Mutatie genetica
Prin mutație genetică înțelegem orice schimbare stabilă și ereditară în secvența nucleotidică a unui genom sau mai general a materialului genetic (atât ADN, cât și ARN ) datorită agenților externi sau întâmplării , dar nu recombinării genetice . [1] Prin urmare, o mutație modifică genotipul unui individ și poate modifica în cele din urmă fenotipul acestuia în funcție de caracteristicile și interacțiunile sale cu mediul.
Mutațiile sunt elementele de bază prin care pot avea loc procesele evolutive . De fapt, mutațiile determină așa-numita variabilitate genetică sau starea pentru care organismele diferă între ele pentru unul sau mai multe caractere . Selecția naturală operează pe această variabilitate, prin recombinare genetică , care promovează mutații favorabile în detrimentul celor nefavorabile sau chiar letale. Fiind o parte a mutațiilor nefavorabile, organismele au dezvoltat diferite mecanisme pentru repararea ADN-ului de la diferitele daune pe care le poate suferi, reducând astfel rata mutației.
Mutațiile se disting de genetici pe baza scării lor de acțiune: alterarea poate viza o singură genă , porțiuni ale genomului sau întregul set cromozomial.
Dacă mutațiile apar într-o celulă somatică, acestea, împreună cu efectele relative, vor fi prezente în toate celulele derivate din aceasta prin mitoză; unele dintre aceste mutații pot face celulele maligne și pot provoca cancer și sunt responsabile de unele malformații congenitale. Dacă mutațiile sunt prezente în celulele germinale sau în gameți, acestea sunt moștenite de generațiile următoare și pot duce în cele din urmă la boli genetice moștenite.
Originea mutațiilor
Mutațiile sunt în general clasificate în două clase în funcție de originea lor.
Mutații spontane
Mutațiile spontane sunt mutații cauzate de factori chimici endogeni și de erori în procesele care au loc pe materialul genetic; definiția mutației spontane este una care apare în absența mutagenilor cunoscuți. Nu sunt foarte frecvente, dar sunt încă inevitabile, având în vedere imperfecțiunea intrinsecă a fiecărui mecanism molecular. Erorile se pot datora:
- Tautomeria - o bază este modificată prin deplasarea unui atom de hidrogen .
- Deaminare - reacție care transformă o bază azotată într-una diferită; de exemplu provoacă tranziția C → U (care poate fi reparată ); există și dezaminarea spontană a 5-metilcitozinei în T și dezaminarea care determină A → HX (adenină → hipoxantină ).
- Purificare - hidroliza legăturii glicozidice și formarea unei nucleotide fără baze (de obicei G sau A).
- Deteriorare oxidativă - datorită formării spontane în celula speciilor cu atomi de oxigen foarte reactivi, capabili să atace ADN-ul și să provoace deteriorarea firului unic sau dublu și deteriorarea bazelor azotate.
- Erori în procesele de replicare , recombinare și reparare a ADN-ului . De exemplu, se poate datora ADN-polimerazei care adaugă nucleotide incorecte; aceasta poate genera o transversie dacă există schimbul de purină cu pirimidină sau invers; o tranziție dacă există schimbul unei purine cu o altă purină sau a unei pirimidine cu o altă pirimidină.
Mutații induse
Mutațiile induse sunt produse în schimb de acțiunea unor agenți fizici sau chimici anumiți numiți agenți mutageni . Mutageneza este procesul care determină o mutație indusă și mutagenizată organismul în care a fost produsă mutația. Daunele datorate mutațiilor induse se disting în:
- Înlocuirea bazelor cu molecule cu o structură similară cu cele prezente în mod obișnuit în ADN, dar care formează perechi diferite și, prin urmare, incorecte.
- Adăugarea grupelor substituente la bazele azotate : de asemenea, în acest caz, generează molecule cu o capacitate incorectă de împerechere.
- Deteriorarea bazelor azotate: ruperea legăturilor sau adăugarea de noi în comparație cu starea normală.
- Inserarea sau ștergerea bazei.
Mutagenii fizici sunt în principal radiații ionizante ( raze X , raze gamma ) și neionizante ( raze UV ); agenții chimici sunt foarte numeroși și aparțin diferitelor clase de compuși. În plus față de natura lor, mutagenii diferă și în spectrul mutațional, adică în tipul (sau tipurile) de mutație pe care îl pot provoca. De multe ori aceeași consecință poate fi cauzată de diferiți mutageni (chiar și de natură), chiar dacă în general mecanismele cu care au acționat sunt profund diferite.
O diferență importantă între mutageni fizici și chimici este că primii acționează independent de organism; mutagenii chimici, pe de altă parte, pot avea efecte diferite în funcție de sistemul biologic. În timp ce o radiație, de fapt, afectează în mod direct materialul genetic, un compus chimic poate interacționa cu alte molecule (enzime, metaboliți, specii reactive ...) prezente în celulă, care îi pot varia caracteristicile.
Mutații genetice
Acestea sunt mutațiile care modifică o singură genă și, prin urmare, „cea mai mică” care poate avea. Ca atare, acestea nu sunt vizibile prin analize microscopice (cu excepția unor cazuri extreme), dar pot fi găsite numai prin analize genetice. Mutațiile genice duc la formarea de noi forme genice sau alele noi, numite alele mutante . Ca atare, acestea sunt rare în populație și diferă de alelele mai frecvente numite tipuri sălbatice . Cu toate acestea, trebuie făcută o distincție între alele mutante și alele morfice . Morfii sunt de fapt două sau mai multe alele ale aceleiași gene cu o frecvență mai mare de 1% ( polimorfism ). În lumina acestui fapt, rezultă că conceptul de mutație nu este absolut: o genă poate suferi o mutație; totuși, dacă alela mutantă găsește condițiile de răspândire în populație și depășește frecvența de 1%, nu vom mai vorbi de mutație ci de morfo.
Acestea pot fi împărțite în trei categorii: mutații punctuale, mutații dinamice și rearanjamente structurale ale genelor.
Mutații punctuale
O mutație punctuală este o variație a secvenței ADN care afectează una sau câteva nucleotide, dar este, de asemenea, posibil să se ia în considerare mutațiile „punctuale” de până la 50 nucleotide. Este posibil ca multe mutații punctuale să nu aibă niciun efect, caz în care se spune că sunt neutre, de fapt o mare parte din ADN-ul dintr-un genom eucariot nu codifică produsele proteice și nu este sigur dacă schimbarea unei singure baze nucleotidice în acest silențios o parte din ADN ar putea afecta sănătatea unui organism. Cu toate acestea, o mutație cu un singur punct poate avea un impact semnificativ asupra fenotipului, așa cum se întâmplă, de exemplu, în anemia falciformă.
Înlocuirea bazelor
Mutațiile de substituție a bazei duc la schimbul unei nucleotide cu alta. Tranzițiile sunt definite atunci când există un schimb de purină cu o altă purină (A> G) sau a unei pirimidine cu o altă pirimidină (C> T); pe de altă parte, se spun transversii când schimbul este de purină cu pirimidină sau invers (C / T> A / G). Tranzițiile sunt în general mai frecvente decât transversiile.
Mutațiile punctuale pot fi de șase tipuri: silențioase, fără sens, ștergeri sau inserții de cadre, inserții fără sens, mutații de schimbare a cadrelor sau mutații de îmbinare.
- Mutațiile silențioase sau sinonime apar atunci când substituirea unei baze azotate într-o secvență ADN nu provoacă o modificare a secvenței de aminoacizi a proteinei în cauză. De exemplu, dacă tripletul TTT se schimbă în TTC, va exista o tranziție (T> C) în a treia poziție a tripletului, dar aminoacidul codificat începând de la tripletul de ARNm corespunzător (UUC) va fi întotdeauna fenilalanină datorită redundanță a codului nostru genetic care a degenerat. Mutațiile silențioase sunt în cea mai mare parte neutre, deoarece aminoacidul nu se modifică și, în consecință, funcționalitatea proteinei codificate în cadrul căreia se găsește tripletul mutat nu se schimbă. [2] Multe dintre mutațiile responsabile pentru un proces de splicing modificat apar în secvențele scurte ESE ( Exon Splicing Enhancer ) ale unor exoni, care sunt esențiale pentru îmbinarea corectă, deoarece anumite proteine implicate în reglarea acestui proces se leagă de ele. Când apar mutații în aceste secvențe, poate apărea includerea intronului în ARNm matur, care, dacă ar fi codificat, ar duce la proteine anormale. Mutațiile silențioase la secvențele ESS (Exonic Splicing Silencer) implicate, de asemenea, în mecanismul de splicing al transcriptului primar, pot duce în schimb la excluderea unui exon din ARNm matur și, în consecință, la codificarea proteinelor trunchiate de către ribozomi.
- Mutațiile Missense apar atunci când o bază azotată este substituită într-o secvență ADN, astfel încât secvența de aminoacizi este modificată. Dacă, de exemplu, tripletul TTT se transformă în TCT, cu o tranziție a bazei în a doua poziție (T> C), aminoacidul codificat nu va mai fi fenilalanină, ci serină. Acest tip de mutație poate fi neutru și nu determină niciun fenotip specific reprezentând pur și simplu un polimorfism nucleotidic unic (SNP) sau o variantă privată, dar poate da naștere și unor boli grave, cum ar fi anemia falciformă. În general, o mutație fără sens poate fi considerată neutră dacă aminoacidul substituit este prezent fără a prezenta un fenotip patologic la un anumit număr de indivizi sub forma unui polimorfism nucleotidic unic sau a unei variante private sau dacă aminoacidul codificat are proprietăți similare cel original. (de exemplu, înlocuirea acidului glutamic cu acid aspartic). Cu toate acestea, mutația poate da naștere unor condiții patologice atunci când aminoacidul codificat de noul triplet are proprietăți foarte diferite de precedentul (de exemplu înlocuirea unei valine cu acid aspartic), dacă nu a fost găsit în cazuri anterioare sau în contextul parental sau când apare într-o regiune foarte conservată a unei proteine. Adesea, chiar și o singură mutație într-o regiune extrem de conservată a unei proteine determină pierderea funcționalității acesteia.
- Ștergerile în cadre și respectiv inserțiile în cadre determină eliminarea unui triplet sau a unui număr de nucleotide divizibil cu 3 sau inserarea unui triplet sau a unui număr de nucleotide divizibil cu 3. Ele sunt „în cadre” deoarece nu sunt ele mută cadru de citire la nivel ribozomal, aceasta ar implica de fapt schimbarea aproape totală a secvenței de aminoacizi a unei proteine. Acest tip de mutație are ca rezultat eliminarea sau adăugarea de aminoacizi în proteina codificată de mARN-ul matur care le conține. Consecințele acestor mutații sunt foarte variate.
- Mutațiile fără sens apar atunci când o mutație nucleotidică a unui triplet are ca rezultat transformarea unui codon de aminoacizi într-un codon stop. De exemplu, tripletul AGC care codifică serina este înlocuit cu ATC, care va fi transcris în ARNm ca UAG, unul dintre cei trei codoni de oprire. Consecința este că proteina codificată nu este exportată sau, dacă este codificată, este trunchiată, deoarece traducerea se termină la codonul stop, ignorând tripletele sale din aval. Consecința acestei mutații este o proteină trunchiată nefuncțională sau nocivă. Cu toate acestea, dacă codonul de oprire este de cel puțin 50 de nucleotide din cea mai apropiată secvență de îmbinare în ARNm, celula activează un mecanism de protecție cunoscut sub numele de Nonsense Mediated Decay (NMD) care degradează ARNm mutant. Alternativ, poate fi activat un alt mecanism cunoscut sub denumirea de Nonsense-Alterated Splicing (NAS) care exclude exonul care conține tripletul mutat în codonul stop, permițând celorlalți exoni să fie asociați într-o proteină mai scurtă.
- Mutațiile de schimbare a cadrelor se datorează ștergerii sau inserțiilor unui număr de nucleotide care nu sunt divizibile cu 3, aceasta implică deplasarea cadrului de citire în aval de mutație și, prin urmare, codificarea unei secvențe de aminoacizi care nu corespunde cu cea a originalului transcriere. [3] Consecința este producerea de proteine anormale care au doar porțiuni din secvența corespunzătoare originalului sau non-exportul sau traducerea ARNm mutant.

- Mutațiile splice sunt un set de patru tipuri de mutații care implică secvențe importante pentru splicarea pre-ARNm. Un prim tip implică situl donator de îmbinare (GT) sau situl acceptor (de obicei AG). Mutațiile din acești doi markeri inițiali și finali ai unei secvențe de introni pot duce la includerea intronului în transcrierea matură sau la îmbinarea incorectă. O a doua tipologie implică scurte secvențe consens în amonte și în aval de situsul donator și site-ul acceptor, sau o secvență consens a sitului ramificator . O a treia tipologie implică mutații într-o secvență ESE sau ESS și poate fi atribuită și mutațiilor silențioase. În cele din urmă, o ultimă tipologie implică mutații care creează noi secvențe de consens în cadrul unui intron și, în acest caz, acesta sau părți din acesta pot fi incluse în transcriere sau într-un exon, caz în care apare saltarea exonului .
Mutații în regiunile de reglare a transcripției
Mutațiile punctiforme pot apărea și în regiunea reglatoare a unei gene. Acest lucru poate duce la consecințe foarte variabile, variind de la niciun efect fenotipic la modificări în expresia genelor care dau naștere unor patologii grave.
Studiile genetice au arătat că entropia unei surse genetice care crește în timp măsoară acumularea mutațiilor degenerative în ADN, progresivă în generațiile de celule reproduse de organism, precum și în generațiile de indivizi [4] .
Chiar dacă mutațiile aleatorii sau induse de om sunt clasificate ca favorabile (pe un orizont pe termen scurt, împotriva unei boli), ele sunt în general asociate cu o pierdere de informații a genei și o reducere a funcționalității acesteia în procesele organice obișnuite. [4 ] .
Mutații dinamice
Mutațiile dinamice se datorează repetării tripletelor de nucleotide într-o regiune de codificare (în acest caz, cel mai frecvent triplet este CAG care codifică glutamina) sau necodificării unei gene. Mutația, care apare în cursul replicării ADN-ului , determină o variație a numărului acestor secvențe repetate; noua catena de ADN poate avea un exces sau un defect. Fenomenul care provoacă mutația se numește derapaj de replicare și se datorează împerecherii proaste a celor două fire complementare. Bolile genetice asociate cu acest tip de mutație sunt coreea Huntington și sindromul X fragil .
Rearanjări structurale ale genelor
Rearanjările structurale ale genelor includ toate acele mutații care modifică genomul variind structura cromozomilor ( mutații cromozomiale ) sau numărul de cromozomi ( mutații genomice ). Sunt definite și anomalii citogenetice sau anomalii cariotipice . Aceste modificări sunt de obicei o consecință a unei erori în diviziunea celulară , meioză sau mitoză . Spre deosebire de mutațiile genetice care pot fi găsite doar prin analiza genetică, acestea pot fi, în multe cazuri, vizibile și la microscop, deoarece conduc la formarea unor structuri cromozomiale particulare în faza de împerechere. Consecințele lor pot varia de la niciun efect fenotipic atunci când mutațiile implică secvențe repetate la patologii genetice severe.

Mutații cromozomiale
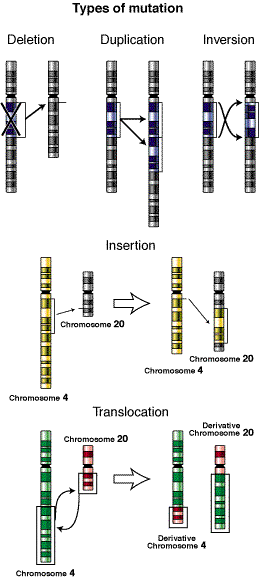
Vorbim despre mutații cromozomiale sau anomalii cromozomiale atunci când structura unuia sau mai multor cromozomi este modificată. Mutațiile cromozomiale pot fi de șase tipuri: deleții sau duplicări, inversiuni, translocații, conversii genetice, transpuneri și cromozomi inelari.
- Ștergerile și duplicările se datorează erorilor în procesul de recombinare omoloagă, numită și încrucișare, care apare în meioză. Datorită prezenței genelor care au un grad ridicat de omologie, a pseudogenelor sau a secvențelor repetate, pot apărea erori în asocierea cromozomilor, astfel încât fragmentele de ADN schimbate între cei doi cromozomi nu sunt egale, astfel încât o deleție pe unul și duplicarea pe de altă parte. Se poate întâmpla ca în timpul unei recombinări neomoloage datorită unei rearanjări incorecte, unele gene din blocurile de ADN să fie plasate într-o zonă cu o prezență heterocromatică puternică. În acest caz, este posibil ca aceste gene să fie inactivate de fenomenul efectului de poziție. Tulburările asociate cu această anomalie sunt sindromul Wolf-Hirschhorn , care este cauzat de pierderea unei părți a brațului scurt al cromozomului 4 și sindromul Jacobsen , care provine din ștergerea capătului cromozomului 11 . Unele tulburări cunoscute datorate duplicării sunt sindromul Bloom și sindromul Rett .
- Translocarea are loc atunci când o regiune a unui cromozom este transferată într-o altă locație pe același sau pe alt cromozom; există două tipuri principale de translocări: translocarea reciprocă și translocarea robertsoniană.
- Inversarea este o mutație datorată inversării orientării unei regiuni a unui cromozom care provoacă o inversare a ordinii genelor. Acestea se datorează prezenței puternice a secvențelor duplicate sau inversate în gena afectată. Omologia celor două secvențe determină plierea ADN-ului și asocierea lor. Celula intervine efectuând o recombinare neomologă care determină inversarea regiunii între cele două repetări.
- Conversia genelor este o mutație în care există transferuri nereciproce de secvențe de ADN între gene sau alele, în primul caz conversia este interalelică în al doilea caz se spune că este interlocus. Dintre cele două secvențe, cea care rămâne neschimbată se numește donator, cea care este modificată se numește acceptor.
- Transpunerea are loc atunci când un element transpozabil precum LINE sau SINE se integrează în genom după ce a fost retranscris. Această mutație poate să nu aibă niciun efect fenotipic dacă afectează regiunile repetate, dar poate da naștere unor patologii atunci când transpunerea are loc într-o genă transcrisă activ.
- Inelul apare atunci când cele două capete ale unui cromozom se împerechează între ele, formând un inel. Această anomalie poate implica sau nu pierderea materialului genetic.
Mutații ale cariotipului
Vorbim de mutație genomică sau anomalii cariotipice atunci când un organism are mai mulți sau mai puțini cromozomi decât în mod normal.
Dacă sunt prezente seturi întregi de cromozomi, mai mult sau mai puțin, vorbim de euploidie aberantă ; dacă, pe de altă parte, este doar o parte a trusei în exces sau în defect, anomalia se numește aneuploidie .
La om și, în general, la toate organismele diploide , care au deci perechi de cromozomi omologi, cele mai frecvente forme de aneuploidie sunt lipsa unui cromozom dintr-o pereche ( monozomie ) sau prezența unui cromozom suplimentar într-o pereche ( trisomie ). Mai rar este cazul pierderii unui întreg cuplu ( nulisomie ).
Un exemplu al efectelor unei astfel de anomalii este sindromul Down , numit și trisomia 21; indivizii cu acest sindrom au trei copii ale cromozomului 21 în loc de două. Sindromul Turner, pe de altă parte, este un exemplu de monozomie; indivizii născuți cu această anomalie au un singur cromozom sexual, femela X. Dintre organismele haploide, cele mai frecvente cazuri de aneuploidie constau în prezența unui cromozom supranumerar (disomie).
Efectele mutațiilor la scară largă
De asemenea, pentru această categorie de mutații, posibilele consecințe asupra organismului sunt variabile. În general, va exista un efect ori de câte ori, în modificarea cromozomului sau genomului , se modifică și secvența sau numărul uneia sau mai multor gene. Spre deosebire de mutațiile genetice, în acest caz efectele vor fi întotdeauna negative.
- Pentru toate mutațiile cromozomiale este necesar să se rupă catena dublă în cel puțin un punct pentru a permite rearanjarea ulterioară: dacă pauza se produce într-o genă la sfârșitul procesului, secvența sa va fi mutată. De exemplu, într-o inversiune , dacă fracturile au avut loc în secvențe de codificare, ca urmare a orientării diferite a fragmentului reintrodus, genele de la capete vor avea o parte din secvența dreaptă și o parte provenind de la celălalt capăt al fragmentului, prin urmare greșit (genele din interiorul fragmentului, pe de altă parte, nu vor fi schimbate, ci doar inversate în ordine). Situația este similară pentru translocații . Ștergerile și duplicările, pe de altă parte, vor avea consecințe suplimentare, fiind rearanjări care nu modifică aranjamentul, ci cantitatea de material genetic. Ștergerea va avea efecte negative proporționale cu dimensiunea fragmentului șters. Duplicarea crește numărul de copii ale genelor conținute în fragmentul duplicat: totuși, și acesta are consecințe dăunătoare, deoarece determină un dezechilibru genetic.
- În mod similar, în mutațiile cariotipului există o creștere sau scădere a dimensiunii genomului celular. Auploidia aberantă este rară, dar totuși letală la animale (cu rare excepții), poate fi decisivă și la plante . Cu toate acestea, studii recente au arătat acum că aneuploidia este una dintre cauzele directe ale multor tumori (și nu o consecință așa cum s-a crezut și ea) [5] .
Alte mutații
- Mutații din sistemele de reparații : în mod paradoxal, mutațiile genetice pot fi, de asemenea, inserate prin anumite procese de reparare a ADN-ului . De fapt, se poate întâmpla ca anumite daune ale ADN-ului să nu fie recunoscute și reparate de către nici un utilaj responsabil pentru această sarcină, până la următorul ciclu de replicare: dacă aceste daune (cum ar fi fotoprodusele induse de radiațiile ultraviolete ) blochează acțiunea ADN polimerazei , adică, previn replicarea ADN în aval de daune, provocând pierderea materialului genetic cu consecințe practic întotdeauna letale pentru celula fiică. S-au dezvoltat așa-numitele mecanisme de reparare SOS , care acționează în aceste cazuri extreme: polimerazele acestui sistem nu se blochează, ci adaugă aceleași nucleotide în fața deteriorării; în majoritatea cazurilor, însă, adăugarea este aleatorie și, prin urmare, are un risc ridicat de a adăuga nucleotide incorecte; de aceea să provoace mutații. Un alt sistem cu consecințe similare este sistemul de reparare a ADN-ului cu două fire, care nu exploatează omologia: așa-numita NHEJ ( unire finală neomologă ). Și aici, pentru a repara pauza și a evita pierderea fragmentului fără centromere în ciclul meiotic sau mitotic ulterior, sistemul provoacă ștergeri ale secvențelor adiacente pauzei. Prin urmare, în ambele cazuri, sistemele evită o deteriorare foarte mare, dar trebuie să plătească prețul pentru includerea mutațiilor care sunt, de asemenea, potențial dăunătoare.
- Mutații condiționale : sunt mutații care, deși prezente, au efect doar în anumite condiții de mediu. Cele mai frecvente cazuri, printre haploizi, sunt mutațiile sensibile la temperatură ; adică acționează numai peste (sau sub) anumite praguri de temperatură.
- Mutații prin transpunere : se datorează inserării, în secvența de codificare sau reglare, a elementelor transpozabile sau a transpozonilor . Acestea au ca rezultat inactivarea completă a genei, dar fiind elemente dinamice, pot scăpa de genă și restabili secvența corectă a acesteia.
Efectele mutațiilor genetice
Efectele pot fi semnificativ diferite în funcție de tipul mutației și de locația în care apare. O mutație nu poate duce la nicio consecință și atunci când se referă la ADN care nu codifică (sau mai degrabă pare că nu codifică) niciun produs genetic (așa-numitul ADN junk sau ADN junk ). Dacă mutația schimbă în schimb secvențele de codare sau genele , există o variație a tipului sau cantității produsului genetic corespunzător, care poate fi o proteină funcțională sau ARN ( ARNr , ARNt , snARN, etc.). În acest caz vorbim de mutație biochimică ; dacă mutația biochimică duce la o schimbare vizibilă a fenotipului se numește mutație morfologică .
De asemenea, distingem, din nou în raport cu efectele, în:
- mutație pozitivă : aduce un avantaj evolutiv;
- mutație neutră : nu are ca rezultat o slăbire a capacității de reproducere a individului;
- mutazione disvitale o semiletale : rende più difficoltosa la perpetuazione riproduttiva dell'individuo (il tipico esempio sono le malattie genetiche che debilitano in qualche modo l'individuo, rendendolo meno capace di riprodursi, senza però impedirglielo totalmente);
- mutazione subletale : non permette all'individuo di raggiungere l'età riproduttiva;
- mutazione letale : porta alla morte dell'individuo in fase embrionale o fetale.
L'efficacia della mutazione, sia positiva che negativa, dipende poi dal tipo di allele mutato così creato; questo potrà essere infatti dominante o recessivo . Nei diploidi se è dominante avrà sempre effetto (sia in un eterozigote che in un omozigote dominante); se è recessivo, essendo aploinsufficiente, per avere effetto ha bisogno che anche l'altro elemento della coppia genica sia mutato (individuo omozigote recessivo). Negli aploidi , che sono emizigoti , la mutazione avrà invece sempre effetto.
Le mutazioni possono essere in alcuni casi pleiotropiche , ovvero possono dar luogo a più effetti: ad esempio nel topo ( Mus musculus ), un comune allele mutante e dominante in condizioni di eterozigosi determina una variazione del colore del mantello; in omozigosi, cioè quando l'allele mutato è presente in duplice copia, provoca invece la morte dell'animale prima ancora della nascita. Si può presumere quindi che il gene mutato controlli non solo il colore della pelliccia, ma anche qualche altro processo biochimico vitale per l'organismo.
Reversione e soppressione
A differenza di mutazioni su larga scala, quelle puntiformi possono essere soggette a reversione: attraverso altre mutazioni le prime possono scomparire o ne può scomparire l'effetto sull'organismo. Nel primo caso parliamo di reversione in senso stretto: la mutazione revertente può riportare il codone mutato così com'era originariamente (si parla comunemente di retromutazione ); oppure la mutazione può alterare sempre il codone mutato trasformandolo in uno diverso da quello iniziale, ma codificante lo stesso amminoacido ( reversione di sito ). Nel caso in cui la seconda mutazione occorra su un codone diverso si parla di soppressione : la soppressione potrà essere interna se il codone è all'interno del gene mutato o esterna se appartiene ad un altro gene. Un esempio di soppressione interna è una delezione (o un'inserzione) che annulla l'effetto di una inserzione (o delezione) nello stesso gene. Il caso più comune di soppressione esterna è invece la mutazione nell'anticodone di un tRNA che annulla quella avvenuta nel codone complementare.
Nomenclatura
È stata sviluppata una particolare nomenclatura per specificare il tipo di mutazione e il tipo di base o amminoacido cambiato.
- Sostituzione di un amminoacido - (ad esempio D111E) La prima lettera rappresenta il codice (ad una lettera) dell'amminoacido originariamente presente; il numero indica la posizione dell'amminoacido a partire dall'estremità N-terminale; la seconda lettera è il codice dell'amminoacido sostituito in seguito alla mutazione. Se la seconda lettera è una X vuol dire che un qualunque amminoacido può sostituire quello iniziale.
- Delezione di un amminoacido - (ad esempio ΔF508) Il simbolo greco Δ ( delta ) indica una delezione; la lettera rappresenta l'amminoacido deleto; il numero è la posizione, sempre dall'N-terminale, dove si trovava l'amminoacido nella sequenza prima della delezione.
Storia

Il primo a introdurre il termine mutazione nel campo della genetica fu Hugo de Vries [6] , nel 1901 , che lo riferiva però alle brusche variazioni nei caratteri di un organismo; in particolare osservando come nella progenie di un ceppo della pianta Oenothera lamarckiana si potevano ottenere alcuni individui inaspettatamente giganti. Il concetto di mutazione così come è inteso oggi, invece, fu usato solo a partire dal 1927 . In generale si può dire, comunque, che le mutazioni genetiche hanno avuto un ruolo essenziale ancora prima, fin dagli albori della genetica; già nei celebri lavori del padre della genetica, Gregor Mendel , infatti, i fenotipi come il colore bianco dei petali o giallo dei semi maturi, usati per formulare le sue leggi, non erano che dovute a mutazioni inattivanti dei corrispettivi geni.

Il primo "sfruttamento" consapevole delle mutazioni avviene a partire dagli studi, condotti ai primi del 900 da Thomas Hunt Morgan e il suo cosiddetto fly group , sul moscerino della frutta Drosophila melanogaster . Morgan e colleghi portarono le prime importanti prove sperimentali della teoria cromosomica dell'ereditarietà , che ipotizzava per la prima volta una stretta connessione tra geni e cromosomi. I ricercatori isolarono in una vasta popolazione di insetti un moscerino dagli occhi bianchi (mentre nel fenotipo selvatico erano rossi). Anche qui il fenotipo particolare era stato provocato da una mutazione spontanea nel gene per il colore degli occhi.
Mutazione che aveva prodotto una nuova forma allelica ; gli incroci tra individui con alleli diversi hanno permesso di ottenere i risultati sopra detti. Morgan isolò per questi incroci, dopo il caso del moscerino dagli occhi bianchi, ben 83 ceppi ciascuno con mutazioni su geni diversi. Le mutazioni ebbero poi un ruolo sempre più crescente da quando furono scoperti i primi agenti mutageni . La maggior parte degli esperimenti chiave nella storia della genetica fecero uso di mutazioni indotte : nel 1941 , nel loro celebre esperimento che portò al dogma un gene-un enzima , Edward Lawrie Tatum e George Wells Beadle fecero ad esempio uso di ceppi di Neurospora crassa mutagenizzati tramite raggi X . In modo analogo Tatum e Joshua Lederberg nel 1946 usarono mutazioni in ceppi di Escherichia coli per dimostrare l'esistenza del processo di coniugazione batterica .
Un importante capitolo nella storia delle mutazioni nella genetica riguarda la disputa sull'origine delle mutazioni nei batteri . Intorno agli anni quaranta infatti alcuni batteriologi misero in dubbio che le mutazioni potessero avvenire nei batteri in modo del tutto spontaneo (come era invece accettato per gli organismi superiori) essi ritenevano piuttosto che le mutazioni erano indotte dalla presenza di particolari condizioni ambientali. Ad esempio, i batteri che sopravvivevano in seguito all'aggiunta di un batteriofago avevano acquisito la resistenza grazie a una mutazione indotta dalla stessa presenza dei fagi (teoria adattativa). Numerosi altri studiosi invece erano convinti che le mutazioni si verificassero così come in tutti gli altri organismi, spontaneamente. Quest'ultima teoria (teoria genetica) fu definitivamente dimostrata da due celebri esperimenti: il cosiddetto test di fluttuazione (o di Salvador Luria e Max Delbrück ), sviluppato nel 1943 e la tecnica della piastratura delle repliche ideata da Joshua e Esther Lederberg .
Applicazioni nelle analisi genetiche
Gli studi genetici che fanno uso di mutazioni possono essere distinte in due categorie a seconda dello scopo dello studio e dei dati che si posseggono: studi di genetica diretta e di genetica indiretta . Il primo approccio è usato qualora si voglia determinare i geni che in un organismo siamo correlati a una certa funzione: in questo caso l'organismo viene esposto a mutageni e successivamente il genetista compie la cosiddetta "caccia al mutante", in cui va a ricercare gli individui i cui sono stati alterati i fenotipi correlati alla funzione che si sta studiando. A questo punto si determina la posizione del gene mutato tramite incroci si isola e si analizza in dettaglio: ne si determina la sequenza nucleotidica e si osserva per quale prodotto genico codifica. Nella genetica diretta quindi si parte dal fenotipo per vedere da quale genotipo è causato. Il secondo tipo di studio compie invece il percorso inverso: parte dal genotipo per studiare il fenotipo: si parte in genere da una sequenza di DNA o RNA nota o addirittura da un prodotto genico (di solito una proteina ), si mutagenizzano in modo selettivo e si vede che effetti fenotipo causano nell'organismo; si parla in questo caso anche di silenziamento genico .
Mutagenesi sito specifica
Molto importanti sono le tecniche che permettono di ottenere mutazioni sito specifiche; mutazioni cioè indotte in modo selettivo nelle zone di interesse di una sequenza. In questo modo per esempio è possibile inserire una mutazione in un particolare dominio di una proteina e, saggiando le conseguenze, determinarne la funzione.
Test di mutagenesi
I test di mutagenesi sono procedure in cui cellule, tessuti o interi organismi sono esposti all'azione di una sostanza chimica, per verificarne e/o quantificarne la mutagenicità; i sistemi biologici in esame sono quindi studiati, dopo un certo periodo di incubazione, e analizzati per vedere la presenza di eventuali mutazioni. In generale la capacità mutagena di un agente è direttamente proporzionale ai mutanti identificati al termine del test. I test routinari sono svolti su batteri , essendo sistemi più conosciuti e di più facile utilizzo. I test sono però volti a scoprire il danno che una sostanza può creare all'uomo, il quale ha, ovviamente, molte caratteristiche biologiche diverse dai procarioti ; per questo si procede a modificare geneticamente i batteri usati nei test per mimare un sistema il più vicino possibile a quello umano, oppure si usano cellule di mammifero (solitamente di roditori). Tra i test più usati ci sono il test di Ames e il test del micronucleo .
Il test di Ames

Un esempio di applicazione delle mutazioni in campo biomedico è il test di Ames . Il test, sviluppato negli anni settanta da Bruce Ames , ha lo scopo di determinare la cancerogenicità di una sostanza studiando la sua capacità di indurre mutazioni; in generale infatti una sostanza mutagena è anche cancerogena. È solitamente usata una forma mutata del batterio Salmonella typhimurium , ad esempio un ceppo che non è in grado di crescere in terreno privo di istidina ; il ceppo è diviso in due piastre separate con terreni privi dell' amminoacido : uno di essi sarà esposto alla sostanza da testare l'altro no. Se la sostanza ha capacità mutagena ci sarà una certa probabilità che induca delle reversioni della mutazione; annulla cioè l'effetto della prima mutazione con una mutazione, permettendo di nuovo al batterio di sopravvivere anche in assenza di istidina. Sul ceppo non mutagenizzato invece non ci sarà nessuna colonia o molto poche (essendo la reversione per mutazione spontanea molto rara). Più colonie sopravviveranno nel campione mutagenizzato, maggiore sarà stato il numero di retromutazioni e quindi maggiore è la cancerogenicità della sostanza.
Esempi di mutazioni positive
- La tolleranza al lattosio , che permette la digeribilità del latte e degli alimenti che lo contengono, è derivata secondo i genetisti da una mutazione favorevole avvenuta circa 10.000 anni fa (8.000 secondo altre fonti) che colpì gli uomini che abitavano la zona del Caucaso . È un chiaro esempio di mutazione favorevole che, in quanto tale, presto si diffuse rapidamente nella popolazione: ad oggi solo una parte della popolazione umana soffre di intolleranza per questa sostanza. Ulteriori dimostrazioni derivano dal fatto che popoli che abitarono zone lontane dall'origine della mutazione, come Asiatici e Africani, e che non vennero in stretto contatto con i caucasici, presentano oggi una maggiore diffusione dell' intolleranza al lattosio congenita.
- Un altro caso che si ritiene essere una mutazione positiva è la delezione di 32 coppie di basi nel gene umano CCR5 (CCR5-32) che conferisce all'uomo la resistenza all' AIDS negli omozigoti e ritarda gli effetti negli eterozigoti . [7] La mutazione è mediamente più diffusa tra coloro che hanno discendenza europea; una teoria per spiegare la maggiore diffusione nella popolazione europea della mutazione CCR-32 la mette in relazione con le forme di resistenza alla peste bubbonica sviluppate nella metà del quattordicesimo secolo. [8]
- La mutazione dell' apolipoproteina Apo A-1 in Apo A-1 Milano , tale mutazione conferisce agli abitanti di Limone sul Garda (portatori di questa mutazione) un'innata resistenza agli effetti dannosi del " colesterolo cattivo " , dei trigliceridi elevati nel sangue e previene la formazione delle placche ateromasiche [9] ; Questa proteina mutata ha conferito, inoltre, agli abitanti del paese un'estrema longevità, una dozzina di residenti ha superato i 100 anni (su circa un migliaio di abitanti).
Esempi di mutazioni negative
- La fenilchetonuria è una malattia provocata da una mutazione genica che rallenta o blocca la capacità di trasformare l' amminoacido fenilalanina in tirosina . Questo dunque si accumula nell'organismo e se in grado di raggiungere il cervello può provocare danni neurologici .
 Un esemplare di gatto Man
Un esemplare di gatto Man

- Il daltonismo ha tra le varie cause possibili quelle genetiche, dovute a mutazioni su geni che codificano fotorecettori .
- L' albinismo è una disfunzione genetica dovuta alla mutazione del gene per la melanina .
- L' anemia drepanocitica o anemia falciforme è una malattia del sangue conseguenza di una mutazione che provoca l'alterazione della struttura e della funzione dei globuli rossi .
- Il gatto Man si è sviluppato a seguito di un'alta frequenza di accoppiamento tra individui consanguinei. La mutazione riguarda il gene cosiddetto "M" e provoca oltre all'assenza di coda anomalie nella struttura scheletrica. Il gene è dominante ma si manifesta con diversa espressività . Gli individui omozigoti dominanti (M/M) non sopravvivono e muoiono quando sono ancora nello stato di feto nell' utero materno.
Mutazioni per scopi commerciali
Mutazioni indotte possono essere alla base di processi per la selezione di organismi mutanti con caratteristiche vantaggiose. Sono pratiche usate principalmente in agricoltura e rivolte a specie vegetali . I vantaggi possono riguardare ad esempio la capacità di crescere in particolari condizioni ambientali, la presenza di frutti più grandi o privi di semi ecc. In molti casi le mutazioni riguardano la variazione nel numero di cromosomi. Esempi sono:
- la produzione di specie con un corredo cromosomico in più del normale e dispari ( aneuploidia ); le banane che troviamo in commercio, ad esempio, sono triploidi invece di diploidi . Lo scopo è di ottenere piante che siano sterili e per questo con frutti privi di semi.
- il raddoppio del corredo cromosomico ( euploidia aberrante ): ad esempio in molte specie di uva , che è solitamente diploide, si agisce bloccando il processo meiotico generando piante tetraploidi (con 4 corredi). In questo caso la conseguenza favorevole è l'aumento delle dimensioni del frutto (l'acino d'uva) in parallelo con l'aumento del materiale genetico.
Queste metodologie non devono essere confuse con quelle usate in ingegneria genetica e che sono alla base degli organismi geneticamente modificati ( OGM ).
Note
- ^ Anche se formalmente le mutazioni riguardano entrambi gli acidi nucleici, esse interessano quasi sempre il DNA. Tra i rari casi di mutazioni su RNA rientrano, ad esempio, quelle che colpiscono gli RNA-virus
- ^ In pratica, tuttavia, molte mutazioni silenti, nonostante non alterino il tipo di amminoacido codificato in virtù della degenerazione del codice a triplette, possono influenzare il processo di splicing del trascritto primario, portando a splicing alternativi che possono generare RNA messaggeri maturi più corti, che portano di conseguenza a proteine più corte. È il caso ad esempio della sindrome HGPS (sindrome della progeria di Hutchinson-Gilford), in cui la mutazione sinonima di un singolo codone genera mRNA maturi e proteine più corte, responsabili di fenotipi alterati.
- ^ Nel caso i nucleotidi aggiunti siano identici a quelli adiacenti è formalmente più corretto parlare di duplicazione invece che di inserzione.
- ^ a b
 La genetica esclude l'evoluzionismo: evidenti perdite di informazioni , su laverascienza . URL consultato il 28 luglio 2018 ( archiviato il 28 luglio 2018) . , in particolare dal minuto 8.08' alla fine, ai primi 2 minuti, e commenti
La genetica esclude l'evoluzionismo: evidenti perdite di informazioni , su laverascienza . URL consultato il 28 luglio 2018 ( archiviato il 28 luglio 2018) . , in particolare dal minuto 8.08' alla fine, ai primi 2 minuti, e commenti - ^ Se i cromosomi danno i numeri, Le Scienze 462 pag.32,2007
- ^ Hugo de Vries sull'ereditarietà
- ^ Il gene CCR5 e l'infezione da HIV, Antonio Pacheco. Archiviato il 16 aprile 2007 in Internet Archive .
- ^ PBS: I misteri della morte nera.
- ^ Franceschini G, Sirtori CR, Capurso A, Weisgraber KH, Mahley RW, AI Milano apoprotein. Decreased high density lipoprotein cholesterol levels with significant lipoprotein modifications and without clinical atherosclerosis in an Italian family , in J. Clin. Invest. , vol. 66, 1980, p. 892–900, DOI : 10.1172/JCI109956 .
Bibliografia
- Lucia Migliore, Mutagenesi ambientale , Bologna, Zanichelli, 2004. ISBN 88-08-07719-5
- Peter J. Russel, Genetica , Napoli, Edises, 2002. ISBN 88-7959-284-X
- Anthony FJ Griffiths, Genetica. Principi di analisi formale. , Zanichelli, 2006
- ( EN ) Leroi A. 2003. Mutants: On the form, varieties & errors of the human body . 1:16-17. Harper Collins 2003
- ( EN ) Maki H. 2002. Origins of spontaneous mutations: specificity and directionality of base-substitution, frameshift, and sequence-substitution mutageneses . Annual Review of Genetics 36:279-303.
- ( EN ) Taggart R. Starr C. Biology The Unity and Diversity of Life: Mutated Genes and Their Protein Products . 4.4:227. Thompson Brooks/Cole 2006.
Libri online
- ( EN ) Capitolo 7, The Molecular Basis of Mutation ida Modern Genetic Analysis , Anthony JF Griffiths, William M. Gelbart, Jeffrey H. Miller e Richard C. Lewontin (1999) pubblicato da WH Freeman and Company ISBN 0-7167-3597-0 .
Voci correlate
Altri progetti
-
 Wikizionario contiene il lemma di dizionario « mutazione »
Wikizionario contiene il lemma di dizionario « mutazione » -
 Wikimedia Commons contiene immagini o altri file su mutazione
Wikimedia Commons contiene immagini o altri file su mutazione
Collegamenti esterni
- Mutazione genetica , su Treccani.it – Enciclopedie on line , Istituto dell'Enciclopedia Italiana .
- ( EN )Mutazione genetica / Mutazione genetica (altra versione) , su Enciclopedia Britannica , Encyclopædia Britannica, Inc.
- ( EN ) Anomalie cromosomiche , su genome.gov .
- ( EN ) Esempi di mutazioni favorevoli , su gate.net . URL consultato il 17 maggio 2007 (archiviato dall' url originale il 27 settembre 2007) .
- ( EN ) IUPAC Gold Book, "mutation" , su goldbook.iupac.org .
- Mutazione genetica , in Treccani.it – Enciclopedie on line , Istituto dell'Enciclopedia Italiana.

| Controllo di autorità | Thesaurus BNCF 25353 · LCCN ( EN ) sh85089128 · GND ( DE ) 4170883-0 · BNF ( FR ) cb11932565x (data) |
|---|