I ragni della famiglia Theridiidae sono noti anche per la tessitura irregolare, ovoidale o intricata delle loro ragnatele tridimensionali, comunemente chiamate cobweb . Questo comportamento sembra portare vantaggi dal punto di vista evolutivo in quanto consente di ridurre la quantità di seta appiccicosa usata, fino a portare, in qualche specie, alla completa assenza di fili appiccicaticci. La costruzione di queste ragnatele è meno organizzata di quelle orbicolari e può richiedere diversi giorni, a seconda del grado di complessità. [85]
Altri tipi di ragnatele

I ragni della famiglia Linyphiidae generalmente costruiscono tele orizzontali di forma disuguale somiglianti ad una superficie a maglie intrecciate, con piccoli grovigli di seta sparsi qua e là. Gli insetti che colpiscono questi grovigli cadono sulla sottostante rete a forma di amaca che trasporta le vibrazioni fino al ragno in attesa, e vengono trattenuti e immobilizzati finché non sopraggiunge il ragno che sta in agguato al di sotto. [88] La costruzione di questo tipo di tela inizia con la deposizione dei fili che costituiranno la periferia e l'intelaiatura della tela stessa; il ragno li dispone prima sul piano orizzontale, fino a definirne i margini del foglio a forma di amaca , poi colloca i fili verticali che formeranno il supporto del foglio stesso. Quindi su questa struttura di base il ragno continua a tendere fili da un capo all'altro rendendola sempre più intrecciata, badando che i filamenti principali, detti di ancoraggio, tengano sempre ben tesa tutta la tela, evitando che si afflosci. Tutto questo lavoro fa sì che non esistano praticamente due tele uguali fatte da questi ragni, dipendendone la struttura dalla collocazione, sempre variabile, dei punti di sostegno e dalla grandezza del ragno. Non essendo i filamenti attaccaticci, quando una mosca o una cavalletta va a sbattere contro i filamenti, nello staccarsene finisce sul foglio a maglie intrecciate sotto il quale è appostato il ragno che prontamente interviene e morde il malcapitato animale. [89]
Ragnatele nello spazio
Nel 1973 lo Skylab 3 prese a bordo, a scopo sperimentale, alcuni ragni per studiarne il comportamento nello spazio e come tessono le ragnatele in assenza di gravità. Nei primi tentativi produssero ragnatele poco consistenti e disorganizzate, poi, col passare del tempo, si adattarono al nuovo ambiente mostrando grandi capacità di recupero. [90]
Ragni fossili

I ragni fossili sono una vera rarità in quanto il loro corpo è molle e non si preserva nel tempo se non in casi eccezionali. Il più vecchio fossile noto è un' ambra che contiene artropodi fossili risalenti a circa 130 milioni di anni fa, del periodo Cretaceo . [91] Oltre a preservare i dettagli anatomici dei ragni con molti particolari, alcune ambre hanno catturato anche istanti di accoppiamento fra ragni, uccisioni di prede, produzioni di seta e, probabilmente, cura dei piccoli. In qualche raro caso l'ambra ha preservato i sacchi ovigeri dei ragni e le ragnatele, alcune persino con la preda intrappolata. La ragnatela fossile più grande scoperta finora ha circa 100 milioni di anni. [92] Si sono conservati anche resti fossili di alcuni ragni nei lagerstätten , luoghi dove le condizioni di seppellimento e di conservazione eccezionalmente consentono di mantenere integra parte dei tessuti molli. [93]

L'aracnide fossile più antico conosciuto è il trigonotarbide Palaeotarbus jerami , di circa 420 milioni di anni fa, del periodo Siluriano , che aveva un cefalotorace di forma triangolare, l'addome segmentato, otto zampe ed un paio di pedipalpi . [94] Il fossile originariamente venne chiamato Eotarbus , ma venne rinominato quando ci si rese conto che un altro aracnide del Carbonifero era già stato chiamato Eotarbus in precedenza: [95] Attercopus fimbriunguis , risalente a 386 milioni di anni fa, nel periodo Devoniano , mostra il primo fusulo per produrre la seta conosciuto e venne perciò identificato come ragno. [10]
In ogni caso questi fusuli erano posti sulla parte inferiore dell'addome piuttosto che su filiere , che consentono con le loro appendici modificate una certa mobilità nel difficile compito di costruire le ragnatele. Per questo motivo i ragni del genere fossile Attercopus e l'aracnide simile del periodo Permiano , il Permarachne , non possono essere considerati dei ragni veri e propri; probabilmente adoperavano la poca seta che riuscivano a filare per ricoprire e proteggere il nido o le uova, piuttosto che per tessere ragnatele. [96] Nel periodo Carbonifero la maggior parte dei ragni apparteneva al sottordine Mesothelae , ragni piuttosto primitivi, la cui unica famiglia rappresentata oggi è quella dei Liphistiidae . [10] Il mesotelide Paleothele montceauensis , del tardo Carbonifero di oltre 299 milioni di anni fa, aveva ben cinque filiere . [97]

Nonostante nel periodo Permiano , all'incirca fra 251 e 299 milioni di anni fa, vi sia stata una rapida diversificazione di insetti volanti, sono ben pochi i resti fossili di ragni rinvenuti appartenenti a quest'epoca. [10]
I gruppi principali in cui si suddividono i ragni moderni, Mygalomorphae e Araneomorphae , hanno fatto la loro prima apparizione nel periodo Triassico , all'incirca 200 milioni di anni fa. Alcuni migalomorfi del Triassico sembrano essere membri della famiglia Hexathelidae , che include le moderne tarantole ei famosi ragni australiani dalla tela a imbuto, le cui filiere sembrano proprio essersi adattate per la costruzione di ragnatele sagomate a forma di imbuto atte a catturare principalmente gli insetti saltatori che, una volta caduti all'interno, non ne possono più uscire. Gli Araneomorphae comprendono la grande maggioranza delle specie e famiglie di ragni moderni, inclusi quelli che tessono la familiare ragnatela a forma orbicolare. Il periodo Giurassico e il periodo Cretaceo offrono una grande quantità di fossili, inclusi parecchi rappresentanti di famiglie moderne. [10]
Filogenesi e tassonomia
Albero filogenetico dei Chelicerati
| Chelicerata |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
zoologo JWSchultz del 2007. [98]
Il simbolo †; indica che l'ordine è estinto.
Fra i ricercatori del settore vi è un sostanziale accordo nel definire i ragni un gruppo monofiletico , ovvero appartenenti ad un gruppo che contiene un antenato comune a tutti i suoi discendenti. [98] Comunque c'è stato un acceso dibattito su quali possano essere i loro parenti evolutivi più vicini e su come tutti questi evolsero dai chelicerati ancestrali che erano animali marini. Il cladogramma sulla destra è basato sulle analisi dell'aracnologo JWShultz del 2007. Gli altri punti di vista principalmente si possono riassumere in due schemi:
- gli scorpioni sarebbero apparentati più agli estinti scorpioni marini detti euripteridi che ai ragni;
- i ragni e gli Amblypygi insieme formano un gruppo monofiletico.
L'aspetto alquanto intricato di molte diramazioni dell'albero evolutivo di questi animali mostra palesemente che vi sono ancora fondamentali incertezze circa le relazioni che sussistono o sono sussistite fra i gruppi coinvolti. [98]
Agli aracnidi mancano diverse caratteristiche presenti negli altri chelicerati, inclusa la possibilità di voltarsi all'indietro spalancando bocche e gnatobasi ( mascelle articolate ) alla base delle proprie zampe; [98] entrambe queste comuni caratteristiche sono parte del sistema di alimentazione dell'artropode ancestrale. [99] Invece loro hanno bocche che si spalancano in avanti e verso il basso e tutti hanno dei sistemi per respirare aria. [98] I ragni, appartenenti all'ordine Araneae, si distinguono dagli altri gruppi di aracnidi per varie caratteristiche, comprese le filiere e, nei maschi, i pedipalpi che hanno sviluppato un adattamento specializzato nel trasferimento dello sperma. [100]
Tassonomia
I ragni sono suddivisi in tre sottordini, Mesothelae , Mygalomorphae e Araneomorphae ; secondo gli ultimi orientamenti tassonomici degli aracnologi questi ultimi due vanno raggruppati nel sottordine Opisthothelae ed in questo caso diverrebbero infraordini come si evince dall'albero tassonomico a lato. La ragione di questa suddivisione è per differenziare specificamente i Mesothelae, ragni dalla struttura e dall'anatomia più primitiva, dagli Opisthothelae, che racchiudono ragni con caratteri fisiologici più evoluti e più moderni, come l'assenza dei gangli e la più efficiente disposizione delle zanne.
A maggio 2017 sono state identificate e classificate dagli aracnologi circa 46.738 specie di ragni raggruppate in 112 famiglie e ben 4.058 generi . [1]
| Suddivisione dei ragni [100] | Tratti distintivi | ||||||
| Sottordine | Specie | Generi | Famiglie | Segmenti appiattiti in cima all'addome [101] | gangli addominali | filiere [101] | Singolare direzione delle zanne [7] |
|---|---|---|---|---|---|---|---|
| Mesothelae | 96 | 8 | 1 | Si | Si | Quattro o più paia sotto la parte mediana dell'addome | In giù e in avanti |
| Mygalomorphae | 2.865 | 351 | 16 | No | No | Due o tre paia sotto la parte posteriore dell'addome | |
| Araneomorphae | 43.777 | 3.699 | 95 | Dai lati verso il centro, simili a pinze | |||
Mesothelae

Le uniche specie viventi del primitivo ordine Mesothelae appartengono alla famiglia Liphistiidae , rinvenute solamente in Asia sudorientale , Cina e Giappone . [100] La maggior parte della struttura delle ragnatele a forma di cunicoli a porta-trappola intessute dai Liphistiidae , hanno la forma di tunnel di seta mimetizzati, caratterizzati da una seconda porta-trappola adoperata come uscita di emergenza. Specie del genere Liphistius secernono fili di seta tutt'intorno all'apertura della porta-trappola al fine di rendersi conto prontamente dell'avvicinarsi di una preda, mentre le specie del genere Heptathela non sprecano tutta questa seta e contano invece sui propri sensori incorporati nelle zampe per accorgersi subito dell'approssimarsi di una potenziale preda. [103] I ragni del genere Heptathela sono gli unici, fra i Mesothelae, a non possedere ghiandole velenifere. [104]
Le famiglie estinte degli Arthrolycosidae , i cui esemplari sono stati rinvenuti in rocce dei periodi Carbonifero e Permiano , e degli Arthromygalidae , finora rinvenuti solo in rocce del Carbonifero , sono state classificate come appartenenti ai Mesothelae. [105]
Opisthothelae
La maggioranza degli aracnologi sembra essere d'accordo nel raggruppare gli Araneomorphae ei Mygalomorphae in un unico sottordine, denominato Opisthothelae , per designare quei ragni che presentano caratteri più evoluti rispetto ai Mesothelae , dotati di caratteristiche più primitive. Ciò ovviamente fa sì che gli araneomorfi ei migalomorfi siano classificati come infraordini . [106]
Mygalomorphae

I Mygalomorphae , che fanno la loro prima apparizione in rocce fossili del periodo Triassico , [10] sono generalmente ben tozzi e pelosi, con grandi e robusti cheliceri e zanne. [100] Esempi molto noti comprendono le tarantole, i ragni che costruiscono le tane a porta-trappola e quelli delle ragnatele a forma di cunicolo, imbutiformi, dell' Asia sudorientale e dell' Australia . [7] Una buona percentuale di essi passa la maggior parte del tempo nel costruire queste elaborate ragnatele tessendo fili su fili, cercando di ampliare la zona di cattura della preda; alcuni costruiscono ragnatele atte a catturare direttamente la preda. In ogni caso i migalomorfi non possono secernere la seta piriforme, a forma di fuso, che gli araneomorfi usano per far aderire i fili della tela in tensione alla maggior parte delle superfici, coprendo aree più ampie; il non poter usare questa tecnica rende più difficile e laboriosa l'edificazione della tela per i migalomorfi. Poiché questi ultimi raramente si disperdono nei dintorni facendosi trasportare dalla corrente, le loro popolazioni spesso formano gruppi molto folti. [100] Oltre ad altri artropodi, i migalomorfi predano rane , lucertole e lumache . [107]
Araneomorphae


Gli araneomorfi da soli rappresentano oltre il 90% delle specie conosciute di ragni, includendo i ragni dalle tele orbicolari o spiraliformi, i ragni salticidi ei ragni corridori , [100] e anche l'unico ragno vegetariano noto, e quindi stili di vita e comportamenti molto differenti fra loro. [4] Sono facili da riconoscere per la posizione dei cheliceri, fronteggiati l'uno con l'altro, il che consente loro un'ampia varietà di prese che congiunta ad una grande capacità di adattamento agli ambienti più disparati, fa sì che siano i ragni più diffusi nel mondo e persino nei luoghi antropizzati, dalle case (soffitte, cantine, ripostigli) ai giardini (cespugli, alberi, orticelli). [108]
I ragni in Italia
L'Italia, per la varietà di habitat che possiede lungo tutto l'arco peninsulare, ha una buona biodiversità aracnologica.
Al 2004 sono stati censiti 375 generi di 49 differenti famiglie, per complessive 1534 specie rinvenute nel territorio della nazione. [109]
Al marzo 2015, nella Checklist dei ragni italiani, risultano sul territorio italiano 1599 specie appartenenti a 422 diversi generi di 54 famiglie di ragni. Le sottospecie identificate sono 26, da riferirsi a 13 specie diverse [110]
Considerazioni analitiche
Una disamina di questi ultimi dati per sottordine ha evidenziato:
- sottordine Mesothelae : sono completamente assenti in territorio italiano [109] [110] .
- sottordine Migalomorfi : in Italia sono risultati piuttosto rari, e rappresentati da 6 famiglie: Atypidae (3 specie), Cyrtaucheniidae (4 specie), Ctenizidae (2 specie), Hexathelidae e Theraphosidae (1 specie), ed è cospicua, in paragone, la presenza dei Nemesiidae (18 specie [110] o 20 specie [109] ).
- sottordine Araneomorfi : con 48 famiglie ampiamente rappresentate, è una vera esplosione di biodiversità [110] :
Nelle tabelle seguenti sono riportate le famiglie rinvenute in territorio italiano col numero di generi e specie:
| Migalomorfi reperiti in Italia [110] | ||
|---|---|---|
| Famiglia | Generi | Specie |
| Atypidae | 1 | 3 |
| Ctenizidae | 1 | 2 |
| Cyrtaucheniidae | 1 | 4 |
| Hexathelidae | 1 | 1 |
| Nemesiidae | 2 | 18 |
| Theraphosidae | 1 | 1 |
| TOTALI | 7 | 31 |
| Araneomorfi reperiti in Italia [110] | ||
|---|---|---|
| Famiglia | Generi | Specie |
| Agelenidae | 13 | 62 |
| Amaurobiidae | 2 | 12 |
| Anapidae | 2 | 3 |
| Anyphaenidae | 1 | 3 |
| Araneidae | 22 | 59 |
| Clubionidae | 1 | 30 |
| Cybaeidae | 2 | 6 |
| Dictynidae | 12 | 28 |
| Dysderidae | 10 | 79 |
| Eresidae | 2 | 5 |
| Eutichuridae | 1 | 14 |
| Filistatidae | 2 | 3 |
| Gnaphosidae | 27 | 154 |
| Hahniidae | 4 | 17 |
| Leptonetidae | 3 | 8 |
| Linyphiidae | 141 | 465 |
| Liocranidae | 9 | 23 |
| Lycosidae | 15 | 103 |
| Mimetidae | 2 | 6 |
| Miturgidae | 2 | 9 |
| Mysmenidae | 3 | 3 |
| Nesticidae | 3 | 7 |
| Oecobiidae | 2 | 4 |
| Oonopidae | 5 | 10 |
| Oxyopidae | 1 | 4 |
| Palpimanidae | 1 | 1 |
| Philodromidae | 4 | 40 |
| Pholcidae | 5 | 7 |
| Phrurolithidae | 2 | 6 |
| Pimoidae | 1 | 1 |
| Pisauridae | 2 | 6 |
| Prodidomidae | 1 | 1 |
| Salticidae | 39 | 135 |
| Scytodidae | 1 | 2 |
| Segestriidae | 2 | 5 |
| Selenopidae | 1 | 1 |
| Sicariidae | 1 | 1 |
| Sparassidae | 3 | 5 |
| Synaphridae | 1 | 1 |
| Tetragnathidae | 4 | 19 |
| Theridiidae | 34 | 108 |
| Theridiosomatidae | 1 | 1 |
| Thomisidae | 14 | 76 |
| Titanoecidae | 2 | 6 |
| Trachelidae | 4 | 4 |
| Uloboridae | 3 | 5 |
| Zodariidae | 2 | 19 |
| Zoropsidae | 1 | 3 |
| TOTALI | 415 | 1568 |
Al 2003, quando erano state rinvenute in Italia 1411 specie, ben 213 di esse erano endemismi , presenti esclusivamente sul territorio italiano in una o più località [111] .
I ragni e l'uomo
Morsi di ragno


La stragrande maggioranza dei ragni morde un essere umano solo in casi estremi di autodifesa e se viene direttamente minacciato; comunque, quasi nella totalità dei casi, l'irritazione è poco più grave di un morso di zanzara o della puntura di un' ape . [112] Alcune specie procurano invece morsi abbastanza seri dal punto di vista medico, come vari generi della famiglia Sicariidae , principalmente i Loxosceles , e le vedove nere , che mordono solo se si sentono minacciati direttamente e in modo insistente, anche se ciò può capitare incidentalmente. [113] [114] Gli araneomorfi , e in particolare gli appartenenti alla famiglia Agelenidae , adoperano tattiche difensive di tipo aggressivo, anche se raramente arrivano a iniettare il veleno. [115] D'altra parte i ragni della famiglia Ctenidae , in modo particolare il genere Phoneutria , reagiscono in modo aggressivo anche a piccole provocazioni. [116] In letteratura vengono riportate approssimativamente cento morti attribuibili a morsi di ragno nell'arco dell'intero ventesimo secolo , [117] a confronto dei circa 1.500 decessi attribuibili ai tentacoli urticanti e pungiglioni di meduse nello stesso periodo. [118] Molti casi attribuiti a morsi di ragno possono anche essere frutto di diagnosi incorrette, [119] cosa che rende più difficile controllare l'effettiva efficacia dei trattamenti contro i morsi veri. [120]
Possibilità di utilizzo per l'uomo

I ragni come le tarantole, cucinati, sono considerati una prelibatezza gastronomica in Cambogia , [121] e dagli indiani Piaroa del Venezuela meridionale, purché i peli estremamente irritanti, sistema difensivo principale di queste specie di ragni, vengano accuratamente rimossi prima di cucinarli. [122]
I veleni secreti dai ragni possono essere un'alternativa concreta meno inquinante agli insetticidi convenzionali; queste tossine, infatti, sono mortali per gli insetti, ma la stragrande maggioranza è assolutamente innocua per l'uomo. Soprattutto gli appartenenti alla famiglia Hexathelidae sono una fonte promettente di produzione di varie tossine in grado di uccidere gli insetti, in quanto non si conoscono specie che sono state in grado di acquisire l' immunità da esse in tutto il mondo; inoltre sono ragni che resistono bene in cattività e abbastanza facili da mungere per estrarne il veleno. Grazie all' ingegneria genetica potrebbe essere possibile codificare i geni che producono queste tossine e inserirli in specifici virus già capaci di infettare le specie parassite delle piante come, ad esempio, quelle del genere Helicoverpa , che parassitano le piante di cotone. [123]
Alcuni ricercatori investigano anche il possibile uso medico di alcune di queste tossine per il trattamento dell' aritmia cardiaca , [124] la malattia di Alzheimer , [125] ictus , ischemie , [126] e la disfunzione erettile . [127]
Poiché la seta del ragno è luminosa e molto resistente, da parte di ingegneri genetici sono stati effettuati tentativi di farla produrre ad altri esseri viventi, in particolare dal latte delle capre e dalle foglie di alcune piante, come il tabacco . [128] [129]
Aracnofilia
In molte persone il solo parlare di ragni provoca ribrezzo e repulsione, per cui ad alcuni risulta difficile concepire che vi siano appassionati che tengono ragni in casa come animali domestici, anche se ben confinati in spazi appositi. In genere si tengono in casa, in appositi terrari , animali di cospicue dimensioni per poterne seguire con attenzione il comportamento e rendersi conto delle necessità che il ragno potrebbe avere. Proprio per il veleno ei peli urticanti di cui dispongono parecchi di loro, vanno maneggiati con la dovuta cautela. [130]
Manutenzione
I ragni di cospicue dimensioni non sono facili da gestire: va approntato un terrario, preferibilmente in vetro, in quanto è più duraturo e stabile della plastica; e, per la pericolosità di alcune specie allevate, deve essere anche a prova di evasione. I vantaggi di quelli in plastica è che sono più leggeri e più facili da pulire, ma col tempo tendono ad ingiallire e le chiusure ad incastro non sono particolarmente affidabili. [131] Bisogna tenere presente che in genere questi ragni abitano in luoghi tropicali, con un certo tasso di umidità e temperatura costante che in qualche modo bisognerà replicare all'interno del terrario, perché riescano a sopravvivere a lungo. Nel terrario andrà inserito un substrato di base: il più adatto a tutte le specie di ragni è la vermiculite , ma anche la perlite va bene ed è più facile da reperire. [132]
Alimentazione
Gli esemplari andranno alimentati principalmente con insetti ( grilli , locuste , mosche , camole ), facilmente reperibili nei negozi specializzati, di taglia proporzionata a quella del ragno e possibilmente vivi, per non far loro perdere le abitudini predatorie che li contraddistinguono. Un po' d'acqua in piccoli contenitori serve a stabilizzare l'umidità ea consentire al ragno di bere se ha sete. Da tenere presente che gli esemplari giovani necessitano di nutrirsi più frequentemente per poter crescere. [133]
Legislazione
La vendita dei ragni nel mondo è regolamentata dalla CITES , in quanto la maggior parte delle specie sono protette. Ad esempio, della numerosa famiglia Theraphosidae , soltanto il genere Euathlus può essere soggetto a commercio ed ogni specie e sottospecie deve riportare una copia del documento CITES. Ogni volta che un esemplare nasce in cattività può essere posto in vendita purché regolarmente certificato. [134]
Più di recente, nel 2006, la CITES ha emesso un comunicato in cui sono ben specificate le specie dei generi Aphonopelma e Brachypelma di cui è fatto divieto assoluto di cattura e commercio esemplari. [135]
Altro
Arachnophilia è anche un programma informatico, un editor di testo in HTML , chiamato con questo nome che deriva dal greco amore per i ragni , in quanto, ed è facile notarne la metafora , consente di costruire reti (informatiche). [136]
Aracnofobia
L' aracnofobia è una specifica fobia , una paura anormale dei ragni e di qualsiasi cosa possa anche solo evocarne il ricordo, come ad esempio, le ragnatele o oggetti che abbiano la forma di ragno. Può essere una forma esagerata di una risposta istintiva atavica che ha aiutato nel passato gli uomini a sopravvivere, [137] o forse un fenomeno culturale che è molto diffuso nelle società prevalentemente europee. [138]
Aracnofobia è anche il titolo di un film del 1990 da cui è derivato un videogioco che porta il titolo originale del film, Arachnophobia , dove una piccola città della California viene invasa da una moltitudine di ragni aggressivi e dal veleno mortale. [139] [140]
I ragni nel simbolismo e nella cultura


I ragni sono il punto focale di molte paure , storie e mitologie di vari popoli sparsi in tutto il mondo e di diverse epoche storiche. [141] Hanno spesso simboleggiato la pazienza, a causa della loro tecnica di caccia che consiste nel costruire la ragnatela e attendere la preda appostandosi, così come hanno anche simboleggiato il dolore e la malevolenza a causa della tossicità del veleno di alcuni di loro. [142] La capacità di costruire le ragnatele ha provocato anche l'associazione del ragno ai miti cosmologici in cui si raffigura qualcosa o qualcuno in grado di costruire da sé il proprio mondo o di apportarvi benefici, con miti molto simili a quello occidentale di Prometeo . [143] La cultura Moche dell'antico Perù adorava la natura; [144] infatti venivano raffigurati molti tipi di animali, in particolare ragni, nelle varie forme della loro arte. [145] Anche l'arte moderna ha spesso raffigurato con vari simbolismi la figura del ragno: ad esempio basti considerare l'opera Maman dell'artista Louise Bourgeois esposta esternamente alla National Gallery of Canada , di Ottawa , dove questo ragno gigantesco alto oltre 9 metri incombe minaccioso, copia in bronzo dell'originale custodito alla Tate Modern di Londra. [146]
Secondo la leggenda, San Norberto avrebbe ingoiato un ragno velenoso caduto nel calice mentre celebrava Messa in una cripta, senza patire danno alcuno. [147]
Note
- ^ a b World Spider Catalogue, versione 20.0, Famiglie di ragni, con numero di generi e specie , su wsc.nmbe.ch . URL consultato il 21 novembre 2019 (archiviato dall' url originale l'11 luglio 2017) .
- ^ Autori vari, Enciclopedia Zanichelli 2002: Dizionario enciclopedico di arti, scienze, tecniche, lettere, filosofia, storia, geografia, diritto, economia , a cura di Edigeo, Zanichelli, 2002, p. 1536, ISBN 978-88-08-09991-4 .
- ^ a b Barnes, RSK, Calow, P., Olive, P., Golding, D., and Spicer, J., Invertebrates with Legs: the Arthropods and Similar Groups , in The Invertebrates: A Synthesis , Blackwell Publishing, 2001, p. 168, ISBN 0-632-04761-5 . URL consultato il 25 settembre 2008 ( archiviato il 20 aprile 2015) .
- ^ a b c d Meehan, C,J. Olson, EJ and Curry, RL, Exploitation of the Pseudomyrmex–Acacia mutualism by a predominantly vegetarian jumping spider ( Bagheera kiplingi ) - 93rd ESA Annual Meeting , 21 agosto 2008. URL consultato il 12 dicembre 2008 (archiviato dall' url originale il 1º dicembre 2019) .
- ^ a b c d Harland, DP, Jackson, RR, "Eight-legged cats" and how they see - a review of recent research on jumping spiders (Araneae: Salticidae) ( PDF ), in Cimbebasia , vol. 16, 2000, pp. 231-240. URL consultato l'11 ottobre 2008 (archiviato dall' url originale il 28 settembre 2006) .
- ^ Mclver, JD, Stonedahl, G., Myrmecomorphy: Morphological and Behavioral Mimicry of Ants , in Annual Review of Entomology , vol. 38, gennaio 1993, pp. 351-377, DOI : 10.1146/annurev.en.38.010193.002031 .
- ^ a b c d e f g h i j k l m n o p q r s t u v w x y z aa ab ac ad ae af ag ah ai aj ak Ruppert, EE, Fox, RS, and Barnes, RD, Invertebrate Zoology , 7ª ed., Brooks / Cole, 2004, pp. 571-584, ISBN 0-03-025982-7 .
- ^ a b Simone Caratozzolo, I ragni giganti: Morfologia, comportamento, alimentazione e riproduzione , De Vecchi Editore, 2000, p. 51, ISBN 88-412-7603-7 .
- ^ Vollrath, F., Eusociality and extraordinary sex ratios in the spider Anelosimus eximius (Araneae: Theridiidae) , in Behavioral Ecology and Sociobiology, , vol. 18, n. 4, 1986, pp. 283-287, ISSN 0340-5443 .
- ^ a b c d e f g Vollrath, F., Selden, PA, The Role of Behavior in the Evolution of Spiders, Silks, and Webs ( PDF ), in Annual Review of Ecology, Evolution, and Systematics , vol. 38, dicembre 2007, pp. 819-846, DOI : 10.1146/annurev.ecolsys.37.091305.110221 . URL consultato il 12 ottobre 2008 ( archiviato il 9 dicembre 2008) .
- ^ a b c Ruppert, EE, Fox, RS, and Barnes, RD, Invertebrate Zoology , 7ª ed., Brooks / Cole, 2004, pp. 554-555, ISBN 0-03-025982-7 .
- ^ a b Ruppert, EE, Fox, RS, and Barnes, RD, Invertebrate Zoology , 7ª ed., Brooks / Cole, 2004, pp. 518-522, ISBN 0-03-025982-7 .
- ^ a b c d e f g Ruppert, EE, Fox, RS, and Barnes, RD, Invertebrate Zoology , 7ª ed., Brooks / Cole, 2004, pp. 559-564, ISBN 0-03-025982-7 .
- ^ EE Ruppert, RS Fox, RD Barnes. Invertebrate Zoology . Ed. 7. Brooks / Cole. ISBN=0030259827. pp. 523-524.
- ^ a b Ruppert, EE, Fox, RS, and Barnes, RD, Invertebrate Zoology , 7ª ed., Brooks / Cole, 2004, pp. 565-569, ISBN 0-03-025982-7 .
- ^ EE Ruppert, RS Fox, RD Barnes. Invertebrate Zoology . 7ª ed. Brooks / Cole, 2004. p. 520. ISBN 0030259827 .
- ^ Stephen Jay Gould , La vita meravigliosa : I fossili di Burgess e la natura della storia , Feltrinelli, 1990, p. 105, ISBN 88-07-81350-5 .
- ^ Ruppert, EE, Fox, RS, and Barnes, RD, Invertebrate Zoology , 7ª ed., Brooks / Cole, 2004, pp. 527-528, ISBN 0-03-025982-7 .
- ^ a b Tracy I. Storer, Robert L. Usinger, Robert C. Stebbins e James W. Nybakken, Zoologia , traduzione di Alfredo Suvero, 10ª ed., Zanichelli , 1992, p. 576, ISBN 978-88-08-00606-6 .
- ^ a b c Coddington, JA & Levi, HW, Systematics and Evolution of Spiders (Araneae) , in Annu. Rev. Ecol. Syst. , vol. 22, 1991, pp. 565-592, DOI : 10.1146/annurev.es.22.110191.003025 .
- ^ autori vari, La Scienza: Il Regno Animale, volume I , collana La Biblioteca di Repubblica, UTET , 2005, p. 336, ISBN977-11-2844522-6 .
- ^ I ragni del Parco del Taro , su parcotaro.it . URL consultato il 7 gennaio 2009 ( archiviato il 4 marzo 2016) .
- ^ a b Tracy I. Storer, Robert L. Usinger, Robert C. Stebbins e James W. Nybakken, Zoologia , traduzione di Alfredo Suvero, 10ª ed., Zanichelli , 1992, p. 575, ISBN 978-88-08-00606-6 .
- ^ a b Ruppert, EE, Fox, RS, and Barnes, RD, Invertebrate Zoology , 7ª ed., Brooks / Cole, 2004, pp. 529-530, ISBN 0-03-025982-7 .
- ^ a b Ruppert, EE, Fox, RS, and Barnes, RD, Invertebrate Zoology , 7ª ed., Brooks / Cole, 2004, pp. 531-532, ISBN 0-03-025982-7 .
- ^ Samuel D. Marshall,Tarantulas and Other Arachnids , 2ª ed., Barron's Educational Series, 2001, pp. 112 , ISBN 978-0-7641-1463-2 .
- ^ Stanley A. Schultz, The Tarantula Keeper's Guide , 2ª ed., Barron's Educational Series, 1998, p. 208, ISBN 978-0-7641-0076-5 .
- ^ a b c d Ruppert, EE, Fox, RS, and Barnes, RD, Invertebrate Zoology , 7ª ed., Brooks / Cole, 2004, pp. 532-537, ISBN 0-03-025982-7 .
- ^ Ruppert, EE, Fox, RS, and Barnes, RD, Invertebrate Zoology , 7ª ed., Brooks / Cole, 2004, pp. 578-580, ISBN 0-03-025982-7 .
- ^ Parry, DA, and Brown, RHJ, The Hydraulic Mechanism of the Spider Leg ( PDF ), in Journal of Experimental Biology , vol. 36, 1959, pp. 423-433. URL consultato il 25 settembre 2008 ( archiviato il 3 ottobre 2008) .
- ^ Anderson, DT, The Chelicerata , in Invertebrate Zoology , 2ª ed., Oxford University Press, 2001, pp. 325-349, ISBN 0-19-551368-1 .
- ^ Vollrath, F., and Knight, DP, Liquid crystalline spinning of spider silk , in Nature , vol. 410, marzo 2001, pp. 541-548, DOI : 10.1038/35069000 . URL consultato il 2 giugno 2016 ( archiviato il 29 giugno 2021) .
- ^ Calamistro , su it.wiktionary.org . URL consultato il 29 aprile 2019 ( archiviato il 3 febbraio 2019) .
- ^ Ruppert, EE, Fox, RS, and Barnes, RD, Invertebrate Zoology , 7ª ed., Brooks / Cole, 2004, pp. 537-539, ISBN 0-03-025982-7 .
- ^ Baerg WJ, 1928 - The life cycle and mating habits of the male Tarantula.
- ^ Gerhardt, U., 1929 - Zur vergleichenden Sexualbiologie primitiver Spinnen, insbesondere der Tetrapneumonen
- ^ Robinson & Robinson, 1973 - The biology of some Argiope species from New Guinea: predatory behaviour and stabilimentum construction (Araneae: Araneidae)
- ^ Bristowe WS, 1941 - The Comity of spiders
- ^ Dondale CD, 1964 - Sexual behavior and its application to a species problem in the spider genus Philodromus (Araneae: Thomisidae)
- ^ Rovner, Jerome S., 1968 - Territoriality in the Sheet-Web Spider Linyphia triangularis (Clerck) (Araneae, Linyphiidae)
- ^ Dondale CD e Hegdekar BM, 1972 - The contact sex pheromone of "Pardosa lapidicina" Emerton (Araneidae: Lycosidae)
- ^ a b Jackson RR, 1977-78 - The mating strategy of "Phidippus johnsoni" (Araneae: Salticidae): 1 - pursuit time and persistence
- ^ Bristowe WS, 1929 - The mating habits of spiders, with special reference to the problems surrounding sex dimorphism
- ^ Crane, J. 1949. Comparative biology of salticid spiders at Rancho Grande, Venezuela. Part IV. An analysis of display
- ^ Kaston BJ, 1936 - The senses involved in the courtship of some vagabond spiders
- ^ Knoflach, B. & van Harten, A., Tidarren argo sp. nov (Araneae: Theridiidae) and its exceptional copulatory behaviour: emasculation, male palpal organ as a mating plug and sexual cannibalism , in Journal of Zoology , vol. 254, 2001, pp. 449-459.
- ^ Andrade, Maydianne CB, Risky mate search and male self-sacrifice in redback spiders , in Behavioral Ecology , vol. 14, 2003, pp. 531-538.
- ^ a b c d Foelix, RF, Reproduction , in Biology of Spiders , Oxford University Press US, 1996, pp. 176-212, ISBN 0-19-509594-4 . URL consultato l'8 ottobre 2008 ( archiviato il 20 aprile 2015) .
- ^ "Strategie riproduttive dei ragni", di Ken e Rod Preston, pubbl. su "Le Scienze", num.365, pag.88-93
- ^ a b autori vari, La Scienza: Il Regno Animale, volume I , collana La Biblioteca di Repubblica, UTET , 2005, p. 344, ISSN 1128-4455 .
- ^ Ruppert, EE, Fox, RS, and Barnes, RD, Invertebrate Zoology , 7ª ed., Brooks / Cole, 2004, pp. 523 -524, ISBN 0-03-025982-7 .
- ^ Simone Caratozzolo, I ragni giganti: Morfologia, comportamento, alimentazione e riproduzione , De Vecchi Editore, 2000, p. 49, ISBN 88-412-7603-7 .
- ^ Simone Caratozzolo, I ragni giganti: Morfologia, comportamento, alimentazione e riproduzione , De Vecchi Editore, 2000, pp. 49-50-51, ISBN 88-412-7603-7 .
- ^ Spiders and their Kin , Herbert W. Levi and Lorna R. Levi, Golden Press, p. 20 and p. 44
- ^ The World Spider Catalog di Platnick, famiglia Theraphosidae , su research.amnh.org . URL consultato il 31 agosto 2014 ( archiviato il 3 settembre 2014) .
- ^ Simone Caratozzolo, I ragni giganti: Morfologia, comportamento, alimentazione e riproduzione , De Vecchi Editore, 2000, p. 91, ISBN 88-412-7603-7 .
- ^ Simone Caratozzolo, I ragni giganti: Morfologia, comportamento, alimentazione e riproduzione , De Vecchi Editore, 2000, p. 6, ISBN 88-412-7603-7 .
- ^ Hünicken, MA (1980). A giant fossil spider ( Megarachne servinei ) from Bajo de Véliz, Upper Carboniferous, Argentina. Boletin de la Academia Nacional de Ciencias, Córdoba, Argentina 53 : 317-341.
- ^ Selden, PA, Corronca, JA & Hünicken, MA (2005). The true identity of the supposed giant fossil spider Megarachne . Biology Letters 1 : 44-48.
- ^ a b c d e Oxford, GS & Gillespie, RG (1998). Evolution and Ecology of Spider Coloration. Annual Review of Entomology 43:619-643.
- ^ a b Jackson, RR et al. , Jumping spiders (Araneae: Salticidae) that feed on nectar ( PDF ), in Journal of Zoology , vol. 255, 2001, pp. 25-29, DOI : 10.1017/S095283690100108X . URL consultato il 27 dicembre 2008 ( archiviato il 18 marzo 2009) .
- ^ Beckman, N. and Hurd, LE, [0881:PFAFIP 2.0.CO;2 Pollen Feeding and Fitness in Praying Mantids: The Vegetarian Side of a Tritrophic Predator ], in Environmental Entomology , vol. 32, n. 4, agosto 2003, pp. 881-885, DOI : 10.1603/0046-225X(2003)032[0881:PFAFIP]2.0.CO;2 .
- ^ Schütz, D., and Taborsky, M., Adaptations to an aquatic life may be responsible for the reversed sexual size dimorphism in the water spider, Argyroneta aquatica. ( PDF ), in Evolutionary Ecology Research , vol. 5, n. 1, 2003, pp. 105-117. URL consultato l'11 ottobre 2008 (archiviato dall' url originale il 16 dicembre 2008) .
- ^ Coddington, J., and Sobrevila, C., Web manipulation and two stereotyped attack behaviors in the ogre-faced spider Deinopis spinosus Marx (Araneae, Deinopidae). ( PDF ), in Journal of Arachnology , vol. 15, 1987, pp. 213-225. URL consultato l'11 ottobre 2008 ( archiviato il 9 dicembre 2008) .
- ^ Eberhard, WG, Aggressive Chemical Mimicry by a Bolas Spider ( PDF ), in Science , vol. 198, n. 4322, dicembre 1977, pp. 1173-1175, DOI : 10.1126/science.198.4322.1173 . URL consultato il 10 ottobre 2008 (archiviato dall' url originale il 9 dicembre 2008) .
- ^ Eberhard, WG, The Natural History and Behavior of the Bolas Spider, Mastophora dizzydeani sp. n. (Araneae) , in Psyche , vol. 87, 1980, pp. 143-170. URL consultato il 10 ottobre 2008 ( archiviato il 20 dicembre 2008) .
- ^ Yeargan, KV, and Quate, LW, Adult male bolas spiders retain juvenile hunting tactics , in Oecologia , vol. 112, n. 4, 1997, pp. 572-576, DOI : 10.1007/s004420050347 .
- ^ Scoperti ragni che si nutrono di pesce , su ansa.it , ANSA , 19 giugno 2014. URL consultato il 19 giugno 2014 ( archiviato il 19 giugno 2014) .
- ^ a b Wilcox, S. and Jackson, R., Jumping Spider Tricksters , in Bekoff, M., Allen, C., and Burghardt, GM (a cura di), The Cognitive Animal: Empirical and Theoretical Perspectives on Animal Cognition , MIT Press, 2002, pp. 27 -34, ISBN 0-262-52322-1 .
- ^ Mclver, JD and Stonedahl, G., Myrmecomorphy: Morphological and Behavioral Mimicry of Ants , in Annual Review of Entomology , vol. 38, gennaio 1993, pp. 351-377, DOI : 10.1146/annurev.en.38.010193.002031 .
- ^ Different smiles, single species , su evolution.berkeley.edu , University of California Museum of Paleontology. URL consultato il 10 ottobre 2008 ( archiviato il 20 dicembre 2008) .
- ^ Cooke, JAL, Roth, VD, and Miller, FH, The urticating hairs of theraphosid spiders , in American Museum novitates , n. 2498, American Museum of Natural History. URL consultato l'11 ottobre 2008 ( archiviato il 21 dicembre 2014) .
- ^ Blackledge, TA, and Wenzel, JW, Silk Mediated Defense by an Orb Web Spider against Predatory Mud-dauber Wasps , in Behaviour , vol. 138, n. 2, 2001, pp. 155-171, DOI : 10.1163/15685390151074357 .
- ^ Armstrong, S., Fog, wind and heat - life in the Namib desert , in New Scientist , 14 luglio 1990. URL consultato l'11 ottobre 2008 ( archiviato il 24 settembre 2008) .
- ^ ( EN ) Jason Daley, The World's Oldest Known Spider Has Died at Age 43 , su Smithsonian . URL consultato il 7 maggio 2018 ( archiviato il 7 maggio 2018) .
- ^ Simone Caratozzolo, I ragni giganti: Morfologia, comportamento, alimentazione e riproduzione , De Vecchi Editore, 2000, p. 96, ISBN 88-412-7603-7 .
- ^ Vollrath, F., Eusociality and extraordinary sex ratios in the spider Anelosimus eximius (Araneae: Theridiidae) , in Behavioral Ecology and Sociobiology , vol. 18, n. 4, 1986, pp. 283-287, ISSN 0340-5443 .
- ^ Agnarsson, I., and Kuntner, M., Madagascar: an unexpected hotspot of social Anelosimus spider diversity (Araneae: Theridiidae) , in Systematic Entomology , vol. 30, n. 4, aprile 2005, pp. 575-592, DOI : 10.1111/j.1365-3113.2005.00289.x .
- ^ Avilés, L., Maddison, WP and Agnarsson, I., A New Independently Derived Social Spider with Explosive Colony Proliferation and a Female Size Dimorphism , in Biotropica , vol. 38, n. 6, agosto 2006, pp. 743-753, DOI : 10.1111/j.1744-7429.2006.00202.x .
- ^ Matsumoto, T., Cooperative prey capture in the communal web spider, Philoponella raffray (Araneae, Uloboridae) ( PDF ), in Journal of Arachnology , vol. 26, 1998, pp. 392-396. URL consultato l'11 ottobre 2008 ( archiviato il 28 agosto 2008) .
- ^ Parassitismo#Cleptoparassita
- ^ Cangialosi, KR, Social spider defense against kleptoparasitism , in Behavioral Ecology and Sociobiology , vol. 27, n. 1, luglio 1990, DOI : 10.1007/BF00183313 .
- ^ Bertani, R., Fukushima, CS, and Martins, R., Sociable widow spiders? Evidence of subsociality in Latrodectus Walckenaer, 1805 (Araneae, Theridiidae) , in Journal of Ethology , vol. 26, n. 2, maggio 2008, DOI : 10.1007/s10164-007-0082-8 .
- ^ a b c d e Eberhard, WG, Function and Phylogeny of Spider Webs ( PDF ), in Annual Reviews in Ecology and Systematics , vol. 21, 1990, pp. 341-372, DOI : 10.1146/annurev.es.21.110190.002013 . URL consultato il 15 ottobre 2008 (archiviato dall' url originale il 9 dicembre 2008) .
- ^ a b Agnarsson, I., Morphological phylogeny of cobweb spiders and their relatives (Araneae, Araneoidea, Theridiidae) , in Zoological Journal of the Linnean Society , vol. 141, n. 4, 2004, pp. 447-626, DOI : 10.1111/j.1096-3642.2004.00120.x .
- ^ Herberstein, ME, Foraging behaviour in orb-web spiders (Araneidae): do web decorations increase prey capture success in Argiope keyserlingi Karsch, 1878? , in Australian Journal of Zoology , vol. 48, 2, pp. 217-223, DOI : 10.1071/ZO00007 .
- ^ Li, D., and Lee, WS, Predator-induced plasticity in web-building behaviour , in Animal Behaviour , vol. 67, n. 2, febbraio 2004, pp. 309-318, DOI : 10.1016/j.anbehav.2003.06.011 .
- ^ Schütt, K., Drapetisca socialis (Araneae: Linyphiidae): Web reduction - ethological and morphological adaptations ( PDF ), in European Journal of Entomology , 9 2, 1995, pp. 553-563. URL consultato il 19 ottobre 2008 ( archiviato il 16 dicembre 2008) .
- ^ James L. Gould e Carol Grant Gould, L'architettura degli animali: Nidi, tane, alveari , traduzione di Cristina Serra, collana Scienza e Idee, Raffaello Cortina Editore , 2007, pp. 57-58, ISBN 978-88-6030-163-5 .
- ^ T homson, Peggy and Park, Edwards, Odd Tales from the Smithsonian , su smithsonianeducation.org . URL consultato il 21 luglio 2008 ( archiviato il 22 aprile 2021) .
- ^ Hecht, H., Oldest spider web found in amber , su newscientist.com , New Scientist. URL consultato l'11 gennaio 2009 ( archiviato il 22 dicembre 2008) .
- ^ Hecht, H., Oldest spider web found in amber , su newscientist.com , New Scientist. URL consultato il 15 ottobre 2008 ( archiviato il 28 agosto 2008) .
- ^ Penney, D., and Selden, PA, Spinning with the dinosaurs: the fossil record of spiders , in Geology Today , vol. 23, n. 6, 2007, pp. 231-237, DOI : 10.1111/j.1365-2451.2007.00641.x .
- ^ Dunlop, JA, A trigonotarbid arachnid from the Upper Silurian of Shropshire ( PDF ), in Palaeontology , vol. 39, n. 3, settembre 1996, pp. 605-614. URL consultato il 12 ottobre 2008 ( archiviato il 16 dicembre 2008) .
- ^ Dunlop, JA, A replacement name for the trigonotarbid arachnid Eotarbus Dunlop , in Palaeontology , vol. 42, n. 1, p. 191.
- ^ Selden, PA, and Shear, WA, Fossil evidence for the origin of spider spinnerets ( PDF ), in Nature Precedings , luglio 2008. URL consultato il 12 ottobre 2008 ( archiviato il 10 agosto 2016) .
- ^ Selden, PA, Fossil mesothele spiders , in Nature , vol. 379, febbraio 1996, pp. 498-499, DOI : 10.1038/379498b0 .
- ^ a b c d e Schultz, JW, A phylogenetic analysis of the arachnid orders based on morphological characters , in Zoological Journal of the Linnean Society 2007; 150: 221–265 , DOI : 10.1111/j.1096-3642.2007.00284.x .
- ^ Stephen Jay Gould , La vita meravigliosa : I fossili di Burgess e la natura della storia , Feltrinelli , 1990, p. 105, ISBN 88-07-81350-5 .
- ^ a b c d e f g Coddington, JA, Phylogeny and Classification of Spiders ( PDF ), in Ubick, D., Paquin, P., Cushing, PE, and Roth, V. (a cura di), Spiders of North America: an identification manual , American Arachnological Society, 2005, pp. 18-24, ISBN 0-9771439-0-2 . URL consultato il 12 ottobre 2008 ( archiviato il 9 dicembre 2008) .
- ^ a b Leroy, J, and Leroy, A., How spiders function , in Spiders of Southern Africa , Struik, 2003, pp. 15-21, ISBN 1-86872-944-3 . URL consultato il 27 dicembre 2008 (archiviato dall' url originale il 20 aprile 2015) .
- ^ Ono, H., New and Remarkable Spiders of the Families Liphistiidae, Argyronetidae, Pisauridae, Theridiidae and Araneidae (Arachnida) from Japan , in Bulletin of the National Science Museum (of Japan), Series A , vol. 28, n. 1, 2002, pp. 51-60.
- ^ Coyle, FA, The Role of Silk in Prey Capture , in Shear, WA (a cura di), Spiders – Webs, Behavior, and Evolution , Stanford University Press, 1986, pp. 272-273, ISBN 0-8047-1203-4 .
- ^ Forster, RR, and Platnick, NI, A review of the archaeid spiders and their relatives, with notes on the limits of the superfamily Palpimanoidea (Arachnida, Araneae) ( abstract ), in Bulletin of the American Museum of Natural History , vol. 178, 1984, pp. 1-106. URL consultato il 13 ottobre 2008 ( archiviato il 29 giugno 2021) . Testo completo su A review of the archaeid spiders and their relatives ( PDF ), su digitallibrary.amnh.org . URL consultato il 13 ottobre 2008 ( archiviato il 16 dicembre 2008) . (60 MB)
- ^ Penney, D., and Selden, PA, Assembling the Tree of Life—Phylogeny of Spiders: a review of the strictly fossil spider families ( PDF ), in Acta Zoologica Bulgarica , European Arachnology 2005 - supplemento N. 1, Deltshev, C., and Stoev, P., pp. 25-39. URL consultato il 13 ottobre 2008 ( archiviato il 16 dicembre 2008) .
- ^ Sito tassonomico olandese Archiviato l'8 marzo 2016 in Internet Archive .; Sito tassonomico internazionale Archiviato il 30 dicembre 2008 in Internet Archive .; Sito tassonomico Archiviato il 3 febbraio 2019 in Internet Archive .
- ^ Natural history of Mygalomorphae , su arc.agric.za , Agricultural Research Council of New Zealand. URL consultato il 13 ottobre 2008 (archiviato dall' url originale il 26 dicembre 2008) .
- ^ autori vari, La Scienza: Il Regno Animale, volume I , collana La Biblioteca di Repubblica, UTET , 2005, p. 800, ISBN977-11-2844522-6 .
- ^ a b c Trotta, Alessio, 2005 - Introduzione ai ragni italiani (Arachnida Araneae) , Memorie della Società Entomologica italiana, vol. 83 , p.3-178 Articolo in PDF Archiviato il 24 settembre 2015 in Internet Archive .
- ^ a b c d e f Checklist dei ragni italiani del Museo di Scienze Naturali di Bergamo a cura di Paolo Pantini e Marco Isaia del 2014 , su museoscienzebergamo.it . URL consultato il 9 marzo 2015 (archiviato dall' url originale il 19 aprile 2015) .
- ^ Checklist of italian fauna, aggiornato al 2003 , su faunaitalia.it . URL consultato il 19 marzo 2013 ( archiviato il 27 dicembre 2012) .
- ^ Spiders , su idph.state.il.us , Illinois Department of Public Health. URL consultato l'11 ottobre 2008 ( archiviato il 15 febbraio 2012) .
- ^ Vetter R, Barger D, An infestation of 2,055 brown recluse spiders (Araneae: Sicariidae) and no envenomations in a Kansas home: implications for bite diagnoses in nonendemic areas , in J Med Entomol , vol. 39, n. 6, 2002, pp. 948-51, PMID.
- ^ Hannum, C.and Miller, DM, Widow Spiders , su ext.vt.edu , Department of Entomology, Virginia Tech. URL consultato l'11 ottobre 2008 (archiviato dall' url originale il 18 ottobre 2008) .
- ^ Funnel web spiders , su avru.org , Australian Venom Research Unit. URL consultato l'11 ottobre 2008 ( archiviato il 9 ottobre 2008) .
- ^ Pub chef bitten by deadly spider , su news.bbc.co.uk , BBC. URL consultato l'11 ottobre 2008 ( archiviato il 14 febbraio 2009) .
- ^ Diaz, JH, The Global Epidemiology, Syndromic Classification, Management, and Prevention of Spider Bites , in American Journal of Tropical Medicine and Hygiene , vol. 71, n. 2, 2004, pp. 239-250. URL consultato l'11 ottobre 2008 ( archiviato l'11 maggio 2011) .
- ^ Williamson, JA, Fenner, PJ, Burnett, JW, and Rifkin, J., Venomous and Poisonous Marine Animals: A Medical and Biological Handbook , UNSW Press, 1996, pp. 65-68, ISBN 0-86840-279-6 . URL consultato il 3 ottobre 2008 ( archiviato il 20 aprile 2015) .
- ^ Nishioka, S de A., Misdiagnosis of brown recluse spider bite , in Western Journal of Medicine , vol. 175, n. 2, agosto 2001, p. 240. URL consultato il 3 ottobre 2008 ( archiviato il 29 giugno 2021) .
- ^ Isbister, GK, Spider mythology across the world , in Western Journal of Medicine , vol. 174, n. 4, aprile 2001, pp. 86-87. URL consultato il 3 ottobre 2008 ( archiviato il 29 giugno 2021) .
- ^ Ray, N., Lonely Planet Cambodia , Lonely Planet Publications, 2002, p. 308, ISBN 1-74059-111-9 .
- ^ Weil, C., Fierce Food , Plume, 2006, ISBN 0-452-28700-6 . URL consultato il 3 ottobre 2008 (archiviato dall' url originale l'11 maggio 2011) .
- ^ Spider Venom Could Yield Eco-Friendly Insecticides , su nsf.gov , National Science Foundation (USA). URL consultato l'11 ottobre 2008 ( archiviato il 20 dicembre 2008) .
- ^ Novak, K., Spider venom helps hearts keep their rhythm , in Nature Medicine , vol. 7, n. 155, 2001, DOI : 10.1038/84588 .
- ^ Lewis, RJ, and Garcia, ML, Therapeutic potential of venom peptides ( PDF ), in Nature Reviews Drug Discovery , vol. 2, ottobre 2003, pp. 790-802, DOI : 10.1038/nrd1197 . URL consultato l'11 ottobre 2008 (archiviato dall' url originale il 16 dicembre 2008) .
- ^ Bogin, O., Venom Peptides and their Mimetics as Potential Drugs ( PDF ), in Modulator , n. 19, Spring 2005. URL consultato l'11 ottobre 2008 (archiviato dall' url originale il 9 dicembre 2008) .
- ^ Andrade, E., Villanova, F., Borra, P., et al, Penile erection induced in vivo by a purified toxin from the Brazilian spider Phoneutria nigriventer . , in British Journal of Urology International , vol. 102, n. 7, pp. 835-837, DOI : 10.1111/j.1464-410X.2008.07762.x .
- ^ Hinman, MB, Jones JA, and Lewis, RW, Synthetic spider silk: a modular fiber ( PDF ), in Trends in Biotechnology , vol. 18, n. 9, settembre 2000, pp. 374-379, DOI : 10.1016/S0167-7799(00)01481-5 . URL consultato il 19 ottobre 2008 (archiviato dall' url originale il 16 dicembre 2008) .
- ^ Menassa, R., Zhu, H., Karatzas, CN, Lazaris, A., Richman, A., and Brandle, J., Spider dragline silk proteins in transgenic tobacco leaves: accumulation and field production , in Plant Biotechnology Journal , vol. 2, n. 5, giugno 2004, pp. 431-438, DOI : 10.1111/j.1467-7652.2004.00087.x .
- ^ Simone Caratozzolo, I ragni giganti: Morfologia, comportamento, alimentazione e riproduzione , De Vecchi Editore, 2000, p. 23, ISBN 88-412-7603-7 .
- ^ Simone Caratozzolo, I ragni giganti: Morfologia, comportamento, alimentazione e riproduzione , De Vecchi Editore, 2000, pp. 34-35, ISBN 88-412-7603-7 .
- ^ Simone Caratozzolo, I ragni giganti: Morfologia, comportamento, alimentazione e riproduzione , De Vecchi Editore, 2000, pp. 37-38, ISBN 88-412-7603-7 .
- ^ Simone Caratozzolo, I ragni giganti: Morfologia, comportamento, alimentazione e riproduzione , De Vecchi Editore, 2000, pp. 41-42-43, ISBN 88-412-7603-7 .
- ^ Simone Caratozzolo, I ragni giganti: Morfologia, comportamento, alimentazione e riproduzione , De Vecchi Editore, 2000, p. 24, ISBN 88-412-7603-7 .
- ^ http://forum.aracnofilia.org/index.php/topic/12083-guida-la-normativa-cites/ Comunicato della CITES Archiviato il 30 ottobre 2011 in Internet Archive .
- ^ Home page di Arachnophilia , su arachnoid.com . URL consultato il 31 dicembre 2008 ( archiviato il 20 dicembre 2008) .
- ^ Friedenberg, J., and Silverman, G., Cognitive Science: An Introduction to the Study of Mind , SAGE, 2005, pp. 244-245, ISBN 1-4129-2568-1 . URL consultato l'11 ottobre 2008 ( archiviato il 20 aprile 2015) .
- ^ Davey, GCL, The “Disgusting” Spider: The Role of Disease and Illness in the Perpetuation of Fear of Spiders , in Society and Animals , vol. 2, n. 1, 1994, pp. 17-25, DOI : 10.1163/156853094X00045 .
- ^ Maltin, L., and Edelman, R. (a cura di), Leonard Maltin's Movie and Video Guide 2002 , Signet, 2002, p. 58, ISBN 978-0-451-20392-2 .
- ^ IGN: Arachnophobia , su uk.pc.ign.com . URL consultato l'11 ottobre 2008 ( archiviato il 7 dicembre 2008) .
- ^ Gail De Vos, Tales, Rumors, and Gossip: Exploring Contemporary Folk Literature in Grades 7-12 , Libraries Unlimited, 1996, p. 186, ISBN 1-56308-190-3 . URL consultato il 22 aprile 2008 ( archiviato il 22 luglio 2011) .
- ^ Jana Garai, The Book of Symbols , New York, Simon & Schuster, 1973, ISBN 0-671-21773-9 .
- ^ Frederica De Laguna et al, American Anthropology: Papers from the American Anthropologist , University of Nebraska Press, 2002, p. 455, ISBN 0-8032-8280-X . URL consultato il 21 aprile 2008 .
- ^ Benson, Elizabeth, The Mochica: A Culture of Peru. New York, NY: Praeger Press. 1972
- ^ Berrin, Katherine & Larco Museum . The Spirit of Ancient Peru:Treasures from the Museo Arqueológico Rafael Larco Herrera . New York: Thames and Hudson, 1997.
- ^ Louise Bourgeois
- ^ ( FR ) Christophe Renault, Reconnaître les saints et les personnages de la Bible , Paris, Éditions Jean-Paul Gisserot, 2002 ISBN 978-287747-658-4 . p. 19
Bibliografia
- AA.VV., La Scienza: Il Regno Animale, volume I , collana La Biblioteca di Repubblica, UTET, 2005, p. 800.
- ( EN ) Bristowe, WS, The World of Spiders , Taplinger Pub Co, 1976, ISBN 0-8008-8598-8 .
- Caratozzolo, S., I ragni giganti: Morfologia, comportamento, alimentazione e riproduzione , De Vecchi Editore, 2000, p. 96, ISBN 88-412-7603-7 .
- ( EN ) Crompton, J., 1950 - The Life of the Spider , Mentor.
- Gould, JL e Gould, CG, L'architettura degli animali: Nidi, tane, alveari , traduzione di Cristina Serra, collana Scienza e Idee, Raffaello Cortina Editore, 2007, p. 380, ISBN 978-88-6030-163-5 .
- ( EN ) Hillyard, P., 1994 - The Book of the Spider , Random House, New York.
- ( EN ) Kaston, BJ, 1953 - How to Know the Spiders , Dubuque.
- ( EN ) Main, BY, 1976 - Spiders , Collins (The Australian Naturalist Library), Sydney.
- Storer, TI, Usinger, RL, Stebbins, RC e Nybakken, JW, Zoologia , traduzione di Alfredo Suvero, 10ª ed., Zanichelli, 1992, p. 1116, ISBN 978-88-08-00606-6 .
- ( EN ) Ubick, D., Paquin P., Cushing, PE, e Roth, V., 2005 - Spiders of North America: an Identification Manual , American Arachnological Society.
- ( EN ) Wise, DH, 1993 - Spiders in Ecological Webs . Cambridge University Press. Great Britain.
- ( DE ) Gerhardt, U., 1929 - Zur vergleichenden Sexualbiologie primitiver Spinnen, insbesondere der Tetrapneumonen . Zeitschrift fur Morphologie und Okologie der Tiere vol. 14 , p. 699–764.
- ( EN ) Baerg WJ, 1928 - The life cycle and mating habits of the male Tarantula. , Quart. Rev. Biol. Baltimore vol.3, nº1, p. 109-116.
- ( EN ) Crane, J. 1949. Comparative biology of salticid spiders at Rancho Grande, Venezuela. Part IV. An analysis of display . Zoologica, vol. 34 , p. 159-214.
- ( EN ) Bristowe WS, 1941 - The Comity of spiders , Ray Society, vol.2, pp. i-xiv + 229-260.
- ( EN ) Dondale CD, 1964 - Sexual behavior and its application to a species problem in the spider genus Philodromus (Araneae: Thomisidae) , Canadian Journal of Zoology. Vol.42, p. 817-826.
- ( DE ) Rovner, Jerome S., 1968 - Territoriality in the Sheet-Web Spider Linyphia triangularis (Clerck) (Araneae, Linyphiidae) , Zeitschrift für Tierpsychologie, vol. 25 (2), p. 232-242. Abstract, in tedesco
- ( EN ) Dondale CD e Hegdekar BM, 1972 - The contact sex pheromone of "Pardosa lapidicina" Emerton (Araneidae: Lycosidae) , Can. J. Zool. vol.51, p. 400-401.
- ( EN ) Robinson, Barbara C. & Robinson, Michael H., 1973 - The biology of some Argiope species from New Guinea: predatory behaviour and stabilimentum construction (Araneae: Araneidae) , Zoological Journal of the Linnean Society, vol. 54 (2), p. 145-159 Abstract
- ( EN ) Kaston BJ, 1936 - The senses involved in the courtship of some vagabond spiders , Ent. Amer. Brooklyn (WS), vol.16, p. 97-167.
- ( EN ) Jackson RR, 1977-78 - The mating strategy of "Phidippus johnsoni" (Araneae: Salticidae): 1 - pursuit time and persistence . Behavioural Ecol. Sociobiol. vol.4 (2), p. 123-132.
- ( EN ) Bristowe WS, 1929 - The mating habits of spiders, with special reference to the problems surrounding sex dimorphism . Proceedings of the Zoological Society of London, p. 309-358.
Altri progetti
-
 Wikiquote contiene citazioni sul ragno
Wikiquote contiene citazioni sul ragno -
 Wikizionario contiene il lemma di dizionario « ragno »
Wikizionario contiene il lemma di dizionario « ragno » -
 Wikimedia Commons contiene immagini o altri file sul ragno
Wikimedia Commons contiene immagini o altri file sul ragno -
 Wikispecies contiene informazioni sul ragno
Wikispecies contiene informazioni sul ragno
Collegamenti esterni
- Araneae , su sapere.it , De Agostini .
- ( EN ) Araneae , su Enciclopedia Britannica , Encyclopædia Britannica, Inc.
- ( EN , FR ) Araneae , su Enciclopedia canadese .
- ( EN ) Araneae , su Fossilworks.org .
Siti di interesse generale
- Arachnology Home Page: Araneae — Ragni , su arachnology.be . URL consultato il 27 dicembre 2008 (archiviato dall' url originale il 1º aprile 2007) .
- Deutsche Arachnologische Gesellschaft e. V. — Società aracnologica tedesca, sito in inglese/tedesco , su dearge.de .
- Sito di Ed Nieuwenhuys sui ragni , su xs4all.nl .
- Spider Pest Control Information — National Pesticide Information Center , su npic.orst.edu .
- Sito dell'International Society of Arachnology, ISA , su arachnology.org .
Siti regionali
- Ragni australiani , su amonline.net.au .
- Ragni portoghesi , su aranhas.info . URL consultato il 28 maggio 2020 (archiviato dall' url originale il 26 febbraio 2019) .
- Ragni della Gran Bretagna , su britishspiders.org.uk .
- Ragni del Caucaso , su caucasus-spiders.info .
- Ragni del Wisconsin , su spiders.entomology.wisc.edu . URL consultato il 27 dicembre 2008 (archiviato dall' url originale il 29 gennaio 2009) .
- Guida per la ricerca dei ragni nel Queensland , su usq.edu.au . URL consultato il 27 dicembre 2008 (archiviato dall' url originale il 10 marzo 2007) .
- Watson, L., and Dallwitz, MJ 2004 onwards. The families of spiders represented in the British Isles. , su delta-intkey.com .
Siti sulla morfologia
- Morfologia, biologia e tassonomia degli Araneae , su parcotaro.it .
Siti sulla tassonomia
- Database dei ragni e degli aracnologi del Canada e della regione neartica , su canadianarachnology.org .
- The world spider catalog - versione 15, del 1º semestre 2014.
- Numero totale dei generi e delle specie descritte al mondo , su research.amnh.org .
- Sito in tedesco e in inglese per la determinazione dei caratteri tassonomici , su araneae.unibe.ch . URL consultato il 18 gennaio 2009 (archiviato dall' url originale il 23 febbraio 2009) .
- Sito tassonomico olandese , su taxonomicon.taxonomy.nl .
- Sito tassonomico internazionale , su zipcodezoo.com .
- Sito tassonomico dell'ITIS , su itis.gov .
Altro
- Naturamediterraneo, forum sugli Artropodi , su naturamediterraneo.com .
- Sito sui Linyphiidae di Tanasevitch , su andtan.newmail.ru . URL consultato il 18 gennaio 2009 (archiviato dall' url originale il 4 febbraio 2012) .
- Sezione del sito di Science Daily aggiornato sui ragni , su sciencedaily.com .
- Sito bibliografico , su www2.udec.cl . URL consultato il 18 gennaio 2009 (archiviato dall' url originale il 23 dicembre 2008) .
- Università della California, valutazioni della gravità del morso di un ragno , su spiders.ucr.edu . URL consultato il 5 febbraio 2001 (archiviato dall' url originale il 5 febbraio 2001) .
- Ragnatele costruite da ragni alimentati con mosche a loro volta nutrite con farmaci e droghe , su cannabis.net .


| Controllo di autorità | Thesaurus BNCF 2829 · LCCN ( EN ) sh85126625 · BNF ( FR ) cb119440506 (data) |
|---|