ARN ribozomal 16S
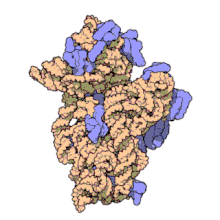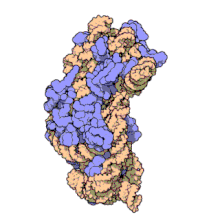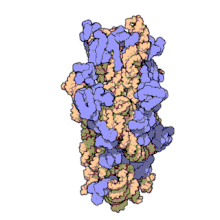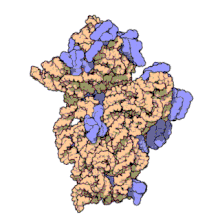
ARN ribozomal 16 S (sau ARNr 16 S ) este o componentă a subunității 30S a unui procariot ribozom care se leagă de secvența Shine-Dalgarno . Genele care o codifică sunt denumite gena ARNr 16S și sunt utilizate în reconstrucția filogeniilor , datorită evoluției lente a acestei regiuni a genei. Carl Woese și George E. Fox au fost cei doi cercetători care au fost pionierii utilizării ARNr-ului 16S pentru cercetarea filogenetică începând cu 1977 . [2]
Funcții
- La fel ca ARN ribozomal mare (23S), joacă un rol structural, acționează ca un schelă care definește pozițiile proteinelor ribozomale.
- Capătul 3 'conține secvența anti -Shine-Dalgarno , care se leagă în amonte codonul de start AUG pe ARNm . Capătul 3 ′ al ARN-ului 16S se leagă de proteinele S1 și S21 despre care se știe că sunt implicate în inițierea sintezei proteinelor [3]
- Interacționează cu 23S, favorizând legarea celor două subunități ribozomale (50S și 30S)
- Stabilizează cuplarea corectă codon-anticodon la locul A prin formarea de legături de hidrogen între atomul N1 al reziduurilor de adenină 1492 și 1493 și gruparea 2'OH a coloanei vertebrale a ARNm
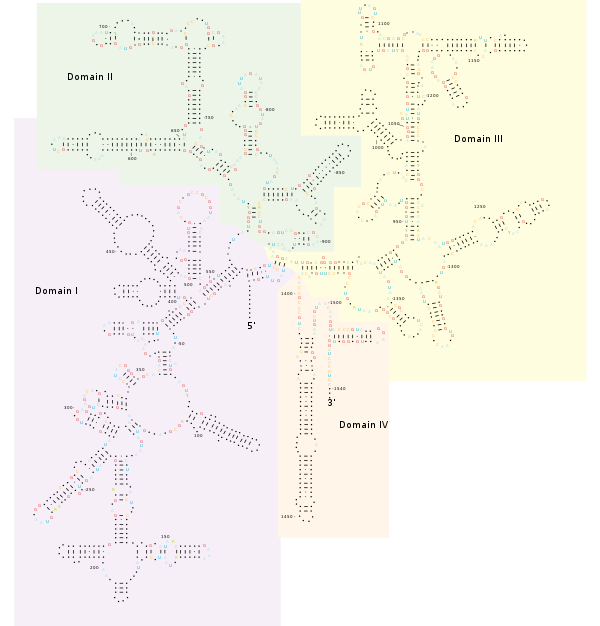
Structura
Aplicații PCR și NGS
În plus față de site-urile de legare a primerului foarte conservate, secvențele genei 16S rARN conțin regiuni hipervariabile care pot furniza secvențe de semnătură specifice speciei utile pentru identificarea bacteriană. [4] [5] În consecință, secvențierea genei 16S rRNA a devenit predominantă în microbiologia medicală ca o alternativă rapidă și rentabilă la metodele fenotipice de identificare a bacteriilor. [6] Deși inițial folosit pentru identificarea bacteriilor, secvențierea 16S s-a dovedit ulterior că poate reclasifica bacteriile în specii complet noi, [7] sau chiar genuri noi. [8] [9] De asemenea, a fost folosit pentru a descrie specii noi care nu au fost niciodată cultivate cu succes. [10] [11] Odată cu secvențierea generației a treia care vine în multe laboratoare, identificarea simultană a mii de secvențe de ARNr 16S este posibilă în câteva ore, permițând studii metagenomice , de exemplu asupra florei intestinale. [12] O aplicație utilă în acest sens pare să fie cercetarea în cercetarea epidemiologică pe bază de ape uzate . [13]
Gena bacteriană 16S conține nouă regiuni hipervariabile (V1-V9), de aproximativ 30 până la 100 de perechi de baze, care sunt implicate în structura secundară a subunității ribozomale mici. [14] Gradul de conservare variază foarte mult între regiunile hipervariabile, cu regiuni mai conservate legate de taxonomie de nivel superior și regiuni mai puțin conservate la niveluri inferioare, cum ar fi genul și speciile. [15] Deși întreaga secvență 16S permite compararea tuturor regiunilor hipervariabile, la o lungime de aproximativ 1.500 de perechi de baze poate fi prohibitiv pentru studiile care caută să identifice sau să caracterizeze diferite comunități bacteriene. Aceste studii utilizează în mod obișnuit platforma Illumina , care produce citiri la viteze de 50 de ori și de 12.000 de ori mai ieftine decât 454 piroza secvențierea și respectiv secvențierea Sanger . [16] Deși mai ieftin și care permite o acoperire comunitară mai profundă, secvențierea Illumina produce citiri de numai 75-250 de perechi de baze lungi (până la 300 de perechi de baze cu Illumina MiSeq) și nu are un protocol stabilit pentru a se asambla în mod corespunzător. probe. [17] Cu toate acestea, regiunile complet hipervariabile pot fi asamblate dintr-o singură cursă Illumina, făcându-le ținte ideale pentru platformă.
Notă
- ^ Structura subunității ribozomale mici activate funcțional la rezoluția de 3,3 angstromi , în Cell , vol. 102, nr. 5, septembrie 2000, pp. 615-23, DOI : 10.1016 / S0092-8674 (00) 00084-2 , PMID 11007480 .
- ^ Către un sistem natural de organisme: propunere pentru domeniile Archaea, Bacteria și Eucarya , în Proceedings of the National Academy of Sciences din Statele Unite ale Americii , vol. 87, nr. 12, iunie 1990, pp. 4576-9, Bibcode : 1990PNAS ... 87.4576W , DOI : 10.1073 / pnas.87.12.4576 , PMID 2112744 .
- ^ AP Czernilofsky, CG Kurland și G. Stöffler, 30S Proteine ribozomale asociate cu capătul 3'-terminal al ARN 16S , în FEBS Letters , vol. 58, nr. 1, 1975, pp. 281-284, DOI : 10.1016 / 0014-5793 (75) 80279-1 , ISSN 0014-5793 , PMID 1225593 .
- ^ Identificarea speciilor prin analiza multiplexă a secvențelor cu lungime variabilă , în Nucleic Acids Research , vol. 38, nr. 22, decembrie 2010, pp. e203, DOI : 10.1093 / nar / gkq865 , PMID 20923781 .
- ^ Secvențierea ADN-ului ribozomal ca instrument pentru identificarea agenților patogeni bacterieni , în Current Opinion in Microbiology , vol. 2, nr. 3, iunie 1999, pp. 299-305, DOI : 10.1016 / S1369-5274 (99) 80052-6 , PMID 10383862 .
- ^ Impactul analizei secvenței genei ARNr 16S pentru identificarea bacteriilor asupra microbiologiei clinice și a bolilor infecțioase , în Clinical Microbiology Reviews , vol. 17, n. 4, octombrie 2004, pp. 840–62, cuprins, DOI : 10.1128 / CMR.17.4.840-862.2004 , PMID 15489351 .
- ^ Transcrierea inversă a ARNr-ului 16S pentru a monitoriza populațiile bacteriene care sintetizează ribozomii din mediu , în Microbiologie aplicată și de mediu , vol. 75, nr. 13, iulie 2009, pp. 4589-98, DOI : 10.1128 / AEM.02970-08 , PMID 19395563 .
- ^ 16S Amplificare ADN ribozomal pentru studiu filogenetic , în Journal of Bacteriology , vol. 173, nr. 2, ianuarie 1991, pp. 697-703, DOI : 10.1128 / jb.173.2.697-703.1991 , PMID 1987160 .
- ^ Burkholderia thailandensis sp. nov., către speciile Burkholderia pseudomallei , în International Journal of Systematic Bacteriology , 48 Pt 1, n. 1, ianuarie 1998, pp. 317-20, DOI : 10.1099 / 00207713-48-1-317 , PMID 9542103 .
- ^ Identificarea filogenetică a agenților patogeni neculturați folosind secvențe de ARN ribozomal , în Methods in Enzymology , vol. 235, 1994, pp. 205–222 , DOI : 10.1016 / 0076-6879 (94) 35142-2 , ISBN 978-0-12-182136-4 .
- ^ Analiza filogenetică a comunităților bacteriene din sedimentele marine , în Microbiologie aplicată și de mediu , vol. 62, nr. 11, noiembrie 1996, pp. 4049-59, PMID 8899989 .
- ^ Secvențierea generației următoare a ampliconilor genei ARN ribozomal 16S , în Journal of Visualized Experiments , n. 90, august 2014, DOI : 10.3791 / 51709 , PMID 25226019 .
- ^ SK Park, CW Lee, DI Park, HY Woo, HS Cheong, HC Shin, K Ahn, MJ Kwon și EJ Joo,Detecția SARS-CoV-2 în probe fecale de la pacienții cu COVID asimptomatic și ușor-19 în Coreea. , în Clinical gastroenterology and hepatology: the official clinic practice journal of the American Gastroenterological Association , 10 iunie 2020, DOI : 10.1016 / j.cgh.2020.06.005 , ISSN 1542-3565 , PMC 7286243 , PMID 32534042 .
- ^ Despre descendența evolutivă a organismelor și organelor: o filogenie globală bazată pe un nucleu structural extrem de conservat în ARN ribozomal de subunitate mică , în Nucleic Acids Research , vol. 12, nr. 14, iulie 1984, pp. 5837-52, DOI : 10.1093 / nar / 12.14.5837 , PMID 6462918 .
- ^ Sensibilitatea și corelația regiunilor hipervariabile în genele ARNr 16S în analiza filogenetică , în BMC Bioinformatics , vol. 17, n. 1, martie 2016, p. 135, DOI : 10.1186 / s12859-016-0992-y , PMID 27000765 .
- ^ Generarea de biblioteci de gene ARNr 16S-secvență multimilionară din comunități microbiene complexe prin asamblarea citirilor iluminate cu capăt asociat , în Microbiologie aplicată și de mediu , vol. 77, nr. 11, iunie 2011, pp. 3846-52, DOI : 10.1128 / AEM.02772-10 , PMID 21460107 .
- ^ O metodă pentru secvențierea de înaltă precizie a genelor ARNr 16S aproape de lungime întreagă pe un Illumina MiSeq , în PeerJ , vol. 4, 20 septembrie 2016, pp. e2492, DOI : 10.7717 / peerj.2492 , PMID 27688981 .