Electrocomunicații

Electrocomunicația este metoda de comunicare utilizată de peștii slab electrici . Peștii slabi electrici sunt un grup de animale care utilizează un canal de comunicare „invizibil” pentru majoritatea celorlalte animale: semnalizarea electrică. Peștii electrici comunică electric prin intermediul unui pește care generează un câmp electric și un al doilea individ care primește câmpul electric cu electroreceptorii săi. Partea destinatară va interpreta frecvențele semnalului, formele de undă și întârzierea etc. [1] Cele mai bine studiate specii sunt două linii de apă dulce: Mormyridae africane și Gymnotiformes din America de Sud . [2] În timp ce peștii slab electrici sunt singurul grup identificat care generează și primesc câmpuri electrice, alte specii fie generează semnale, fie le primesc, dar nu ambele. Animalele care generează sau primesc câmpuri electrice se găsesc numai în medii acvatice (sau cel puțin umede) datorită rezistenței mari a tuturor celorlalte medii (de exemplu aerul). [3] Până în prezent, s-a stabilit că comunicarea dintre peștii electrici servește în principal scopului de transmitere a informațiilor în legătură cu:
- recunoașterea speciilor
- curtarea și recunoașterea sexuală
- stare motivațională (avertizare de atac sau supunere) e
- conditii de mediu.
Prezentare generală a peștilor slab electrici

Peștii electrici sunt capabili să genereze câmpuri electrice externe sau să primească câmpuri electrice ( electrorecepție ). Peștii electrici pot fi împărțiți în trei categorii: descărcare puternică, descărcare slabă și pești care simt, dar nu pot genera câmpuri electrice. [1] Peștii puternic electrici generează un câmp electric puternic de până la 500 de volți în scopuri de pradă. [4] Peștii puternic electrici includ atât peștii marini, cât și peștii de apă dulce (doi pești de apă dulce - somnul electric african ( Malapterurus electricus ) și anghila electrică neotropicală ( Electrophorus electricus ) și torpilele marine ( Torpedo )). Peștii slab electrici generează câmpuri electrice în principal în scopuri de comunicare și electrolocalizare; Peștii slabi electrici se găsesc numai în apă dulce și includ Mormyridae și Gymnarchus din apă dulce africană și pește-spadă electric neotropical. În cele din urmă, peștii care sunt capabili doar să detecteze semnale electrice includ rechini, raze, somn și alte câteva grupuri (vezi Electrorecepția ). [4]
Peștii electrici generează descărcări de la organele electrice situate în apropierea regiunii cozii. Organele electrice sunt în mare parte derivate din celule musculare (miogene); o excepție este o familie de gimnotiforme care are organul electric derivat din neuroni (organe neurogene). Pentru a detecta semnalele electrice, peștele electric are două tipuri de celule receptive - electroceptori ampulari și tuberoși.
Organe electro-receptive
Toate organismele răspund la șocuri electrice suficient de puternice, dar numai unele vertebrate acvatice pot detecta și utiliza câmpuri electrice slabe, cum ar fi cele care apar în mod natural. Aceste organisme acvatice sunt, prin urmare, numite electroreceptive. (De exemplu, oamenii reacționează la curenți electrici puternici cu un sentiment de durere și uneori cu un amestec de alte simțuri; totuși, sunt incapabili să detecteze câmpuri electrice slabe și, prin urmare, nu sunt electroreceptivi.) Capacitatea de a percepe și utiliza câmpurile electrice este găsit aproape exclusiv la vertebratele acvatice inferioare (pești și unii amfibieni). Animalele terestre, cu foarte puține excepții, nu au acest canal de detecție electrică din cauza conductivității scăzute a aerului, a solului sau a mediilor, altele decât mediul apos. Excepții includ monotremele australiene, adică echidna , care mănâncă în principal furnici și termite, și ornitorincul semiaquatic , care vânează folosind câmpurile electrice generate de prada nevertebratelor. [5]
Pentru a detecta câmpuri electrice slabe, animalele trebuie să aibă electroreceptori (organe receptive) care detectează diferențele de potențial electric. Pentru peștii electrici, organele receptive sunt grupuri de celule senzoriale înrădăcinate în gropi epidermice, care arată ca niște pete minuscule pe piele. În fiecare organ receptiv, există celule senzoriale încorporate în partea de jos a „puțului” deschis care se îndreaptă spre exterior. Electroreceptorii detectează semnale electrice prin crearea unei diferențe de potențial între mediul extern și mediul intern al corpului peștilor. Fluxul de curent datorat acestei diferențe de potențial are ca rezultat un potențial receptiv care este presinaptic al fibrelor senzoriale. În cele din urmă, acest potențial receptiv duce la potențialul de acțiune lansat de celulele senzoriale. [6]
Peștii electrici poartă o varietate de organe receptive sensibile care sunt reglate pe diferite tipuri și game de semnale. Pentru a clasifica tipurile de electroreceptori, primul punct de diferențiere ar trebui făcut între organele ampulare și tuberoase, care există atât în mormiride, cât și în gimnotiforme. Aceste două tipuri de receptori electrici au diferențe anatomice foarte distincte: organele ampulare au „fântâna” deschisă formată într-o structură asemănătoare canalelor și umplută cu substanță mucoasă; „groapa” unui organ tuberos, pe de altă parte, este umplută slab cu celule epiteliale. Pe lângă diferențele anatomice, acești doi receptori au și diferențe funcționale distincte. Organele ampulare sunt cele mai sensibile și reglate la un interval de frecvență scăzut de 1-10 Hz, care este gama surselor de electricitate non-electrogene și biologice. Prin urmare, organele ampulare sunt utilizate în principal pentru electrolocația pasivă. Pe de altă parte, tuberozele, care sunt utilizate pentru electrocomunicații de pești slab electrici, sunt mai puțin sensibile și reglate la frecvențe mult mai mari. [6] [7]
Clasificarea celor două tipuri de organe receptive [8]
| Tip | Structura | Funcţie | Sensibilitate | Unde este |
|---|---|---|---|---|
| Ampulează | Deschis / Umplut cu mucus | Electro-localizare / Localizare pradă | 0,01 μV / cm la speciile marine, 0,01 mV / cm în apă dulce; sensibil la câmpurile de frecvență continuă / joasă sub 50 Hz | Rechini și raze; pești non-teleost; anumite teleoste (mormiridae, anumite notopteri, gimnotiforme, somn); amfibieni (cu excepția broaștelor și broaștelor) |
| Tuberoasă | Acoperit cu piele - ușor umplut cu celule epiteliale | Electrocomunicații | 0,1 mV la 10 mV / cm / zeci de Hz la mai mult de 1 kHz. | Pește Mormirida; pești gimnotiformi |
Organe tuberoase
Organele tuberoase, tipul de organ receptiv utilizat pentru electrocomunicații, pot fi împărțite în două tipuri, în funcție de modul în care sunt codificate informațiile: codificatori de timp și codificatori de amplitudine. Există multiple forme de organe tuberoase în fiecare codificator de timp și amplitudine, iar toate speciile de pești slab electrici posedă cel puțin o formă a celor doi codificatori. Codificatorul de timp declanșează potențialul de acțiune al blocului de fază (ceea ce înseamnă că forma de undă a potențialului de acțiune este întotdeauna aceeași) la un timp de întârziere fix după formarea fiecărui tranzitoriu extern. Prin urmare, codificatoarele de timp neglijează informațiile despre forma de undă și amplitudine, dar se concentrează pe frecvența semnalului și aruncă potențialele de acțiune pe o bază 1: 1 pe tranzitorul extern. Codificatoarele de amplitudine, pe de altă parte, se declanșează în funcție de amplitudinea descărcării de organe electrice (SOE). Deși atât peștii de tip undă, cât și peștii cu impulsuri au codificatori de amplitudine, aceștia se lansează în moduri diferite: receptorii de pește de tip undă se lansează continuu cu o rată bazată pe amplitudinea lor SOE; pe de altă parte, receptorii impulsivi ai peștilor lansează rafale de vârfuri la fiecare SOE detectat, iar numărul de vârfuri în fiecare rafală este legat de amplitudinea SOE-ului. Electroreceptorii tuberoși prezintă o curbă de reglare a pragului în formă de V (similară cu sistemul auditiv), ceea ce înseamnă că sunt reglați la o anumită frecvență. Această frecvență specială este de obicei strâns legată de frecvența SOE. [9]
Clasificarea organelor tuberoase
| Tip | A doua lansare | Metoda de codificare | Este localizat în |
|---|---|---|---|
| Codificator de timp | Frecvența SOE primită | Lansați potențialul de acțiune într-un raport 1: 1 cu SOE primit | Ambele tipuri de pești slab electrici |
| Codificator de amplitudine | Amplitudinea SOE-ului primit | Tip de undă: lansare continuă la o viteză în funcție de amplitudinea tipului SOE / impuls: numărul de explozii în fiecare vârf depinde de amplitudinea SOE | Ambele tipuri de pești slab electrici |
Organe electrice
Peștii slabi electric generează descărcarea de organe electrice (SOE) cu compartimente specializate numite organe electrice . Aproape toți peștii slab electrici au organe electrice derivate din celule musculare (miogene); singura excepție este reprezentată de apteronotide, o familie de gimnotiforme care are organe electrice derivate din celule nervoase (neurogene). Electrocitele miogene sunt aranjate în coloane de celule mici, asemănătoare unui disc, numite placi electrice. Familia care face excepție, cea a apteronotidelor, transportă organele electrice miogene în stadiile larvelor. Cu toate acestea, pe măsură ce peștele se maturizează, organele electrogene derivate din măduva spinării centrală înlocuiesc treptat celulele electrice derivate din celulele musculare. [10]
Descărcarea unui organ electric începe cu comanda centrală a unui stimulator cardiac medular care determină rata și ritmul SOE. Aceste două caracteristici ( frecvența și ritmul ) SOE sunt, de asemenea, numite secvență de impulsuri (SII). Comanda stimulatorului cardiac medular este apoi transmisă de la neuronii măduvei spinării către electrocitele care formează organul electric, care determină forma de undă a SOE-urilor pe baza proprietăților sale morfofiziologice. Când controlul stimulatorului cardiac ajunge la organul electric, acesta deschide toate canalele de sodiu, provocând un flux net de ioni de sodiu într-o singură direcție. Direcția va fi fie spre sau departe de cap și duce la depolarizarea simultană a tuturor electroicitelor de pe aceeași parte a celulei. Rezultatul este o polaritate pozitivă la capul peștilor față de coadă sau invers: un sistem dipolar . Polaritatea creată de organul electric declanșează apoi un câmp electrostatic în apă. [4] [11]

Organele electrice sunt destul de diferite între mormiride și gimnotiforme și, prin urmare, vor fi prezentate separat.
Mormiridi
La mormiride, organul electric este destul de mic și se găsește numai în regiunea pedunculului caudal (partea îngustă a corpului unui pește unde este atașată aripa caudală). Organele electrice sunt compuse din electrocite în formă de disc conectate în serie între ele în două coloane și fiecare coloană se află pe o parte a măduvei spinării. Electrocitele miogene sunt identice între ele și sunt descărcate în sincronie. Potențialul electric înregistrat de un singur electrocit este echivalent cu versiunea în miniatură a unui SOE complet măsurat în afara peștilor. Electrocitele au, de asemenea, o structură importantă numită "pedicul", care sunt structuri tentaculare sau tubulare care se extind de la fiecare electrocit. Au fost observate mai multe sisteme de plumb de tulpină, care includ pediculi care pătrund în conducte, inervează conductele din partea posterioară sau anterioară. Diversi pediculi dintr-un electrocit se topesc în cele din urmă pentru a forma un pedicul mare care primește inervația de la neuronii spinării-electromotori. Diferite structuri morfologice ale sistemelor pediculare / electrocitale determină diferențe în fluxul de curent electric, care duc în continuare la diferite forme de undă. [4] [9] [12]
Gimnotiforme
La Gymnotiformes, electrocitele diferă între peștii electrici de tip undă și de tip pulsat. La peștii de tip val, electrocitele sunt sub formă tubulară. La peștii cu impuls, electrocitele tind să fie discuri turtite. Electrocitele formează și coloane, dar spre deosebire de dimensiunea mai mică a organului electric din mormiride, gimnotiformele au organe electrice lungi care se extind aproape pe toată lungimea corpului longitudinal. De asemenea, spre deosebire de sistemul pedicular din Mormyrids, pediculii din Gymnotiformes fac un singur tip de inervație pe partea posterioară a electrocitului. Gimnotiformele de tip impulsiv prezintă, în general, o complexitate mai mare decât peștii de tip val. De exemplu, electrocitele lor pot fi cilindrice sau în formă de tambur, cu grinzi inervate din spate sau din față. O altă diferență importantă este că, spre deosebire de mormiride sau gimnotiforme de tip de undă, electrocitele gimnastelor de tip de undă nu sunt omogene de-a lungul lungului organ electric care trece prin corpul peștilor. Diferite părți ale organelor electrice ale unor gimnotiforme sunt inervate diferit sau pot avea proprietăți de activare a celulelor diferite.
Apteronotidele, componente ale gimnotiformelor de tip val, sunt diferite de toți ceilalți pești electrici, deoarece sunt singura familie care posedă electrocite neurogene. Organul electric al apteronotidelor este derivat din neuroni; mai precis, acestea sunt formate din axonii neuronilor măduvei spinării. Această structură elimină un decalaj sinaptic între neuronul electromotor spinal și electrocitele miogene, care ar putea contribui la frecvența mai mare a apteronotidului SOE (> 2000 Hz) în rândul peștilor electrici. [9]
Semnale
Tipuri de semnale
Există două tipuri de semnale generate de peștii electrici: puls și undă. Un SOE pulsat este caracterizat de un impuls SOE discret separat de intervale silentioase relativ lungi, mult mai lungi decât descărcările; dimpotrivă, o undă SOE are perioada de lansare și perioada de tăcere aproximativ egală în lungime și, astfel, se formează un semnal continuu cu o formă de undă aproape sinusoidală . Dintre Mormiridae și Gymnotiformes, atât peștele impulsiv, cât și cel de undă au caracteristici consistente în cadrul diferitelor grupuri. [13]
Proprietățile fizice ale semnalelor
Câmp electric
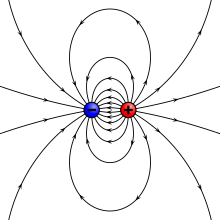
Peștii electrici generează un câmp electrostatic în formă de dipol, cu linii de câmp care descriu un arc curbat de la polul pozitiv la cel negativ. Semnalele de câmp electric diferă de alte moduri de comunicare, cum ar fi sunetul sau optica, care utilizează semnale care se propagă ca unde. În timp ce undele sonore pentru comunicarea acustică sau undele luminoase (unde electromagnetice) pentru comunicațiile vizuale se propagă toate, semnalele electrice nu (ele sunt diferite de undele electromagnetice). Ca un câmp electric, amplitudinea semnalului scade în conformitate cu legea pătratului invers, ceea ce face ca transmiterea și formarea semnalului să fie un proces ridicat al costului energiei . Peștii electrici potrivesc impedanța organului lor electric cu conductivitatea apei pentru a atinge minimul de energie pierdută , iar rezultatul final sunt semnale electrice care circulă cel mult pentru câțiva metri . Deși peștii electrici sunt limitați la un interval de comunicare scurt, semnalele rămân necorupte de ecou și reverberație, care afectează sunetul și lumina. Deteriorarea undelor include reflectarea , refracția , absorbția , interferența și așa mai departe. În consecință, caracteristicile temporale, care sunt foarte importante pentru semnalele electrice ale peștilor, rămân constante în timpul transmisiei. [14]
Spațiu activ
Atunci când se transmit semnale electrice într-un mediu acvatic, natura fizică și chimică a mediului înconjurător poate face diferențe majore în transmiterea semnalului. Factorii de mediu care ar putea impune influențe includ concentrația solutului, temperatura și zgomotul electric de fundal (fulgere sau structuri artificiale) etc. Pentru a înțelege eficacitatea transmiterii semnalului electric, este necesar să se definească termenul „spațiu activ”: zona / volumul în care un semnal poate provoca răspunsuri de la alte organisme. Spațiul activ al unui pește electric are în mod normal o formă elipsoidală datorită aranjamentului dipolilor formați de organele sale electrice. În timp ce atât comunicația electrică, cât și electrolocalizarea se bazează pe semnale generate de organele electrice, electrocomunicația are un spațiu activ de zece ori mai mare decât electrolocația datorită sensibilității extreme a receptorilor tuberoși ai electrocomunicației. [15]
Unul dintre cei mai mari factori care afectează dimensiunea spațiului activ va fi conductanța apei mediată de concentrația solutului din apă. S-a demonstrat că Momiridae și-au adaptat mediul activ optim la habitate cu conductivitate scăzută. Un fenomen natural care susține această teorie este acela că multe specii reproduc în perioada în care râurile / lacurile au cea mai mică conductivitate din cauza precipitațiilor abundente. A avea un spațiu activ mai mare în apă cu conductivitate scăzută va promova, prin urmare, împerecherea și curtarea. [16] O altă explicație testată de Kim și Moller este că, având un spațiu activ mai mic în timpul sezonului uscat, atunci când împerecherea nu are loc, există spațiu social aglomerat, fără transmisie de semnal inutilă între indivizi. [4]
Frecvența și forma de undă
Peștii electrici comunică cu semnale electrice care au două calități principale: frecvența și forma de undă . Informațiile despre forma de undă sunt încorporate în descărcarea organelor electrice (SOE), care este determinată și fixată de anatomia și fiziologia organului electric în sine. Forma de undă a SOE, la unele specii, se modifică odată cu etapele de dezvoltare. Frecvența SOE-urilor și durata intervalului dintre ele se numesc secvență de impulsuri (SII), care sunt controlate de interneuronii de comandă din creierul mediu și medulla oblongată, așa cum se arată în organele electrice . Modificarea IBS produce indicii sociale foarte diferite în rândul peștilor electrici în timpul împerecherii, avertismentului sau identificării. Aceste două proprietăți (forma de undă / SOE și frecvența / SII) sunt utilizate atât de pești de tip undă, cât și de tip impulsiv pentru recunoaștere și comunicare. [17]
Frecvența SOE
Frecvența este numărul de apariții ale unui eveniment recurent pe unitate de timp. Aici, frecvența SOE se referă la frecvența de emisie a unui pește electric. Peștii de tip val efectuează recunoașterea speciilor prin media frecvențelor SOE, care includ frecvențele de bază ale emisiilor și modulația frecvenței care are ca rezultat creșteri, căderi, răsturnări și încetarea frecvențelor SOE. De exemplu, unele specii de Gimnotiformi folosesc „ciripituri” ( ciripituri ), o creștere bruscă a frecvenței, în timpul curtei.
Forma de undă a SOE
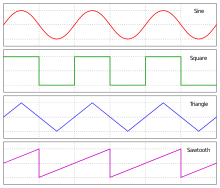
Forma de undă este forma și forma unei unde. Fiecare specie de pește electric are propria sa formă de undă SOE distinctă. Peștii impulsivi efectuează recunoașterea speciilor, acordând atenție diferențelor în forma de undă SOE, care includ proprietăți precum: durata SOE, numărul fazelor și forma fazei. Între timp, unele proprietăți indirecte ascunse în forma de undă sunt folosite și de peștii de tip impulsiv: gradient de amplitudine, raporturi de durată de fază și ordine de fază și semne.
Diferențe și schimbări în semnale
Peștii electrici au în mod normal o frecvență de bază și o formă de undă a semnalelor lor; alterarea ambelor calități are loc în mod continuu, între diferite specii, sexe, stadii de dezvoltare și stări de dominanță. În timp ce mai multe modificări apar pentru a semnaliza generații bazate pe identitățile peștilor, nivelul și tipurile de alterare sunt limitate de sistemul senzorial al peștilor, care este condiționat să perceapă semnale care au o frecvență similară cu frecvența propriilor descărcări. [6]
Semnale și sex
Pe măsură ce peștii electrici se maturizează, unii taxoni dezvoltă diferențe în SOE între bărbați și femele (adică dimorfism sexual ). În general, peștii masculi electrici au o frecvență SOE mai mică și o durată SOE mai mare decât femelele; în rândul masculilor, peștele dominant și cel mai mare posedă în general cea mai mică frecvență. De exemplu, măsurătorile efectuate pe Sternopygus marucus [18] au arătat că masculii generează de obicei SOE la aproximativ 80 Hz, în timp ce femelele generează SOE la aproximativ 150 Hz. Aceste diferențe în SOE între sexe pot fi urmărite la modificări ale potențialului. Acțiunea electrocitelor. Pe măsură ce peștele electric se maturizează din stadiul juvenil, peștii masculi se măresc cu cozile mai lungi și mai groase, ceea ce ar putea implica organe electrice mai mari, generând SOE cu frecvență mai mică.[19] [18] Unul dintre factorii fiziologici care s-au dovedit că contribuie la dimorfismul sexual al SOEs este nivelul hormonului teleostic, al androgenului 11-ketotestosteron (11-KT) și al estrogenului . Experimentele au arătat că prin injectarea 11-KT în peștii de sex feminin, nu numai formele de undă SOE și frecvențele lor s-au apropiat de cele ale masculilor, ci și electroreceptorii lor tuberoși au fost modificați pentru a putea detecta semnalele. . Cu toate acestea, când s-a aplicat estrogen, SOE-urile de pește electric masculin s-au apropiat treptat de SOEs feminine. [6] [20]
Dimorfismul sexual în formele de undă și frecvențele SOE impune, de asemenea, o influență asupra dimensiunii spațiului activ. Folosind Sternopygus marucus ca exemplu, masculii emit frecvențe de aproape jumătate din cele ale femelelor (80 Hz cf. 150 Hz). Cu toate acestea, deoarece majoritatea electroreceptorilor sunt reglați pentru a semnaliza frecvențe mai apropiate de frecvența naturală a receptorilor, diferența de frecvență SOE are ca rezultat o capacitate diferită a peștilor electrici de a percepe semnale de ambele sexe, ceea ce duce în continuare la dimensiuni ale spațiului activ diferit. Așa cum s-a măsurat în Sternopygus marucus de Hagedorn, peștii masculi pot detecta doar femelele într-un interval de 6 cm , în timp ce peștele femel poate detecta peștii masculi într-un interval mult mai larg de 39 cm . Se presupune că această diferență de dimensiune a spațiului activ oferă o șansă mai mare ca femeile să se apropie de potențialele perechi și să aleagă o persoană cu care să se împerecheze. [4][19] [18]
Semne și etape de dezvoltare
Studiile efectuate atât pe Gymnotiformes, cât și pe Mormyrids au arătat că există specii din ambele grupuri care au modificări semnificative în SOE de la larve la adulți. Larvele gimnotiforme au toate SOE care sunt simple, monofazic similare cu o singură perioadă funcția cosinusului și formate cu un spectru foarte larg la un interval de frecvență mai mic. Se observă că, pe măsură ce larvele se maturizează, spectrul de frecvență scade, forma de undă de descărcare devine mai ascuțită, iar formele de undă mai complexe care pot consta în faze multiple înlocuiesc treptat SOE larvară simplă. [21]
Pentru peștii miogeni, această modificare a formei de undă a semnalului are loc cu electrocitele larvare inițiale „fuzionând” împreună pentru a forma electrocite noi cu diferite forme, împreună cu redistribuirea canalelor cu porțile ionice , formarea de noi structuri extracelulare pe electrocite , etc. Unii pești cu impuls dezvoltă, de asemenea, organe electrice accesorii situate pe alte părți ale corpului; aceste organe electrice suplimentare joacă, de asemenea, un rol în adăugarea de faze la SOE. Pentru singurele pești neurogeni cunoscuți până acum, apteronotidele, modificările SOE în timpul procesului de dezvoltare păreau a fi mai dramatice decât cele ale peștilor miogeni, ceea ce poate indica faptul că electrocitele neurogene sunt mai ușor de modificat. Similar cu peștii miogeni, apteronotidele au organul lor electric format din miocite. Pe măsură ce apteronotidele se maturizează, noile electrocite neurogene derivate din neuronii motori spinali înlocuiesc electrocitele miogene. [10]
Au fost propuse două ipoteze cu privire la motivul pentru care semnalele electrice s-au schimbat în timpul etapelor de dezvoltare a peștilor. În primul rând, așa cum s-a menționat mai sus, electroreceptorii peștilor sunt de obicei reglați la un anumit interval de frecvențe. Prin urmare, pentru a face comunicarea eficientă, peștii electrici trebuie să restrângă spectrul larg de frecvențe SOE larvare. În al doilea rând, se știe că electroreceptorii somnului, gimnotiformelor și majorității peștilor pre-teleostici sunt reglați la frecvențe mai mici. Prin urmare, menținerea frecvenței SOE larvare scăzută va crește riscul de a fi detectată de prădători. [6]
Semnalele și starea de dominație
Măsurătorile au arătat că peștele electric, de obicei masculin, care domină, are de obicei o frecvență SOE mai mică și o durată SOE mai lungă. Un experiment a arătat că atunci când doi masculi sunt plasați în același rezervor de pești, ambii pești își cresc SOE în prima perioadă scurtă de timp. Cu toate acestea, după ce a lăsat peștele într-o perioadă întunecată (imitând noaptea), masculul cu o amplitudine SOE mai mare, care este de obicei și masculul cu o construcție mai mare, își va crește în continuare SOE; dimpotrivă, masculul cu dimensiuni mai mici ale corpului / SOE mai mic nu își mărește SOE. [22]
Semnalele speciale
În comunicarea electrică, există câteva tipuri distincte de semnale care servesc unor scopuri speciale, cum ar fi curtarea sau agresiunea. Printre aceste SOE speciale includ „raspamenti“ (rașpele), „ciripituri“ ( țârâituri ) și „netezi accelerare“ (netedă accelerație). Raspingul este o rafală de impulsuri la o frecvență relativ constantă efectuată de unele specii în timpul curtei. Scârțâitul este o creștere sau scădere rapidă a frecvenței. Accelerația uniformă este o perioadă de zeci până la sute de milisecunde pe care viteza SOE-urilor o mărește uniform. Datorită legii conservării energiei , amplitudinea SOE poate scădea cu câteva puncte procentuale, dar modificările generale ale formei de undă și amplitudinii sunt mici. Gimnotiformii masculi emit aceste semnale accelerate în timpul agresiunii și curții. La peștii studiați, dacă curtarea este în regulă și duce la reproducere, peștele electric masculin începe să folosească un alt tip special de SOE - scârțâitul. Zgomotul durează, de asemenea, de la zeci la sute de milisecunde; cu toate acestea, creșterea frecvenței este atât de mare încât electrocitele nu se pot recupera suficient de repede și, prin urmare, ciripitele au o amplitudine foarte mică și o formă de undă care este deviată de la forma de undă originală. [17] [23]
Notă
- ^ a b Masashi Kawasaki, The electric fish , su people.virginia.edu . Adus la 3 decembrie 2011 .
- ^ Map of Life - Convergent evolution online , at mapoflife.org , 2018. Accesat la 28 aprilie 2018 .
- ^ NU Czech-Damal, A. Liebschner și L. Miersch,Electroreception in the Guyana delfin (Sotalia guianensis) , în Proceedings: Biological Sciences , vol. 279, nr. 1729, februarie 2012, pp. 663-8, DOI : 10.1098 / rspb.2011.1127 , PMC 3248726 , PMID 21795271 .
- ^ a b c d e f P. Moller, Electric Fishes: History and Behavior , Springer, 1995, ISBN 978-0-412-37380-0 .
- ^ Bernd Kramer, Electrorecepție și comunicare în pești ( PDF ), în progres în zoologie , vol. 42, G. Fisher, 1996.
- ^ a b c d e Friedrich Ladich, Communication in fishes , Enfield, NH, Science Publishers, 2006, ISBN 978-1-57808-405-0 .
- ^ Carl Hopkins, Electroreception , su nbb.cornell.edu . URL consultato il 5 dicembre 2011 .
- ^ H. Zakon, The electroreceptive periphery , in TH Bullock e WF Heiligenberg (a cura di), Electroreception , New York, John Wiley & Sons, 1986, pp. 103-156.
- ^ a b c Masashi Kawasaki, Chapter 7: Physiology of Tuberous Electrosensory Systems , in Theodore H. Bullock, Carl D. Hopkins, Arthur N. Popper e Richard R. Fay (a cura di), Electroreception , New York, Springer, 2005, ISBN 978-3-642-11481-6 .
- ^ a b MVL Bennett, Electric organs , in WS Hoar e DJ Randall (a cura di), Fish Physiology , Londra, Academic Press, 1971.
- ^ Philip K. Stoddard, Electric Signals and Electric Fishes ( PDF ), su pdfs.semanticscholar.org , 2009. URL consultato il 28 aprile 2018 .
- ^ Carl D. Hopkins, Design features for electric communication ( PDF ), in Journal of Experimental Biology , vol. 202, n. 10, 1999, p. 1217.
- ^ Philip K. Stoddard, Electric signals: predation, sex, and environmental constraints , in Advances in the Study of Behaviour , vol. 31, 2002, pp. 201-242, DOI : 10.1016/S0065-3454(02)80009-2 .
- ^ Carl D. Hopkins, Temporal structure of non-propagated electric communication signals brain behavior and evolution ( PDF ), vol. 28, 1986, p. 43.
- ^ William H. Bossert e Edward O. Wilson, The analysis of olfactory communication among animals ( PDF ), in Journal of Theoretical Biology , vol. 5, n. 3, 1963, pp. 443-469. URL consultato il 3 maggio 2018 (archiviato dall' url originale il 10 agosto 2017) .
- ^ Ann Squire e Peter Moller, Effects of water conductivity on electrocommunication in the weak-electric fish Brienomyrus niger (Mormyriformes) , in Animal Behaviour , vol. 30, n. 2, 1982, pp. 375-382.
- ^ a b Carl D. Hopkins, Neuroethology of electric communication , in Annual Review of Neuroscience , vol. 11, 1988, pp. 497-535.
- ^ a b c M. Hagedorn, The ecology, courtship,and mating of gymnotiform electric fish , in TH Bullock e W. Heiligenberg (a cura di), Electroreception , New York, Wiley, 1986, pp. 497-525.
- ^ a b CD Hopkins, Sex differences in electric signaling in anelectric fish , in Science , vol. 176, 1972, pp. 1036-7.
- ^ AH Bass e CD Hopkins, Shifts in frequency tuning of electroreceptors in androgen-treated mormyrid fish , in Journal of Comparative Physiology , vol. 155, n. 6, pp. 713-724, DOI : 10.1007/BF00611588 .
- ^ GW Max Westby e Frank Kirschbaum, Emergence and development of the electric organ discharge in the mormyrid fish, Pollimyrus isidori , in Journal of Comparative Physiology A , vol. 122, n. 2, pp. 251-271, DOI : 10.1007/BF00611894 .
- ^ CR Franchina, VL Salazar, CH Volmar e PK Stoddard, Plasticity of the electric organ discharge waveform of male Brachyhypopomus pinnicaudatus II. Social effects , in Journal of Comparative Physiology B , vol. 187, n. 1, 2001 p., p. 45.
- ^ Carl Hopkins, Behavioral Evidence for Species Recognition , su nbb.cornell.edu . URL consultato il 6 dicembre 2011 .
Voci correlate
Collegamenti esterni
- ( EN ) Un video del corteggiamento tra un maschio e una femmina di Brienomyrus brachyistius si può vedere qui .
- ( EN ) Metodi per ascoltare i pesci elettrici in casa .