Haplogroup I-M253
Haplogroup I-M253, de asemenea , cunoscut sub numele de I1, este un haplogroup Y cromozom . Markerii genetici confirmați ca identificatori I-M253 sunt SNPs M253, M307.2 / P203.2, M450 / S109, P30, P40, L64, L75, L80, L81, L118, L121 / S62, L123, L124 / S64, L125 / S65, L157.1, L186 și L187. Este o ramură primară a haplogrupului I-M170 (I *).
Haplogrupul atinge cele mai mari frecvențe în Suedia (52% dintre bărbați în județul Västra Götaland ) și vestul Finlandei (peste 50% în provincia Satakunta ) [1] . În ceea ce privește mediile naționale, I-M253 se găsește la 35-38% dintre bărbații suedezi , [2] 32,8% dintre bărbații danezi , aproximativ 31,5% dintre bărbații norvegieni [3] și aproximativ 28% dintre bărbații finlandezi . [4]
haplogrupul I-M253 este o ramură primară a haplogrupului I * (I-M170), prezentă în Europa încă din cele mai vechi timpuri. Cealaltă ramură primară a lui I * este I-M438, cunoscută și sub numele de I2.
Înainte de reclasificare în 2008, grupul era cunoscut sub numele de I1a , un nume care a fost reatribuit de atunci unei ramuri primare, haplogrupul I-DF29. Celelalte ramuri primare ale lui I1 (M253) sunt I1b (S249 / Z131) și I1c (Y18119 / Z17925).
Origini
Potrivit unui studiu publicat în 2010, I-M253 s-a născut între 3.170 și 5.000 de ani în urmă, în Europa calcolitică . [5] Un nou studiu din 2015 a estimat originea între 3 470 și 5 070 de ani în urmă sau între 3 180 și 3 760 de ani în urmă, utilizând două tehnici diferite. [6] S-a sugerat că inițial a fost dispersat din zona care este acum Danemarca. [7]
Un studiu din 2014 realizat în Ungaria a descoperit rămășițele a nouă indivizi din cultura ceramică liniară , dintre care unul a fost găsit purtând M253 SNP care definește haplogrupul I1. Se crede că această cultură a existat între 6.500 și 7.500 de ani în urmă. [8]
Structura
I-M253 ( M253, M307.2 / P203.2, M450 / S109, P30, P40, L64, L75, L80, L81, L118, L121 / S62, L123, L124 / S64, L125 / S65, L157.1, L186 și L187 ) sau I1 [9]
- I-DF29 (DF29 / S438); I1a
- I-CTS6364 (CTS6364 / Z2336); I1a1
- I-M227; I1a1a
- I-L22 (L22 / S142); I1a1b
- I-P109; I1a1b1
- I-L205 (L205.1 / L939.1 / S239.1); I1a1b2
- I-Z74; I1a1b3
- I-L300 (L300 / S241); I1a1b4
- I-L287
- I-L258 (L258 / S335)
- I-L813
- I-L287
- I-Z58 (S244 / Z58); I1a2
- I-Z59 (S246 / Z59); I1a2a
- I-Z60 (S337 / Z60, S439 / Z61, Z62); I1a2a1
- I-Z140 (Z140, Z141)
- I-L338
- I-F2642 (F2642)
- I-Z73
- I-L1302
- I-L573
- I-L803
- I-Z140 (Z140, Z141)
- I-Z382; I1a2a2
- I-Z60 (S337 / Z60, S439 / Z61, Z62); I1a2a1
- I-Z138 (S296 / Z138, Z139); I1a2b
- I-Z2541
- I-Z59 (S246 / Z59); I1a2a
- I-Z63 (S243 / Z63); I1a3
- I-BY151; I1a3a
- I-L849.2; I1a3a1
- I-BY351; I1a3a2
- I-CTS10345
- I-Y10994
- I-Y7075
- I-CTS10345
- I-S2078
- I-S2077
- I-Y2245 (Y2245 / PR683)
- I-L1237
- I-FGC9550
- I-S10360
- I-S15301
- I-Y7234
- I-L1237
- I-Y2245 (Y2245 / PR683)
- I-S2077
- I-BY62 (BY62); I1a3a3
- I-BY151; I1a3a
- I-CTS6364 (CTS6364 / Z2336); I1a1
- I-Z131 (Z131 / S249); I1b
- I-CTS6397 ; I1b1
- I-Z17943 (Y18119 / Z17925, S2304 / Z17937); I1c
Distribuție geografică
I-M253 se găsește la cea mai mare densitate în Europa de Nord și în alte țări care au cunoscut migrații extinse din Europa de Nord, fie în perioada de migrație , fie în perioada vikingă sau în timpurile moderne. Se găsește în toate locurile invadate de vechile popoare germanice și de vikingi.
În epoca modernă, populațiile I-M253 au prins rădăcini și în națiunile imigranților și în fostele colonii europene, cum ar fi Statele Unite, Australia și Canada.
| Populația | Marime de mostra | Eu (total) | I1 (I-M253) | I1a1a (I-M227) | Sursă |
|---|---|---|---|---|---|
| Austria | 43 | 9.3 | 2.3 | 0,0 | Underhill și colab. 2007 |
| Belarus: Vitebsk | 100 | 15 | 1.0 | 0,0 | Underhill și colab. 2007 |
| Belarus: Brest | 97 | 20.6 | 1.0 | 0,0 | Underhill și colab. 2007 |
| Bosnia | 100 | 42 | 2.0 | 0,0 | Rootsi și colab. 2004 |
| Bulgaria | 808 | 26.6 | 4.3 | 0,0 | Karachanak și colab. 2013 |
| Republica Cehă | 47 | 31.9 | 8.5 | 0,0 | Underhill și colab. 2007 |
| Republica Cehă | 53 | 17.0 | 1.9 | 0,0 | Rootsi și colab. 2004 |
| Danemarca | 122 | 39.3 | 32,8 | 0,0 | Underhill și colab. 2007 |
| Anglia | 104 | 19.2 | 15.4 | 0,0 | Underhill și colab. 2007 |
| Estonia | 210 | 18.6 | 14.8 | 0,5 | Rootsi și colab. 2004 |
| Estonia | 118 | 11.9 | Lappalainen și colab. 2008 | ||
| Finlanda (național) | 28.0 | Lappalainen și colab. 2006 | |||
| Finlanda: vest | 230 | 40 | Lappalainen și colab. 2008 | ||
| Finlanda: est | 306 | 19 | Lappalainen și colab. 2008 | ||
| Finlanda: regiunea Satakunta | Peste 50 de ani | Lappalainen și colab. 20089 | |||
| Franţa | 58 | 17.2 | 8.6 | 1.7 | Underhill și colab. 2007 |
| Franţa | 12 | 16.7 | 16.7 | 0,0 | Cann și colab. 2002 |
| Franța (Normandia de Jos) | 42 | 21.4 | 11.9 | 0,0 | Rootsi și colab. 2004 |
| Germania | 125 | 24 | 15.2 | 0,0 | Underhill și colab. 2007 |
| Grecia | 171 | 15.8 | 2.3 | 0,0 | Underhill și colab. 2007 |
| Ungaria | 113 | 25.7 | 13.3 | 0,0 | Rootsi și colab. 2004 |
| Irlanda | 100 | 11 | 6.0 | 0,0 | Underhill și colab. 2007 |
| Tătarii Kazan | 53 | 13.2 | 11.3 | 0,0 | Trofimova 2015 |
| Letonia | 113 | 3.5 | Lappalainen și colab. 2008 | ||
| Lituania | 164 | 4.9 | Lappalainen și colab. 2008 | ||
| Olanda | 93 | 20.4 | 14 | 0,0 | Underhill și colab. 2007 |
| Norvegia | 2826 | 31,5 | Eupedia 2017 | ||
| Rusia (națională) | 16 | 25 | 12.5 | 0,0 | Cann și colab. 2002 |
| Rusia: Pskov | 130 | 16.9 | 5.4 | 0,0 | Underhill și colab. 2007 |
| Rusia: Kostroma | 53 | 26.4 | 11.3 | 0,0 | Underhill și colab. 2007 |
| Rusia: Smolensk | 103 | 12.6 | 1.9 | 0,0 | Underhill și colab. 2007 |
| Rusia: Voronez | 96 | 19,8 | 3.1 | 0,0 | Underhill și colab. 2007 |
| Rusia: Arhanghelsk | 145 | 15.8 | 7.6 | 0,0 | Underhill și colab. 2007 |
| Rusia: cazaci | 89 | 24.7 | 4.5 | 0,0 | Underhill și colab. 2007 |
| Rusia: Kareliani | 140 | 10 | 8.6 | 0,0 | Underhill și colab. 2007 |
| Rusia: Kareliani | 132 | 15.2 | Lappalainen și colab. 2008 | ||
| Rusia: Vepsa | 39 | 5.1 | 2.6 | 0,0 | Underhill și colab. 2007 |
| Slovacia | 70 | 14.3 | 4.3 | 0,0 | Rootsi și colab. 2004 |
| Slovenia | 95 | 26.3 | 7.4 | 0,0 | Underhill și colab. 2007 |
| Suedia (național) | 160 | 35.6 | Lappalainen și colab. 2008 | ||
| Suedia (național) | 38.0 | Lappalainen și colab. 2009 | |||
| Suedia: Västra Götaland | 52 | Lappalainen și colab. 2009 | |||
| elvețian | 144 | 7.6 | 5.6 | 0,0 | Rootsi și colab. 2004 |
| curcan | 523 | 5.4 | 1.1 | 0,0 | Underhill și colab. 2007 |
| Ucraina: Lvov | 101 | 23.8 | 4.9 | 0,0 | Underhill și colab. 2007 |
| Ucraina: Ivanovo-Frankov | 56 | 21.4 | 1.8 | 0,0 | Underhill și colab. 2007 |
| Ucraina: Hmelnitz | 176 | 26.2 | 6.1 | 0,0 | Underhill și colab. 2007 |
| Ucraina: Cherkassy | 114 | 28.1 | 4.3 | 0,0 | Underhill și colab. 2007 |
| Ucraina: Belgorod | 56 | 26,8 | 5.3 | 0,0 | Underhill și colab. 2007 |
Suedia
Danemarca
Norvegia
Finlanda
Marea Britanie
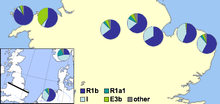
În 2002, a fost publicată o lucrare a lui Michael E. Weale și a colegilor săi care arată dovezi genetice pentru diferențele de populație dintre populațiile engleze și galeze, inclusiv un nivel semnificativ mai ridicat de haplogrup Y-ADN I în Anglia decât în Țara Galilor. Ei au văzut-o ca o mărturie convingătoare a invaziei masive anglo-saxone din estul Marii Britanii din nordul Germaniei și Danemarcei în timpul perioadei migratorii. Autorii presupun că populațiile cu proporții mari de haplogrup I au venit din nordul Germaniei sau din sudul Scandinaviei, în special din Danemarca, și că strămoșii lor ar fi migrat peste Marea Nordului cu migrații anglo-saxone și vikingi danezi. Principala afirmație a cercetătorilor a fost:
Că ar fi nevoie de un eveniment de imigrare anglo-saxon care să afecteze 50-100% din structura genetică masculină din centrul Angliei la acea vreme. Reamintim, totuși, că datele noastre nu ne permit să distingem un eveniment care pur și simplu s-a adăugat la ținuta masculină indigenă engleză centrală din India în care bărbații indigeni au fost strămutați în altă parte sau unul în care bărbații indigeni au fost reduși în număr ... Acest studiu demonstrează că granița din Țara Galilor era mai mult o barieră genetică pentru fluxul genetic anglo-saxon al cromozomului Y decât Marea Nordului ... Aceste rezultate indică faptul că o graniță politică poate fi mai importantă decât una geofizică în structurarea genetică a populației .

În 2003, a fost publicat un articol al lui Christian Capelli și colegii care susțin, dar au modificat, concluziile lui Weale și ale colegilor. Această lucrare, care a probat Marea Britanie și Irlanda pe o grilă, a găsit diferențe minore între campionii galezi și englezi, cu o scădere treptată a frecvenței haplogrupului I pe măsură ce se deplasează spre vest spre sudul Marii Britanii. Rezultatele au sugerat autorilor că invadatorii nordici vikingi ar fi influențat puternic zona de nord a insulelor britanice, dar că atât campionii englezi, cât și cei scoțieni (de pe insula principală) au toți o influență germană / daneză.
Membri importanți ai I-M253
Alexander Hamilton , prin genealogia și analizele descendenților săi (presupunând o paternitate regală corespunzătoare genealogiei sale), a fost plasat în cadrul haplogrupului I-M253 din Y-ADN. [10]
Birger Jarl , „Ducele Suediei” al Casei Goților din Bjalbo, fondatorul Stockholmului , ale cărui rămășițe îngropate în biserică au fost testate în 2002 și s-a dovedit a fi și I-M253
Pasagerii Mayflower William Brewster , Edward Winslow și George Soule pentru testul ADN
Marcatori

Ulterior, sunt raportate specificațiile tehnice cunoscute pentru mutațiile SNP și STR ale haplogrupului I-M253.
Nume: M253
Tip: SNP
Nume: M30
Tip: SNP
Nume: P30
Tip: SNP
Nume: P40
Tip: SNP
Notă
- ^ T. Lappalainen, V. Laitinen, E. Salmela, P. Andersen, K. Huoponen, M.-L. Savontaus și P. Lahermo, Migration Waves to the Baltic Sea Region , în Annals of Human Genetics , vol. 72, nr. 3, 2008, pp. 337-348, DOI : 10.1111 / j.1469-1809.2007.00429.x , PMID 18294359 .
- ^ T. Lappalainen, U. Hannelius, E. Salmela, U. von Döbeln, CM Lindgren, K. Huoponen, M.-L. Savontaus, J. Kere și P. Lahermo, Structura populației în Suedia contemporană: o analiză a ADN-ului cromozomial și mitocondrial , în Annals of Human Genetics , vol. 73, nr. 1, 2009, pp. 61–73, DOI : 10.1111 / j.1469-1809.2008.00487.x , PMID 19040656 .
- ^ Eupedia , „Distribuția haplogrupurilor de ADN-cromozom european Y (ADN-Y) după țară în procente” (31 ianuarie 2017).
- ^ Lappalainen T., Koivumäki S., Salmela E., Huoponen K., Sistonen P., Savontaus ML, Lahermo P.; 2006, „Diferențe regionale între finlandezi: o perspectivă cromozomială Y”, Gene vol. 376, nr. 2, pp. 207-15.
- ^ Pedro Soares, Alessandro Achilli, Ornella Semino, William Davies, Vincent Macaulay, Hans-Jürgen Bandelt, Antonio Torroni și Martin B. Richards, The Archaeogenetics of Europe, Current Biology , vol. 20 (23 februarie 2010), R174 - R183. yDNA Haplogroup I: Subclade I1 , Family Tree DNA,
- ^ TMRCA-urile marilor haplogrupuri din Europa estimate folosind două metode. : Extinderea recentă la scară largă a patriliniilor europene, arătată de resecvențierea populației: Nature Communications: Nature Publishing Group , la www.nature.com . Adus pe 19 mai 2015 .
- ^ Peter A. Underhill și colab., New Phylogenetic Relationsies for Y-chromosome Haplogroup I: Reappraising its Phylogeography and Prehistory, în Rethinking the Human Revolution (2007), pp. 33-42.
- ^ Urmărirea originii genetice a primilor fermieri europeni dezvăluie informații despre organizarea lor socială , la biorxiv.org .
- ^ ISOGG, Y-DNA Haplogroup I and its Subclades - 2017 (31 ianuarie 2017).
- ^ Părintele fondator DNA , la isogg.org .
Proiecte
- Bazele de date Haplogroup I.
- Proiect Haplogroup I1 la FTDNA
- Proiectul DNA Danez regional ADN la FTDNA
- Proiect Haplogroup I-P109
- British Isles DNA Project
- Baze de date generale Y-ADN
Există mai multe baze de date cu acces public care includ I-M253, inclusiv: