Haplogrupuri ale cromozomului Y

În genetică umană , haplogrupurile cromozomului Y sunt grupări de combinații de markeri ( haplotipuri ) definite de diferențele din regiunea nerecombinantă a ADN-ului cromozomului Y (numit NRY din cromozomul Y care nu este recombinat ). Aceste diferențe se referă la polimorfisme bialelelice (SNP, polimorfisme cu nucleotide unice ).
Consorțiul YCC Y Chromosome a stabilit un sistem pentru definirea haplogrupurilor cromozomului Y pe baza literelor de la A la T, cu diviziune suplimentară folosind cifre și litere de subindice.
Cromozomul ancestral Y (definit în glumă de către savanți ca „Adam”) este cel care a aparținut unui bărbat teoretic care reprezintă cel mai recent strămoș comun (MRCA Most Recent Common Ancestor ) dintre toți masculii actuali de-a lungul liniei patriliniare, de când cromozomul Y se transmite doar de la tată la fiii săi.
Estimările timpului în care a trăit acest individ teoretic variază între studii.
Arborele filogenetic al umanității (cromozomul Y)
Y , așa-numitul „ Adam ancestral ”
- A00 (AF6): A fost descoperit într-o familie afro-americană , se află în Camerun și are o vechime de aproximativ 235.000 de ani. [1]
- A0 (V148): întâlnit în Camerun , Algeria și Ghana . [2]
- A1 (P305)
- A1a (M31): dispersat în Africa de Vest și rar în Europa de Nord. [3]
- A1b (L419): prezent în Africa subsahariană , în special în rândul boșimanilor (din grupul etnic Khoisan ) [4] și al popoarelor nilotice ( Sudanul de Sud ).
- BT (M42, M91, M94)
- B (M60, M181, P85, P90): răspândit în mare parte din Africa subsahariană , în special în rândul pigmeilor [4] și Hadza .
- CT (M168) a apărut acum aproximativ 88.000 de ani. [5]
- DE (M1 / YAP)
- E (M40, M96, P29): acoperă cea mai mare parte a Africii . [6] Prezent și în Orientul Apropiat și în sudul Europei , în special în Peloponez . [7]
- D (CTS3946): tipic sudului Japoniei , Tibetului și Insulelor Andaman . [8]
- CF (P143)
- C (M130 / RPS4Y711, M216): prezent în principal în Asia de Est și Oceania.
- C1 (F3393, K30): nu foarte frecvent în Japonia , [9] prezent în insulele Pacificului , inclusiv Polinezia , Micronezia și Melanesia ; prezent în frecvență înaltă la australienii aborigeni , [10] prezenți și în Asia de Sud , Asia Centrală și Europa la frecvențe joase.
- C2 (M217): prezent într-o vastă zonă geografică între Siberia , Asia Centrală , Asia de Est și America de Nord .
- F (M89, P14)
- F *: prezent mai ales în popoarele tribale indigene din India . [11]
- G (M201, U2): cele mai mari frecvențe se găsesc în Caucaz , dar provin din Asia de Vest. [12] Prezent și în Orientul Mijlociu și sudul Europei .
- H (M69, M370): întâlnit în principal în subcontinentul indian [13] și la țigani .
- IJK (L15, L16)
- IJ (M429 / P125, S2)
- J (12f2.1, M304, S6): cel mai important dintre popoarele din Orientul Apropiat . Este haplogrupul popoarelor semite. [7]
- I (M170, M258, P19): răspândit în principal în Europa , descendent din triburile proto europene. [14]
- I1 (M253, L64, P30) ramură nordică / nordică
- I2 (M438 / S31, L68) ramura sudică / balcanică
- K (M9)
- LT (L298)
- L (M11, M20): răspândit în principal în Asia de Sud , [15] în special în Afganistan [16] și frecvențe mici în populațiile din Asia Centrală, Orientul Apropiat și Europa.
- T (M184) difuzat în principal în zonele cele mai sudice ale Italiei , în Orientul Apropiat , în sudul Indiei și în Cornul Africii . Clada sa principală este T-M70.
- K2 sau MNOPS (M526)
- NU (M2335)
- N (M231): prezent până în Orientul Îndepărtat și în Siberia , comun în rândul popoarelor urale . [17]
- O (M175, P186): predominant în Asia de Est și Asia de Sud-Est , cu o frecvență apropiată de 75%. [18]
- MS (P397): caracteristic Oceania și parte a Insulindiei .
- M (P256): predominant în Melanesia . [19]
- S (P405, B254): tipic pentru Oceania, comun cu nativii din Australia , insulele Pacificului și Negritosii nativi din Filipine. [20]
- P (PF5850)
- P *: puțin în Asia de Sud-Est. [21]
- P2 (F20148): în Filipine. [22]
- P1 (M45, 92R7, M74, P27.1)
- Q (M242): este principalul haplogrup în aproape toți nativii americani [23] și în rândul asiaticilor.
- R (M207 / UTY2, M306 / S1, P224)
- R1 (M173)
- R1a (M420 / L146, L62): este predominant în populațiile slave din estul Europei și în regiunea Pamir între Asia Centrală și de Sud. [24]
- R1b (M343): este predominant în Europa atlantică [25] , unde este cel mai răspândit haplogrup și în nordul Camerunului .
- R2 (M479): este important în subcontinentul indian . [26]
- R1 (M173)
- NU (M2335)
- LT (L298)
- IJ (M429 / P125, S2)
- C (M130 / RPS4Y711, M216): prezent în principal în Asia de Est și Oceania.
- DE (M1 / YAP)
Migrații
Oamenii moderni, conform studiilor cromozomiale Y, au provenit din Africa subsahariană și apoi au colonizat Eurasia (acum aproximativ 70.000 de ani) [27] urmând coasta de sud a Asiei. [28] Grupurile care au plecat din Africa au fost ulterior dispersate conform următoarei hărți:

Relația cu alte specii umane
Studiul paleontologic al neanderthalienilor a arătat că haplotipul cromozomial Y al Neanderthalului ar fi dispărut și divergența dintre neanderthalieni și oamenii moderni s-ar fi produs acum aproximativ 590.000 de ani. [29] Divergența cu Homo a lui Denisova , pe de altă parte, s-ar fi produs cu aproximativ 700.000 de ani în urmă. [30]
Analiza haplogrupurilor mitocondriale umane a dat rezultate foarte similare, atât de mult încât specia umană analizată prezintă următoarele relații: [31]
|
Principalele haplogrupuri
Principalele haplogrupuri ale cromozomului Y includ:
Haplogrupurile A și B.
Strămoșul comun al tuturor filiațiilor este Adam cromozomial cu 275.000 de ani. [32] Cele mai vechi descendențe ale cromozomilor Y sunt paragrupul A și haplogrupul B și se găsesc în special în Africa Subsahariană sau în populații cu aceeași origine, cum ar fi, de exemplu, afro-americani aduși în străinătate cu comerțul cu sclavi . Cele mai importante frecvențe ale paraphyletic grupa A sunt găsite printre Bushmen , Khung și sudanez . [33] Cele mai ridicate frecvențe ale B printre Biaka și Mbuti pigmeii . După cum se poate observa din arborele filogenetic al haplogrupurilor cromozomului Y, toată diversitatea modernă a fost generată în Africa, astfel încât populațiile contemporane sunt descendenți ai africanilor care au rămas în Africa sau ai acelor populații care au emigrat din Africa pentru a popula restul. a continentelor.
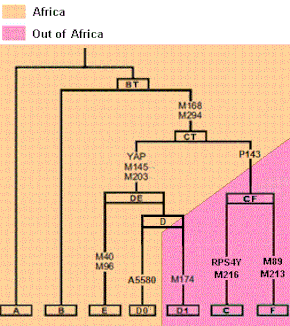
Haplogrupul A este ancestral și parafiletic. Relația sa cu B merge în conformitate cu următorul arbore:
| Adam - Y |
| ||||||||||||||||||||||||||||||||||||
Haplogroup DE (D și E)
Mutațiile M168 și M294 (CT), absente în A și B, definesc toate haplogrupurile de la C la T. Aceste mutații au precedat migrația din Africa , fiind prezente atât în Africa, cât și dincolo. Mutațiile care caracterizează DE (M1 / YAP, M145) s-au produs în Africa acum mai bine de 76.000 de ani. [34] Haplogroup E a rămas inițial în Africa și cele mai frecvente subclade ale sale sunt E1b1a și E1b1b. E1b1a-M2 (denumită anterior E3a) are cea mai mare frecvență și cea mai mare diversitate din vestul Africii subsahariene (peste 80%). [35] Pe de altă parte, sub-clada E1b1b (M215), fostă E3b, este de origine africană, este comună în vorbitorii de limbă afro-asiatică , cea mai mare diversitate a acesteia se găsește în Etiopia și este dispersată în toată Marea Mediterană, ajungând la cea mai înaltă frecvențe în Cornul Africii și Africa de Nord, fiind de 81% în Somalia; [36] și frecvența de 27% în Grecia. Haplogrupa D se găsește numai în Himalaya și în foarte puține zone din sudul Japoniei, unde a fost introdus de primii colonizatori. Mutații tipice care definesc haplogrupurile menționate în text:
Haplogroup C.
Haplogroup C este definit de M130 / RPS4Y și M216, are origine asiatică și are o vechime de aproximativ 53.000 de ani. [37] Dintre populațiile native, cele mai mari frecvențe ale ramurii C1 (F3393) sunt în părți din Oceania , așa cum se poate observa în Polinezia centrală și Australia , deoarece C1 s-a dispersat în Asia de Sud, îndreptându-se în principal spre sud-est, care este legat la prima colonizare a Asiei de Sud-Est și a continentului Sahul . [38]
Ramura C2 se răspândește în Asia de Nord și este deosebit de frecventă în popoarele cu limbă altaică și na dené . [39]
Haplogroup F (FT)
Haplogroup F (M89) a apărut cu aproximativ 66.000 de ani în urmă, [40] probabil în subcontinentul indian [41], deoarece aici se găsește cea mai mare diversitate, cu F1 (P91) în Sri Lanka, F3 (M481) în sud-vestul Indiei [42] ] și F2 / F4 (Y27277) în Extremul Orient.
Haplogrupurile descendente din haplogrupul F reprezintă 90% din populația lumii, dar sunt distribuite aproape exclusiv în afara Africii subsahariene. IJ corespunde probabil unui val migrator din Orientul Mijlociu sau Asia de Vest începând cu 45 000 de ani în urmă, care s-a răspândit apoi în Europa cu omul Cro-Magnon . Haplogroup G , care și-a luat originea în Orientul Mijlociu, sau poate mai la est în Pakistan, acum aproximativ 30 000 de ani, potrivit unor studii, s-ar fi putut răspândi în Europa în neolitic sau, având în vedere discontinuitatea sa puternică, au ajuns deja în Europa în paleoliticul . Haplogrupul H a luat naștere probabil în India acum 30-40.000 de ani, unde a persistat până când s-a răspândit în timpurile istorice cu țiganii. Mutații care caracterizează haplogrupurile menționate în secțiune:

| Haplogroup F. |
| ||||||||||||||||||||||||||||||
Haplogroup I
Haplogroup I (M170) reprezintă aproximativ o cincime din cromozomii Y europeni. Este aproape exclusiv pentru Europa, prin urmare, se crede că a provenit din această zonă. Cel mai vechi pe care l-am găsit (I *) este cel al unui individ cunoscut sub numele de Krems WA3 ( Austria Inferioară ), datând de acum aproximativ 31.000 de ani. [43] Gemeni au fost, de asemenea, găsiți în același loc, ambii atribuiți haplogrupului I *. [44] Este probabil că a fost limitat la refugiul balcanic în timpul glaciației și apoi s-a răspândit în nord odată cu retragerea ghețarilor. În ciuda faptului că sunt relativ frecvente în populațiile scandinave, sarde și balcanice, aceste popoare au diferite subclade ale haplogrupului I. Acest lucru sugerează că fiecare dintre populațiile ancestrale este acum dominată de o anumită subcladă care a marcat o expansiune independentă a populației de-a lungul diferitelor rute migratoare în timpul și imediat după glaciație .
Principalele subclade ale haplogrupului I sunt:
I1 ( M253 , M307, P30, P40) (fostul I1a) cu cele mai mari frecvențe în Scandinavia, Islanda și nord-estul Europei. În Insulele Britanice, mutația I1-M253 este adesea utilizată ca marker pentru invaziile vikinge sau anglo-saxone.
I2 (S31) (fostul I1b) care include I2a1 (P37.2) care este cea mai comună formă în Balcani și Sardinia (unde reprezintă cel mai vizibil haplogrup cu un procent de 38% în rândul bărbaților sarde în varianta I2a1a1, M26 ) și I2a2 (P214) care ating frecvențe discrete de-a lungul coastelor nord-vestice ale Europei continentale cu un vârf în Germania. I2a2a1 (M284) tipic insulelor britanice este derivat din linia I2a2.
Haplogroup K.
Haplogroup K își are originea probabil în Asia de Sud [45] și de acolo s-a răspândit în toată lumea.
Din aceasta au fost generate două clade principale: LT (L298), răspândită acum în Orientul Mijlociu și întâlnită în principal în sudul Asiei , zone în care sunt de frecvență considerabilă; și K2 sau MNOPS (M526), care ar fi fost înființate mai devreme în Asia de Sud-Est , deoarece această regiune își formează centrul de expansiune. [46]
Haplogroup K se caracterizează prin următoarele haplotipuri și următoarele mutații:
| Haplogroup K (M9) |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
Haplogroup M este răspândit în insulele Papua Noua Guinee . Haplogrupurile N și O au apărut acum 35-40.000 de ani în Asia de Est .
Haplogrupul N a apărut probabil în Mongolia și s-a răspândit în Orientul Îndepărtat și Siberia , precum și în vest, fiind cel mai comun grup din rândul popoarelor urale. Haplogroup O se găsește în Extremul Orient. Haplogrupul P se găsește în cea mai mare parte ca subtipuri Q sau R (vezi mai jos), rareori nediferențiate. Probabil a evoluat în Asia Centrală sau în regiunea Altai . Haplogroup Q își are originea și în Asia de Nord , a migrat spre est și a ajuns în America de Nord prin strâmtoarea Bering .
Haplogroup R
Toate haplotipurile aferente haplogrupului R împărtășesc mutațiile M207 (UTY2), M306 (S1), S4, S8, S9 și pot fi împărțite în trei linii evolutive principale: R1a, R1b și R2.
R1a ar fi putut proveni din vecinătatea Iranului cu aproximativ 25.000 de ani în urmă. [47] Este asociat cu cultura Kurgan , cunoscută pentru domesticirea calului (acum aproximativ 5 000 de ani). Această linie este prezentă în prezent în Asia Centrală , India și în populațiile slave din Europa de Est.
Linia R1b este cea mai frecventă la populațiile europene. În vestul Irlandei atinge o frecvență apropiată de 100%. A apărut înainte de sfârșitul ultimei ere glaciare din Asia de Vest . [48]
Nu există dovezi ale existenței R1b în Europa înainte de neolitic ; primul eșantion european de R1b a fost găsit în Spania la Els Trocs acum aproximativ 7 000 de ani [49] , dar R1b prezent în prezent în Europa de Vest nu coboară din clada sa (V88, în prezent aproape complet asimilată). De fapt, se presupune că R1b european a sosit odată cu expansiunile indo-europene din sudul Rusiei, întrucât diferite culturi R1b-L23 (principala cladă R1b răspândită în Europa) au fost găsite în cultura Yamna . [49]
R2 este comun în subcontinentul indian. Prezent și în Orientul Apropiat, Caucaz și Asia Centrală.
Notă
- ^ A00 Y full.tree 2012-2020 YFull
- ^ Scozzari R, Massaia A, D'Atanasio E, Myres NM, Perego UA, și colab. (2012) Disecția moleculară a cladelor bazale în arborele filogenetic al cromozomului Y uman. PLoS ONE 7 (11): e49170. doi: 10.1371 / journal.pone.0049170
- ^ King, TE, Parkin, EJ, Swinfield, G., Cruciani, F., Scozzari, R., Rosa, A., Lim, SK, Xue, Y., Tyler-Smith, C. și Jobling, MA ( 2007). Africani în Yorkshire? Cea mai adâncă înrădăcinare a filogeniei Y într-o genealogie engleză. Revista europeană de genetică umană : EJHG, 15 (3), 288-293. https://doi.org/10.1038/sj.ejhg.5201771
- ^ a b Elizabeth Wood și colab. 2005 Modele contrastante ale variației cromozomului Y și mtDNA în Africa: dovezi pentru procesele demografice orientate spre sex European Journal of Human Genetics 13 , 867-876. doi: 10.1038 / sj.ejhg.5201408
- ^ Y-full CT tree Y Tree v9.02.00 2021 YFull
- ^ Tatiana Karafet și colab., 2008, Noi polimorfisme binare se remodelează și cresc rezoluția arborelui haplogrupului cromozomial Y uman.
- ^ a b Ornella Semino, Chiara Magri, Giorgia Benuzzi, Alice A. Lin, Nadia Al-Zahery, Vincenza Battaglia, Liliana Maccioni, Costas Triantaphyllidis, Peidong Shen, Peter J. Oefner, Lev A. Zhivotovsky, Roy King, Antonio Torroni, L. Luca Cavalli-Sforza, Peter A. Underhill și A. Silvana Santachiara-Benerecetti, Originea, difuzia și diferențierea haplogrupurilor E-J ale cromozomului Y: inferențe despre neolitizarea Europei și evenimente migratorii ulterioare în zona mediteraneană , în Revista Americană de Genetică Umană , vol. 74, nr. 5, -, pp. 1023-1034, DOI : 10.1086 / 386295 , PMC PMC1181965 , PMID 15069642 .
- ^ Shi H, Zhong H, Peng Y și colab. (2008). „Dovezile cromozomului Y ale primei așezări umane moderne din Asia de Est și originile multiple ale populațiilor tibetane și japoneze”. BMC Biol. 6: 45. doi: 10.1186 / 1741-7007-6-45. PMID 18959782 . PMC 2605740. https://www.biomedcentral.com/1741-7007/6/45 .
- ^ Y-DNA Haplogroup C și subclasele sale - 2010
- ^ Georgi Hudjashov, Toomas Kivisild, Peter A. Underhill și colab., "Dezvăluind așezarea preistorică a Australiei prin cromozomul Y și analiza mtDNA", PNAS, vol. 104, nr. 21, pp. 8726–8730 (22 mai 2007)
- ^ Cordaux, Richard et al 2004, Originile independente de indian castă și linii paterne tribale depuse la 8 iunie 2011 , la Internet Archive .
- ^ Y-DNA Haplogroup G și subclasele sale - 2010
- ^ R. Spencer Wells și colab.: "The Eurasian Heartland: O perspectivă continentală asupra diversității cromozomilor Y". Proceedings of the National Academy of Sciences of the United States of America v.98 (18); 28 august 2001
- ^ Siiri Rootsi, Toomas Kivisild, Giorgia Benuzzi, Hela Help, Marina Bermisheva, Ildus Kutuev, Lovorka Barać, Marijana Peričić, Oleg Balanovsky, Andrey Pshenichnov, Daniel Dion, Monica Grobei, Lev A. Zhivotovsky, Vincenza Battaglia, Alessandro Achilli, Nadia Aladia -Zahery, Jüri Parik, Roy King, Cengiz Cinnioğlu, Elsa Khusnutdinova, Pavao Rudan, Elena Balanovska, Wolfgang Scheffrahn, Maya Simonescu, Antonio Brehm, Rita Goncalves, Alexandra Rosa, Jean-Paul Moisan, Andre Chaventre, Vladimir Ferak, Sandor Für Für , Peter J. Oefner, Peidong Shen, Lars Beckman, Ilia Mikerezi, Rifet Terzić, Dragan Primorac, Anne Cambon-Thomsen, Astrida Krumina, Antonio Torroni, Peter A. Underhill, A. Silvana Santachiara-Benerecetti, Richard Villems, Chiara Magri și Ornella Semino, Phylogeography of Y-Chromosome Haplogroup I I Reveals Distinct Domenins of Prehistoric Gene Flow in Europe , in The American Journal of Human Genetics , vol. 75, nr. 1, -, pp. 128–137, DOI : 10.1086 / 422196 .
- ^ Wells RS, Yuldasheva N, Ruzibakiev R, și colab. (August 2001). „Inima eurasiatică: o perspectivă continentală asupra diversității cromozomilor Y”. Proc. Natl. Acad. SUA 98 (18): 10244-9. doi: 10.1073 / pnas.171305098. PMID 11526236 . PMC 56946. http://www.pnas.org/cgi/pmidlookup?view=long&pmid=11526236 .
- ^ Haber Marc, Platt DE, Ashrafian Bonab M, Youhanna SC, Soria-Hernanz DF și colab. 2012 Grupurile etnice din Afganistan împărtășesc un patrimoniu cromozomial Y structurat de evenimente istorice PLoS ONE 7 (3): e34288. doi: 10.1371 / journal.pone.0034288
- ^ Siiri Rootsi, Lev A Zhivotovsky, Marian Baldovič și colab., "O cale nordică în sens invers acelor de ceasornic a haplogrupului N-cromozom Y din Asia de Sud-Est spre Europa", European Journal of Human Genetics (2007) 15, 204-211
- ^ Harta grupurilor Y Haplogroups , la scs.uiuc.edu . Adus la 30 aprilie 2007 (arhivat din original la 28 iulie 2004) .
- ^ Kayser M, Brauer S, Weiss G, Schiefenho¨vel W, Underhill P, Shen P, Oefner P, Tommaseo-Ponzetta M, Stoneking (2003) Cromozom Y redus, dar nu ADN mitocondrial, diversitate în populațiile umane din vestul nou Guinea Am J Hum Genet 72: 281-302
- ^ Y-DNA Haplogroup S și subclasele sale - 2019-2020 ISOGG Y-DNA Haplogroup Tree
- ^ Haplogroup P YTree v8.09.00 2012-2020 YFull
- ^ Y-DNA Haplogroup P și subclasele sale - 2019-2020 ISOGG 2020
- ^ Maria-Catira Bortolini și colab. 2003 Evidența cromozomului Y pentru diferite istorii demografice antice în America
- ^ Peter A Underhill și colab. 2009, Separarea coancesterii post-glaciare a cromozomilor Y europeni și asiatici în cadrul haplogrupului R1a
- ^ Haplogrupuri bialelice cromozomiale Y
- ^ Manoukian, Jean-Grégoire (2006). O sinteză a Haplogroup R2
- ^ Karafet, Tatiana și colab . (2008), Noi polimorfisme binare se remodelează și cresc rezoluția arborelui haplogrupului cromozomial Y uman , Genome Research , DOI : 10.1101 / gr. 7172008
- ^ Kivisild et al 2003, Moștenirea genetică a primilor coloniști persistă atât în populațiile tribale indiene, cât și în cele de castă
- ^ Mendez, FL, Poznik, GD, Castellano, S. și Bustamante, CD (2016). Divergența cromozomilor Y umani moderni neandertali și moderni . Revista americană de genetică umană, 98 (4), 728-734.
- ^ Martin Petr, Mateja Hajdinjak, Qiaomei Fu, Elena Essel, Hélène Rougie și colab. 2020,Istoria evolutivă a cromozomilor Y din Neanderthal și Denisovan . Știință 25 septembrie 2020: Vol. 369, Ediția 6511, pp. 1653-1656 DOI: 10.1126 / science.abb6460
- ^ Y-DNA Haplogroup A și subclasele sale - 2019-2020 ISOGG (Societatea Internațională de Genealogie Genetică)
- ^ Fernando L. Mendez, G. David Poznik, Sergi Castellano și Carlos D. Bustamante (2016) The Divergence of Neandertal and Modern Human Y Chomosomes Yurnal american of human genetics, 98 (4), 728-734. https://doi.org/10.1016/j.ajhg.2016.02.02
- ^ Wood, E., Stover, D., Ehret, C. și colab. Modele contrastante ale variației cromozomului Y și ADNmt în Africa: dovezi pentru procesele demografice orientate spre sex. Eur J Hum Genet 13, 867-876 (2005). https://doi.org/10.1038/sj.ejhg.5201408
- ^ Haber M, Jones AL, Connell BA, Asan, Arciero E, Yang H, Thomas MG, Xue Y, Tyler-Smith C. A Rare Deep-Rooting D0 African Y-Chromosomal Haplogroup și implicațiile sale pentru extinderea oamenilor moderni din Africa. Genetica. 2019 aug; 212 (4): 1421-1428. doi: 10.1534 / genetics.119.302368. Epub 2019 13 iunie. PMID: 31196864; PMCID: PMC6707464.
- ^ Sims LM, Garvey D, Ballantyne J. Subpopulațiile din cadrul haplogrupurilor majore derivate din Europa și Africa R1b3 și E3a sunt diferențiate de SN-Y-uri nedefinite anterior filogenetic. Hum Mutat. 2007 ianuarie; 28 (1): 97. doi: 10.1002 / humu.9469. PMID: 17154278.
- ^ Sanchez, J., Hallenberg, C., Børsting, C. și colab. Frecvențe înalte ale descendențelor cromozomului Y caracterizate prin E3b1, DYS19-11, DYS392-12 la bărbații somalezi. Eur J Hum Genet 13, 856-866 (2005). https://doi.org/10.1038/sj.ejhg.5201390
- ^ Hallast, P., Agdzhoyan, A., Balanovsky, O. și colab. O origine din Asia de Sud-Est pentru cromozomii Y umani non-africani actuali. Hum Genet 140, 299-307 (2021). https://doi.org/10.1007/s00439-020-02204-9
- ^ Amy Werner 2015, cercetătorii genografici din Australia descoperă ramuri unice ale arborelui genealogic uman Redacția National Geographic Society
- ^ Petit, F., Minnai, F., Chiaroni, J. și colab. Extinderea radială a polimorfismelor sistemului grupului sanguin Diego în Asia: semn al co-migrației cu cuceririle mongole. Eur J Hum Genet 27, 125-132 (2019). https://doi.org/10.1038/s41431-018-0245-9
- ^ F Y-full tree Haplogroup YTree v8.09.01 (2020) YFull ™
- ^ Sengupta, Sanghamitra și colab. „Polaritatea și temporalitatea distribuțiilor cromozomilor y de înaltă rezoluție în India identifică atât expansiunile indigene, cât și cele exogene și dezvăluie influența genetică minoră a pastorilor din Asia Centrală”. Revista americană de genetică umană vol. 78,2 (2006): 202-21. doi: 10.1086 / 499411
- ^ Y-DNA Haplogroup F și subclasele sale - 2019-2020 ISOGG 2020
- ^ Y-SNP solicită Krems WA3 Genetiker. Revizuit în 2021
- ^ Teschler-Nicola, M., Fernandes, D., Händel, M. și colab. ADN-ul antic dezvăluie gemeni nou-născuți monozigoți din paleoliticul superior. Commun Biol 3, 650 (2020). https://doi.org/10.1038/s42003-020-01372-8
- ^ Kivisild T, Rootsi S, Metspalu M, și colab. Moștenirea genetică a primilor coloniști persistă atât în populațiile tribale indiene, cât și în cele de castă. Sunt J Hum Genet. (2003); 72 (2): 313-332. doi: 10.1086 / 346068
- ^ Karafet, T., Mendez, F., Sudoyo, H. și colab. Îmbunătățirea rezoluției filogenetice și diversificarea rapidă a haplogrupului K-M526 al cromozomului Y în Asia de Sud-Est. Eur J Hum Genet 23, 369-373 (2015). https://doi.org/10.1038/ejhg.2014.106
- ^ Underhill PA, Poznik GD, Rootsi S și colab. Structura filogenetică și geografică a haplogrupului R1a al cromozomului Y. Eur J Hum Genet. 2015; 23 (1): 124-131. doi: 10.1038 / ejhg.2014.50
- ^ (EN) Natalie M. Myres, Siiri Rootsi și Alice A. Lin, A major cromozom Y-haplogroup R1b Holocene era fundatory effect in Central and Western Europe , in the European Journal of Human Genetics, vol. 19, nr. 1, 1 ianuarie 2011, pp. 95-101, DOI : 10.1038 / ejhg.2010.146 . Adus la 18 decembrie 2015 .
- ^ A b (EN) Wolfgang Haak, Iosif Lazaridis și Nick Patterson, Migrația masivă din stepe a fost o sursă pentru limbile indo-europene în Europa , în Nature, vol. 522, nr. 7555, 11 iunie 2015, pp. 207-211, DOI : 10.1038 / nature14317 . Adus la 18 decembrie 2015 .
Bibliografie
- https://web.archive.org/web/20170116195823/http://ycc.biosci.arizona.edu/ Y Chromosome Consortium
- 2005 Arborele filogenetic al cromozomului Y , de la FamilyTreeDNA.com
- Un sistem de nomenclatură pentru Arborele Haplogrupurilor Y-cromozomiale umane , Genome.org
Elemente conexe
- Adam cromozomial
- Haplogrupuri mitocondriale umane
- Ipoteze genetice asupra populației Europei
- Genetica sardinilor
- Teoria catastrofei Toba
Alte proiecte
-
 Wikimedia Commons conține imagini sau alte fișiere pe grupuri Haplog ale cromozomului Y
Wikimedia Commons conține imagini sau alte fișiere pe grupuri Haplog ale cromozomului Y
linkuri externe
- Hărți Haplogroups din lume ( PDF ), pe scs.uiuc.edu . Adus la 30 aprilie 2007 (arhivat din original la 28 iulie 2004) .
- ISOGG Y-DNA Haplogroup Tree 2010 , pe isogg.org .
