Homo neanderthalensis
| Om de Neanderthal | |
|---|---|
 | |
| Starea de conservare | |
Fosil Perioada de fosilizare: Pleistocen superior ( Paleolitic mediu) | |
| Clasificare științifică | |
| Domeniu | Eukaryota |
| Regatul | Animalia |
| Sub-regat | Eumetazoa |
| Superphylum | Deuterostomie |
| Phylum | Chordata |
| ( cladă ) | Craniata |
| Subfilum | Vertebrate |
| Infraphylum | Gnathostomata |
| Superclasă | Tetrapoda |
| Clasă | Mammalia |
| Subclasă | Theria |
| Infraclasă | Eutheria |
| Superordine | Euarchontoglires |
| ( cladă ) | Euarchonta |
| Ordin | Primatele |
| Subordine | Haplorrhini |
| Infraordon | Simiiformes |
| Parvorder | Catarrhini |
| Superfamilie | Hominoidea |
| Familie | Hominidae |
| Subfamilie | Homininae |
| Trib | Hominini |
| Subtrib | Hominina |
| Tip | Homo |
| Specii | H. neanderthalensis |
| Nomenclatura binominala | |
| Homo neanderthalensis Regele , 1864 | |
| Sinonime | |
Homo sapiens neanderthalensis | |
| Denumiri comune | |
Neanderthal, Neanderthal, Paleantropo | |
Homo neanderthalensis , regele 1864 , cunoscut sub numele de Omul Neanderthal [1] , este un hominid strâns legat de Homo sapiens care a trăit în perioada paleolitică mijlocie, între 200.000 și 40.000 de ani în urmă.
Își ia numele de pe valea Neander ( Neandertal ) lângă Düsseldorf, în Germania , unde au fost găsite primele rămășițe fosile . El a fost un „ Homo ” foarte evoluat, care deținea tehnologii litice ridicate și un comportament social destul de avansat, ca sapiens din diferite perioade paleolitice.
Convivând în ultima perioadă a existenței sale cu însuși Homo sapiens , Homo neanderthalensis a dispărut într-un timp relativ scurt, eveniment care constituie o enigmă științifică care este studiată în mod activ astăzi.
Istorie
Rămășițele care au dat numele speciei au fost descoperite în august 1856 de către săpători de calcar într-o peșteră numită „Kleine Feldhofer” din Valea Neander , lângă orașul Feldhof din Germania . Descoperirile i-au fost arătate lui Johann Fuhlrott , profesor de științe, care a recunoscut că este o nouă specie din genul Homo . Descoperirea fosilelor a fost anunțată oficial abia la 4 februarie 1857. [2]
Documentat între 130.000 (pentru forme arhaice) și 30.000 (documentat cu înregistrări fosile) −22.000 (în absența fosilelor, dar cu dovezi culturale contestate) cu ani în urmă, în principal în Europa și Asia și, într-o măsură limitată, în Africa .
Unele studii din 2010 sugerează, printre unele ipoteze probabile legate de apropierea genetică dintre H. neanderthalensis și H. sapiens , că hibridizările dintre cele două ar fi putut avea loc în Orientul Apropiat cu aproximativ 80.000 și 50.000 de ani în urmă, datorită prezenței în om contemporan cu un procent între 1 și 4% din material genetic specific neanderthalian. Astfel de urme genetice sunt prezente la eurasiatici și nativi americani, dar nu și la africani subsaharieni: acest lucru sugerează, printre cel puțin patru ipoteze diferite, că hibridizarea ar fi putut avea loc în primele etape ale migrației speciei umane din Africa, probabil când a intrat în contact cu neanderthalienii care locuiau în Orientul Apropiat în urmă cu aproximativ 80.000 de ani. [3] [4]
În trecut, specia fusese numită și „omul neanderthalian”, din numele specific științific original, și Homo sapiens neanderthalensis când era încă considerată subspecie de homo sapiens; aceste denumiri sunt uneori încă găsite și același lucru se întâmplă și în alte limbi. Problema numelui ca subspecific nu este, spre deosebire de prima, doar formală, ci reflectă, așa cum se va explica mai târziu, diferite căi evolutive posibile și diferite grade de re-încrucișare cu sapiens .
Despre dispariția H. neanderthalensis sunt dezvoltate numeroase teorii.
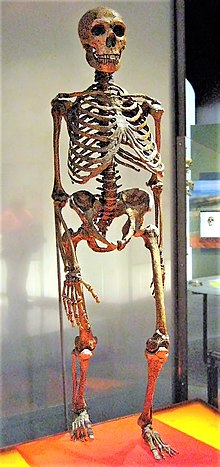
Sinteza aspectului extern
Aspectul fizic extern al Neanderthalului clasic în lumina cunoștințelor actuale, crescut semnificativ în comparație cu primele ipoteze și extrapolații din secolul al XIX-lea, este cel al unui om de înălțime medie-mică (1,60 m) perfect erect și foarte muscular robust, cu un raport suprafață / volum mai mic decât cel al altor specii de oameni, ceea ce duce la o dispersie mai mică a căldurii și, prin urmare, la o rezistență mai mare la temperaturi scăzute. Înălțimea medie, bazată pe 45 de oase lungi [5] a cel puțin 14 bărbați și 7 femele, a fost de aproximativ 164-168 cm pentru bărbați și aproximativ 152-156 cm pentru femele. [6] Eșantioanele a 26 de exemplare studiate în 2010 arată o greutate medie de 77,6 kg pentru bărbați și 66,4 kg pentru femele. [7]
Capul, comparativ cu un sapiens este alungit antero-posterior, chiar dacă se suprapune în general în variabilitatea sapiens , are un volum cerebral de 1 500 cm³ în medie, cu 10% mai mare decât bărbații actuali [8] și crestele proeminente ale frunții. Are un prognatism maxilar marcat, iar bărbia poate fi evazivă, cel puțin la tipurile arhaice. De-a lungul timpului, în unele zone și spre sfârșitul paleoliticului , un tip mai subțire se răspândește cu o bărbie osoasă mai pronunțată, în timp ce pomeții sunt mult mai puțin accentuați, iar arcurile sprâncenelor, dimpotrivă, mai proeminente.
O teză prezentată în 2006 și confirmată în 2007 [9] se bazează pe cercetări avansate cu tehnici de biologie moleculară și presupune că specia, în Europa, a dezvoltat indivizi cu ten alb cu păr roșu : tipul de pigmentare este în acord cu cei săraci iradiere solară ( ultravioletă ) a teritoriului colonizat, similar distribuției geografice actuale a pigmentării la tipurile umane. [10] În ciuda acestui fapt, s-a demonstrat că variabilitatea genetică a populației neandertaliene [11] [12] sugerează o variabilitate destul de largă a fenotipului , similară cu cea actuală a H. sapiens.
Studii recente, bazate pe analiza unor secvențe genetice ADNmt , sugerează că, fără a vorbi de subspecii, a existat cu siguranță o subdiviziune în trei (sau poate patru, dar metoda nu reușește încă să clarifice această ipoteză) mai multe grupuri mari de populație [13] . Existența reală a grupurilor din Europa de Sud (Iberia de Sud, Subalpină, Balcanică), Europa Centrală și de Est (din zona iberică de nord până la Marea Caspică) și Asia Centrală (până la granițele estice ale Kazahului) fusese anterior chestionată frecvent pe numai baza înregistrării fosile.
Tehnică, cultură și artă
Omul neanderthalian începe să evolueze într-un context cultural Acheulean superior, unde artefactele cu două fețe își schimbă forma, îmbunătățesc vârful și scad grosimea.
Noua tehnică de ciobire levalloisiană apare în industria litică (de la Levallois , la periferia Parisului ). Dintr-un nucleu litic inițial, aspru pentru a-l aduce la o formă biconvexă, lateral pe o față se despart aschii în formă regulată paralel cu un plan de bază. Această tehnică evoluează și formele numite amigdale (migdale) ale Acheuleanului dispar, deși la sud de Sahara va continua până la aproximativ 50.000 î.Hr.
În Europa, principalul teritoriu al Neanderthalului, se vorbește despre cultura Mousteriană , din descoperirile din Le Moustier, în Dordogne . Avem vârfuri triunghiulare, răzuitoare (pentru prepararea pieilor) foarte rafinate, cu o muchie de tăiere fin retușată. Mousterianul este împărțit în diferite culturi, geografice și cronologice (Mousterian de tradiție Acheuleană, Mousterian tipic, Mousterian denticulat, Charentian, Mousterian pontinian în Lazio etc.).
Trecerea după castelperronian [14] pare confirmată, cu o oarecare incertitudine reziduală, cu descoperiri atribuite în mod fiabil neandertalienilor, cum ar fi lame de ras, artefacte osoase și ornamente corporale.
Evoluția culturală
Încă nu există o viziune comună asupra evoluției culturale a Homo Neanderthalensis. În fața antropologilor ca Ian Tattersall care nu recunosc realizarea unor niveluri culturale care depășesc simpla tehnologie și, în special, nu împărtășesc faptul că viziunea simbolică a fost atinsă, înțeleasă ca o dimensiune simbolică în sens psihologic, o premisă la ritualuri, artă și comportamente relative, există indicații ale comportamentelor culturale avansate.

Culturile litice care vor evolua apoi ( Castelperroniano , Aurignaziano și foarte îndoielnic Gravettiano , cu siguranță împărtășite de sapiens ) sunt, prin urmare, încă studiate pentru atribuirea sigură posibilă neanderthalienilor; primul pas pare să fi fost constatat cu o semnificație bună.
În rezumat și, în linii mari, se poate spune că cultura neanderthaliană dominantă a fost Mousterianul și că limita superioară convențională (actuală) se află între Castelperronian și Aurignacian. Utilizarea pieilor este larg răspândită, de asemenea, pentru construcția de adăposturi de vară în aer liber, spre deosebire de practica troglodită de iarnă. Structurile de pietre sau oase se găsesc pentru a asigura marginile pielilor de sol.
Urmele abundente de ocru roșu sugerează utilizări rituale și religioase. Tot în această perspectivă, înmormântarea este evidențiată ca o practică larg răspândită, în gropi de formă ovală, cu echipamente funerare (alimente, coarne și unelte de piatră), adesea acoperite cu lespezi pentru a fura corpurile din târguri, depuneri de flori (studii privind polenul în găsește în Asia Mică ). Focul , în cercurile de izolare a pietrelor, este utilizat pe scară largă.
Potrivit unei cercetări în desfășurare la situl peșterii Bruniquel din Franța, neanderthalensis ar fi creat structuri complexe (a căror funcție este încă în curs de investigare) în urmă cu aproximativ 175.000 de ani folosind stalagmite special rupte și apoi aranjate în formațiuni concentrice., Demonstrând o organizare avansată, constructivă și poate capacitate simbolică. [15]

Poate (studiile sunt încă în desfășurare), cu neanderthalensis avem primul exemplu de instrument muzical nu de percuție ci în ton (în detaliu, cu patru note compatibile [16] cu scara diatonică greacă naturală), datorită descoperirii a așa-numitului flaut Divje Babe (în Slovenia ): un fragment al femurului unui urs peșteră perforat în mod regulat. Începe și arta figurativă în sens strict, considerată apanajul sapiens, dar din stratigrafia atribuită recent și neanderthalensis .
Progresele moleculare recente în studiul populațiilor din Neanderthal și poziția lor geografică, împreună cu cele din industria litică și alte artefacte, vor permite în viitor să clarifice mai bine relațiile dintre diferitele culturi și evoluția lor în spațiu și timp.
În ansamblu, tehnologia Homo Neanderthalensis poate fi rezumată după cum urmează:
- Toporile de mână sau amigdalele: sunt restul nodulilor mari de silex, ciupiți la margini pentru a face așchii mai mici.
- Puncte de silex: pentru a fi utilizate pe bețe grele utilizate ca sulițe la vânătoarea animalelor mari.
- Denticolati, adică cioburi de silex fără vârf cu margine zimțată: se spune că sunt fierăstrău de mână primitive, utilizate pentru prelucrarea lemnului, a oaselor și a tendoanelor.
- Răzuitoare: sunt cuțite de silex pentru a fi utilizate fără mâner, pentru tăierea cărnii.
- Flaute, adică oase lungi străpunse, care ar fi mai ușoare: bețe de lemn erau frecate în găuri pentru a aprinde paie.
Un articol publicat în Proceedings of the National Academy of Sciences la 13 noiembrie 2012 de un grup al Departamentului de Evoluție Umană, Institutul Max Planck de Antropologie Evolutivă din Leipzig [17] , aruncă o nouă lumină asupra activității simbolice și artistice din ce în ce mai probabile a neanderthalienilor. în tranziția de la paleoliticul mediu (MP) la paleoliticul superior (UP), o perioadă caracterizată prin înlocuirea ultimilor neandertali cu oameni moderni din Europa între 50.000 și 40.000 de ani în urmă.
Colecțiile de artefacte Castelperroniano (CP) găsite în centrul Franței și nordul Spaniei pot fi datate la acest interval de timp. Până acum, este singurul tip de colecție care a permis rămășițelor neandertaliene să fie asociate cu artefacte în stil UP. Artefactele CP includ, de asemenea, podoabe pentru corp, practic necunoscute în întreaga lume neanderthaliană. Cu toate acestea, s-a susținut că artefactele CP nu au fost fabricate de neanderthalieni, ci mai degrabă procesele de formare a sitului și amestecarea straturilor au condus la asocierea aleatorie a rămășițelor neandertaliene, ansamblurilor CP și ornamentelor corporale.
Încă din paleoliticul mediu, pasul de mesteacăn a fost utilizat pe scară largă de neanderthalieni ca lipici . Un studiu din 2019 a arătat cum aceste primele producții de pitch au fost realizate printr-o tehnică relativ simplă, folosind focare deschise [18] .
Autorii publică o serie de datări radiocarbonate obținute cu spectrometrie de accelerație de masă prin analiza colagenului osos ultrafiltrat extras din 40 de fragmente osoase bine conservate din Mousterianul târziu, CP și din straturile Protoaurignatiene de pe site-ul Grotte du Renne (la Arcy-sur- Cure, Franța). Rezultatele sunt incompatibile cu ipoteza amestecului. Mai mult, se raportează o dată calculată direct pe un fragment osos compact al tibiei neandertale găsite în Saint-Césaire (Franța). Această dată confirmă atribuirea artefactelor CP către ultimii neandertali din Europa de Vest. Mai mult, și mai important, datele stabilesc că producția de ornamente corporale în CP este ulterioară sosirii oamenilor moderni în regiunile adiacente ale Europei. Prin urmare, acest nou comportament ar putea fi rezultatul unei răspândiri culturale de la grupurile moderne la grupurile neandertaliene.
Distribuția teritorială
Omul neanderthalian este un gen originar [19] , [20] din Europa Centrală și de Vest , apoi a emigrat pe drumul spre Orientul Mijlociu , pe teritoriile actuale ale Irakului , Siriei și Israelului , cu puțini indivizi până în Asia Centrală ( Uzbekistan ) și Siberia .
Evoluția care a dus la apariția Homo neanderthalensis , sau „neandertalizare”, a fost lentă și progresivă, din grupuri europene izolate ( Homo erectus , Homo georgicus , Homo antecessor ). Poate fi urmat începând cu pre-neanderthalieni și până la neanderthalieni recenți.
- Vechii pre-neandertali
- Prima etapă corespunde fosilelor atribuite în general lui Homo heidelbergensis , posibil strămoș, conform unor teorii chiar și ale sapiensului modern: acesta este cazul Omului Tautavel (−400 000 de ani), găsit în Franța , al mandibulei lui Mauer ( −600 000 de ani), găsit lângă Heidelberg în Germania sau craniul Petralona ( Grecia ).
- Pre-neandertalieni recenți
- Prima etapă corespunde fosilelor din Swanscombe ( Anglia ), Steinheim (Germania) sau Sima de los Huesos à Atapuerca ( Spania ).
- Vechii neandertali
- Fosilele ulterioare cu trăsături incontestabil neandertaline au o vechime cuprinsă între -250.000 și -110.000 de ani. Putem menționa craniul Biache-Saint-Vaast ( Pas de Calais ), rămășițele la Chaise ( Charente ), mandibule din Montmaurin ( Alta-Garonne ), craniile italiene din Saccopastore ( Lazio ) sau materialul abundent al Krapina în Croația .
- Neanderthalii clasici
- Neanderthalii tipici, cu caractere derivate mai marcate, au o vârstă cuprinsă între -100.000 și -28.000 de ani, data ultimelor descoperiri fosile și, probabil, a dispariției lor.
Mai mult, pe lângă fosilele din Neanderthal în sine (aproximativ −42 000 de ani), există scheletele din La Chapelle-aux-Saints , din Moustier , din La Ferrassie , [21] din La Quina , din Saint-Césaire în sud-vestul Franței sau al Spionului din Belgia .
Ultimii neandertali cunoscuți au fost găsiți în Portugalia , Spania ( Zafarraya −30.000 ani [22] , [23] ), Croația (Vindija, −32.000 ani [24] , [25] ) și nord-vestul Caucazului (Mezmaiskaya, −29.000 ani) ). După cum sa văzut deja, aceste date trebuie luate în considerare cu prudență și sunt deseori controversate.
Cercetările efectuate din 1999 până în 2005 pe Peștera Gorham din Gibraltar sugerează că neanderthalienii au locuit-o de la -28.000 la -24.000 de ani [26] , [27] . De aici o lungă conviețuire cu Homo sapiens , prezentă geografic de la -32 000 de ani. Joao Zilhao, de la Universitatea din Bristol, critică cu tărie aceste ultime descoperiri mutând data dispariției în urmă cu 37.000 de ani în urmă. Diferitele teorii cu privire la datele de dispariție și de conviețuire cu Hss în prezent pot fi definite doar în faza de dezvoltare și verificare, de asemenea, în lumina descoperirilor mereu noi, de asemenea ale al treilea co-locuitor, în intervalul de timp de 30.000 - 50.000 de ani . în urmă [28] .
Antropologia fizică
Următoarea este o listă de trăsături fizice care disting neandertalienii de oamenii moderni; cu toate acestea, nu toate pot fi folosite pentru a distinge populații specifice neandertaliene, provenind din diferite zone geografice sau perioade de evoluție, de alte specii umane dispărute. Mai mult, multe dintre aceste trăsături apar ocazional la bărbații moderni, în special în rândul anumitor grupuri etnice. Nu se știe încă nimic despre culoarea pielii sau despre forma părților moi, cum ar fi ochii, urechile și buzele neandertalienilor, cu excepția modelării.
Comparativ cu oamenii moderni, neanderthalienii aveau o constituție mai robustă și aveau caractere morfologice distincte, în special în ceea ce privește craniul , care a acumulat treptat mai multe aspecte derivate (în sens cladistic ), în special în regiunile geografice relativ izolate. Și aici trebuie remarcat faptul că populațiile anatomice moderne din paleoliticul mediu (aproape de est ca în Skul și Qafzeh), erau la fel de masive. Se consideră că statura lor relativ înaltă, dar robustă, este o adaptare la climatul rece al Europei în timpul Pleistocenului ( legea lui Allen ), deși în realitate aceste trăsături par a fi deja prezente în perioadele calde.
Tabelul următor este un rezumat, cu adăugiri, al recenziei pe Sc. American de E. Trinkaus și WW Howells pe Neanderthal.
| Cranian | Post-cranian |
|---|---|
| Fossa soprainiaca, o canelură deasupra inionului sau a liniei superioare a cefei. | În general, sensibil robust, dar sapiens coeval au fost la fel de robuste. |
| Bump occipital , o protuberanță a osului occipital . | Extremitățile degetelor (tuberozitate) largi și rotunjite, inserții puternice ale tendoanelor relative. |
| Prognatism , marginea celui mai avansat prim molar al pomeții | Cusca toracică tinde să fie cilindrică |
| Craniul plat plat (platicefalia) și alungit anterior posterior | Genunchii mari |
| Baza craniului plat, văzută din spate cu o formă rotunjită, spre deosebire de cea sapiens pentagonală | Oase lungi ale gâtului |
| Torul supraorbital , un arc superciliar proeminent, trabeculat (spongiform) | Omoplatul cu cavitate dorsală, opus celui modern cu o cavitate ventrală predominantă (4/5), pentru o mai bună inserție a mușchiului teres minor |
| 1 200 - 1 750 cm³ de capacitate craniană (cu 10% mai mare decât media umană actuală) | Femurul gros și curbat, indicând o putere musculară bună |
| Barbă osoasă nu foarte proeminentă, de fapt adesea mascată de prognatism; la exemplarele mai recente caracterul nu este evident | Tibia și fibula scurtă, raza idem și ulna |
| Creasta procesului mastoidian din spatele deschiderii auriculare | Osul pubian grațios și alungit al ambelor sexe (ramura pubiană superioară), mărirea probabilă a canalului de naștere |
| Nu există caneluri pe dinții canini | Extinderea suprafețelor articulare ale astragalului |
| Diastema posterioară până la al treilea molar, consecință a alungirii maxilare | Inserții musculare puternice pe arcul plantar al piciorului (împingere puternică la alergare) |
| Proiecții osoase pe laturile deschiderii nazale | |
| Forma specială a labirintului , în urechea internă | |
| Foramen mandibular mare pentru circulația sângelui prevăzut cu o punte osoasă, la sapiens în general cu duș. | |
| Nas larg, dar nu turtit |
Craniometrie
O statistică precisă, bazată pe analiza multivariată a formei și dimensiunii craniului, totuși, în timp ce detectează un habitus tipic neanderthalian, arată o apropiere surprinzătoare de craniile actuale de forme extreme (C. Stringer, de la British Museum).
Pigmentare
Dintr-un studiu din 2001, unii comentatori speculează că neanderthalienii demonstrează un tip cu părul roșu, împărtășind moștenirea cu bărbații actuali de tip pistruiat și roșcat; [29] în orice caz, alți cercetători ai vremii nu au fost de acord. [30] Studiile moleculare din 2007 citate în paragraful inițial Sinteza aspectului extern risipesc orice îndoieli, adică pe baza studiului asupra variantelor receptorului melanocortinei 1 (mc1r) (sau receptorului hormonului care stimulează melanocitele (MSHR) ), receptorul peptidic care activează melanina sau receptorul melanotropinei) o clasă de receptori celulari, receptori cuplați la proteinele G , care sunt liganzi ai hormonilor peptidici hipofizari , rezultă că funcția redusă a acestor porturi de a dezvolta un fenotip puțin pigmentat în mod clar și pielea părului roșu. Studiul a fost realizat cu ajutorul unor tehnici de amplificare a genei pe o gena responsabila pentru pigmentarea uman, și anume cel legat de receptor al melanocortinei 1 (MC1R), care codifică șapte transmembranar GTP-ligand (proteina G) receptor -coupled (GPCR) proteine heterometric . Părul roșu și pielea deschisă rezultă din alele care implică pierderea completă sau parțială a funcției MC1R uman (huMC1R), modificând echilibrul în sinteza eumelanină-feomelanină . Probele au provenit din ADN bine conservat (pe baza datelor referitoare la conținutul abundent de aminoacizi și la racemizarea sa slabă) a unui exemplar din Munții Lessini (în Italia) și din exemplarul El Sidrón 1252 (Cueva de El Sidrón, în Spania în Asturia).
Dezvoltarea fizică
O altă diferență, potrivit paleoantropologilor, ar fi putut fi în creștere, de fapt, se crede, analizând creșterea primului molar comparativ cu cea a dinților Homo sapiens, că neandertalienii au crescut mai repede decât bărbații moderni, ajungând la vârsta adultă deja la 15 ani. Această creștere rapidă, potrivit experților, este o caracteristică tipică a primelor hominide [ neclar ] . Potrivit cercetătorilor, perioadele de dezvoltare mai rapide par să fi fost o necesitate pentru o specie care trăia în țări foarte reci și inospitaliere, unde mortalitatea infantilă era foarte mare și speranța de viață era scurtă.
Hibridizare cu Homo sapiens
Acest articol sau secțiune referitoare la antropologie este considerat a fi verificat . |

În a doua jumătate a secolului al XX-lea a existat o mare dezbatere în domeniul academic dacă omul neandertalian trebuie considerat o subspecie dispărută a Homo sapiens (numită Homo sapiens neanderthalensis ), cu posibilitatea consecventă de încrucișare și hibridizare între cele două, sau dacă este o specie autonomă ( Homo neanderthalensis ). Oricum ar fi, cele două grupuri au coexistat mii de ani în Europa și Asia Mică. Investigațiile genetice efectuate în 2008 asupra ADN-ului fosil recuperat pe unele rămășițe umane din Neanderthal par să indice că cele două tipuri umane parțial contemporane aparțineau a două specii diferite [31], cu toate că descoperiri mai recente referitoare la genomul uman par să poată stabili că neanderthalensis iar sapiens ar aparține aceleiași specii și că diferențele anatomice și somatice s-ar datora cauzelor climatice, de mediu, morfologice și dietetice [32] .
Analiza ADN
Biologul molecular suedez Svante Pääbo a testat cel puțin 70 de probe din neanderthalieni înainte de a găsi unul suficient pentru analiza ADN; inițial secvențele osoase vechi de 38.000 de ani de pe site-ul croat Vindija arată 99,5% partajarea ADN-ului cu sapiens : cei doi aparținând genului Homo s-au îndepărtat de acest lucru de aproximativ 500.000 de ani. Un articol din revista Nature estimează divergența la 516.000 de ani, unde dovezile paleontologice, suficient de în acord, arată o perioadă de 400.000 de ani. Svante Pääbo a propus un proiect pentru reconstrucția totală a patrimoniului genetic al Homo neanderthalensis . Edward Rubin de la Lawrence Berkeley National Laboratory din Berkeley , California , arată că genomul neanderthalian este similar cu 99,5 până la 99,9% cu al nostru. [33] [34]
În noiembrie 2006, un nou articol este publicat în revista științifică Proceedings of the National Academy of Sciences , unde un grup de cercetare european propune tema hibridizării [35] . În același timp, pe 16 noiembrie 2006 Science Daily publică teza opusă, dezvoltată de Berkeley Lab și Joint Genome Institute, conform căreia nu există dovezi de încrucișări între cele două grupuri umane, iar suprapunerea genetică nu depășește 99,5 %.
Edward Rubin, directorul ambelor institute, concluzionează:
| ( EN ) "Cu toate că nu putem concluziona definitiv că nu s-a produs încrucișarea între cele două specii de oameni, analiza ADN-ului nuclear din Neanderthal sugerează probabilitatea redusă ca acesta să aibă loc la orice nivel apreciabil." | ( IT ) "Deși nu suntem în măsură să afirmăm definitiv că hibridizarea dintre cele două specii nu a avut loc, analiza ADN-ului nuclear din Neanderthal sugerează o probabilitate redusă ca acest lucru să aibă loc la un nivel apreciabil." |
Cea mai precisă analiză moleculară partajată la nivel internațional și confirmată în prezent estimează că H. sapiens și H. neanderthalensis s-au separat în urmă cu nu mai mult de 800 000 de ani și încă nu există dovezi clare acceptate de majoritatea comunității științifice cu privire la o posibilă reunificare. [36] În prezent, comunitatea științifică, pe baza unor studii efectuate de Svante Pääbo de la Institutul Max Planck din Leipzig, consideră încă posibilă, printre diferitele ipoteze, o reuniune genetică între cele două specii, pe baza prezenței în genom al unei părți a populației umane actuale (cu excepția africanilor), aproximativ 1-4% material genetic compatibil cu cel neanderthalian.
Într-un articol suplimentar al lui Svante Pääbo, în colaborare cu Departamentul de Genetică de la Harvard și MIT, este propusă o datare a ultimei perioade de schimb de gene între neandertali și oameni moderni (H. sapiens sapiens) [37] .
Una delle scoperte chiave dell'analisi del genoma di Neandertal è che i Neandertal condividono più varianti genetiche con non-africani che con gli africani. Questa osservazione è in linea con due ipotesi: 1) l'incrocio tra Neandertal e gli esseri umani moderni è avvenuto dopo che gli umani moderni sono usciti dell'Africa o 2) la struttura delle popolazioni degli antenati di Neandertal e di quelli degli esseri umani moderni erano in parte simili. Queste ipotesi possono fare previsioni diverse sulla data dell'ultimo scambio di geni tra gli antenati di Neandertal e gli umani moderni non africani. Gli autori stimano questa data misurando il grado di linkage disequilibrium (LD) (spesso definita "associazione allelica"; particolari alleli in due o più loci vicini mostrano associazione allelica se si verificano con frequenze significativamente diverse da quelle previste in base alle frequenze alleliche dei singoli) tra il genoma degli attuali europei e trovano che l'ultimo flusso genico da Neandertal agli europei si è probabilmente verificato 37 000-86 000 anni fa (BP), e più probabilmente 47 000-65 000 anni BP. Ciò supporta l'ipotesi di un incrocio recente e suggerisce che l'incrocio si è verificato quando gli esseri umani moderni portatori delle tecnologie del Paleolitico superiore incontrarono i Neandertal a seguito della loro espansione fuori dall'Africa.
Le tracce neandertaliane più recenti
Si dibatte lungamente sugli ultimi rappresentanti noti dei nostri parenti più prossimi. Quest'argomento, assieme al mistero della scomparsa della popolazione neanderthaliana, è forse uno dei più controversi della scienza paleoantropologica e il più soggetto a evoluzione. Le datazioni tramite spettrometria di massa da parte di Fred Smith ed Erik Trinkaus, (Northern Illinois University e Washington University rispettivamente), da scavi di Vindija (Croazia) portano a 28 000 anni i reperti più recenti, con articoli pubblicati a cavallo del 2000. Nuove datazioni ricollocano indietro [38] a 32 000 anni i reperti. Vengono scoperti altri fossili recenti sulle coste atlantiche del Portogallo, e si riscontrano evoluzioni morfologiche verso una maggiore modernità. Contemporaneamente viene messa in discussione l'associazione di alcune culture litiche ( Aurignaziano , Musteriano ) ai vari Homo .
La scomparsa dell'intera popolazione neanderthaliana
La teoria del "rimpiazzamento"

Si dibatte sulla completa scomparsa della popolazione, non perfettamente spiegabile sulla base delle sole caratteristiche fisiche degli individui. Si trattava di una specie lungamente adattata all'ambiente colonizzato, con un volume cranico pari o superiore ai sapiens attuali, e di cultura tecnica almeno inizialmente sovrapponibile nelle due popolazioni. Le prime ipotesi teorizzate parlano di lenta ibridazione con Sapiens moderni , eliminazione fisica ( genocidio ), competizione , o selezione sessuale. La difficoltà ad analizzare lo scarso materiale genetico sopravvissuto completa il quadro [39] . Certamente la lunga coesistenza di uomo di Neandertal e uomo moderno pongono sul tavolo della discussione molti problemi irrisolti e l'argomento è in costante riscrittura.
Nel 2005 sul Journal of Economic Behaviour and Organization Jason Shogren , economista dell'Università del Wyoming di Laramie , pubblica con i suoi collaboratori un articolo in cui avanza una teoria sulla scomparsa dell'uomo di Neandertal [40] . Lo studioso avanza l'ipotesi che H. neanderthalensis si sia dovuto scontrare con la particolare cultura dell' H. sapiens : questa cultura si basava su tecniche avanzate di commercio , cosa che portava più tempo libero rispetto a una cultura basata sulla caccia . Il tempo libero ottenuto avrebbe permesso lo sviluppo di specializzazioni non strettamente legate alla sussistenza, come costruire utensili sempre più complessi o dedicarsi all'arte. La complessità e la versatilità di una tale cultura avrebbe avuto esito fatale per la più "tradizionale" cultura dei Neandertal.
Stephen Kuhn e Mary Stiner dell'università dell' Arizona , sulla rivista Current Anthropology , propongono, documentandola, la tesi per cui la principale causa di estinzione fu la mancata suddivisione dei lavori tra i sessi. I più organizzati sapiens, più efficientemente, poterono competere affidando alle donne compiti stanziali, e meno gravosi, affidando ai maschi i ruoli di cacciatori e approvvigionatori di materiali. La prole, protetta e anch'essa stanziale, avrebbe avuto più possibilità di sopravvivenza [41] . Un'altra ipotesi sulla loro estinzione, fa riferimento invece ai drastici cambiamenti nella composizione della magnetosfera terrestre, dovuti all'azione di forti variazioni del campo magnetico terrestre , con conseguente variazione della radiazione solare che avrebbe comportato, a sua volta, mutazioni negative per la specie, fino a portarla all'estinzione. [42]
Due caratteristiche dei neanderthalensis vanno rimarcate. La prima è che il loro fisico era strutturato per esprimere al meglio la forza, mentre quello di alcuni, ma non tutti, i sapiens loro contemporanei come i Cromagnonoidi , a gambe più lunghe ea bacino più stretto e compatto, privilegiava le capacità di resistenza nella corsa. Quindi quei sapiens sarebbero stati più resistenti nel percorso di sensibili distanze con quel tipo di andatura, ma svantaggiati nella forza fisica e nella lotta. Una caratteristica che si pensava differenziasse sapiens e Neandertal, la diversa alimentazione, con sapiens marcatamente onnivori e Neandertal carnivori si è variamente rivelata parziale e dipendente esclusivamente da singole situazioni. Entrambi ad esempio erano specie ben adattate agli ambienti costieri [43] con un'alimentazione basata su frutta e verdura, prodotti della pesca, raccolta di molluschi e caccia [44] . Soggetta a periodiche discussioni è anche la capacità dei Neandertal di cacciare grosse prede (come la megafauna ); mentre esistono prove che anche i più antichi esponenti della nostra specie utilizzassero zagaglie , e altri ordigni da lancio, non si sa se i Neandertal utilizzassero armi da lancio. Avrebbero cioè dovuto approcciarsi alle prede (ea eventuali nemici) a distanza ravvicinata. I Neandertal, almeno alcune popolazioni, erano comunque prettamente carnivori [45] e superpredatori .
La teoria dell'"ibridazione"
Uno studio pubblicato su Science Advance nel 2020 afferma che circa 50.000 anni fa, mentre Homo sapiens si diffondeva fuori dell'Africa, avvennero accoppiamenti fertili con popolazioni neandertaliane e denisoviane sino al loro assorbimento e sostituzione con popolazioni tipicamente sapiens ma nel cui genoma rimase traccia dell'avvenuta mescolanza genetica. [46]
Nel 2010 sono stati pubblicati i risultati di una ricerca quadriennale che mirava a mappare il genoma dei Neanderthal, è stato comparato del DNA raccolto da dei fossili di Neanderthal con del Dna degli umani contemporanei, ei risultati hanno stupefatto la comunità scientifica, nel DNA unico condiviso delle moderne popolazioni una porzione rilevante tra 1 e il 4% era costituito da DNA Neanderthaliano. [ senza fonte ]
Ricostruzioni dell'aspetto, secondo la scienza e gli stereotipi


«Se [un uomo di Neandertal] si potesse reincarnare e porre nella metropolitana di New York , opportunamente lavato, sbarbato e modernamente vestito, si dubita che potrebbe attirare più attenzione di alcuni dei suoi altri abitanti.» |
| ( William Straus [47] ) |


Molto ma non tutto si può dire del vero aspetto di questo ominide ricostruendolo dai suoi resti fossili. La scienza contemporanea ha messo a punto varie tecniche per la ricostruzione delle parti non fossilizzabili, i tessuti molli, a partire da elementi quali le inserzioni tendinee sulle ossa, le linee di forza sulle stesse, i livelli di consunzione dei denti e altro ancora. La biologia molecolare ha poi fornito ulteriori elementi per valutare l'espressione genica di caratteri non conservabili, appunto come il colore dei capelli. Il Neandertal possedeva la tecnologia necessaria a confezionare indumenti; a tale scopo utilizzava prevalentemente pelli, per la concia delle quali costruiva utensili quali i raschiatoi musteriani . Le zone climatiche frequentate imponevano sicuramente l'uso di coperture, e possiamo rappresentare il Neandertal tipico prevalentemente vestito.
Oltre a ciò si segnala la capacità simbolica e artistica, che ha portato all'uso, almeno episodico, di monili e pendagli [48] . In passato, al contrario, è spesso stato rappresentato come lo stereotipo dell'uomo cavernicolo, tale ricostruzione ripete né più né meno la figura dell' uomo selvatico presente nella tradizione popolare europea e raffigurata sin dal Medioevo. Secondo alcune interpretazioni di studiosi di criptozoologia , non di antropologi o paleontologi, poteva però essere di aspetto scimmiesco [49] , dotato di un fitto villo pilifero distribuito su tutta la superficie del corpo che lo proteggeva dal clima freddo europeo.
Un'immagine da alcuni supposta e interpretata di Neandertal è una rappresentazione preistorica nella caverna di Isturits sui bassi Pirenei francesi, frequentata da 80 000 a 10 000 anni fa prima dai Neandertal e poi dai Cro-magnon. Vi compare la rappresentazione di una testa-trofeo di un essere antropomorfo con testa ovale senza fronte e mento, collo grosso, occhi a mandorla infossati sotto grandi arcate frontali, con un grosso naso, orecchie piccole, con capelli corti a spazzola e dritti a cresta sulla testa che proseguivano sulla schiena in una sorta di criniera equina, e peli fitti simili su quasi tutta la faccia e collo simili al manto di un cervo. [ senza fonte ]
Storia dei principali reperti fossili europei


I resti rinvenuti nel 1856 da Johann Fuhlrott nella valle di Neander consistevano nella parte superiore del cranio , alcune ossa, parte dell'osso pelvico , alcune costole , e ossa del braccio e della spalla.
In precedenza erano stati scoperti altri fossili, infatti già nel 1829 nel Belgio venne trovato parte di un cranio di un bambino di due anni e mezzo. Questi, però, venne riconosciuto come arcaico soltanto nel 1836 . Nel 1848 a Gibilterra venne trovato un cranio adulto, poi battezzato Gibraltar 1 , ma la sua esistenza rimase sconosciuta alla scienza fino al 1864 , quando venne riconosciuto come appartenente a un individuo Neandertal.
Altri due scheletri di Homo neanderthalensis , di almeno 60 000 anni, vennero trovati in Belgio nel 1886 da Marcel de Puydt e Max Lohest . Altri rinvenimenti importanti vennero fatti in Croazia nel 1899 da Dragutin Gorjanovic-Kramberger e nel 1908 in Francia a La Chapelle-aux-Saints da Jean Bouyssonie che rinvenne lo scheletro di un uomo anziano, risalente a 50 000 anni, il cui volume cranico era di ben 1620 centimetri cubi.

Nel 1939 venne rinvenuto nella grotta Guattari a San Felice Circeo , un cranio presumibilmente appartenente a H. neanderthalensis .
Nel secondo dopo guerra emersero ancora altri resti importanti; tra il 1953 e il 1960 nella grotta di Shanidar in Iraq vennero scoperti 9 scheletri di uomini di Neandertal, risalenti a un periodo compreso tra i 70 ei 40 000 anni fa, e nel 1979 nel villaggio di Saint-Césaire in Francia uno scheletro completo risalente a 35 000 anni fa.
Nel 1868 a Cro-Magnon in Francia vennero trovati da alcuni operai i resti di un uomo risalenti a 28 000 anni fa; era venuto alla luce uno dei più antichi progenitori della nostra specie ( Homo sapiens ). Era un rappresentante della nostra specie umana, che proveniente dall'Africa o dall'Asia, migrando stava insediandosi in Europa, confinando verso la penisola iberica gli ultimi Neandertal.
Resti neandertaliani rinvenuti in territorio italiano
I resti portati alla luce in Italia non sono molto numerosi rispetto all' Europa continentale. Si distingue in ogni caso tra ritrovamenti fossili (rari) e ritrovamenti di tracce e manufatti, non sempre associati. Nel secondo caso, alcuni siti sono discussi; alcune culture, infatti, furono condivise con soggetti appartenenti al genere Homo. Per i siti composti da soli reperti litici si veda la voce Paleolitico .
- Si riportano i siti tardo pleistocenici con reperti ominidi fossili in Italia, definibili di caratterizzazione Neandertaliana
Piemonte
- Monte Fenera ( Ciota Ciara , Ciutarun) Vercelli : frammenti cranici, denti
Liguria
- Grotte dei Balzi Rossi . Caverne e grotte a strapiombo sul mare, rosse dalla ricchezza di composti ferrosi nel terreno, vicino al confine francese, monte Bellinda a Grimaldi , frazione di Ventimiglia . Ritrovamenti litici appartenenti al Musteriano . Abitate in successione da diverse culture e popolazioni, dal paleolitico medio fino al neolitico. Sepolture, principalmente Sapiens, utensili litici, arte rupestre, spesso di incerta attribuzione. I pochi fossili ritrovati sono di tipo pre-neandertaliano o neandertaliano antico medio pleistocenico.
- Caverna delle Fate , Finale Ligure ( Savona ) [50] manufatti Litici di tipo Levalloisiano e Musteriano datazione 100 000 - 50 000 aC, ancora allo studio per una datazione palinologica . Frammenti ossei vari, di scavo anni novanta ; si son conservati resti ossei, almeno di cinque Neandertal bambini e adulti, caso unico nell'archeologia dell'Italia settentrionale [51] . Nella vicina Arma delle Mànie si ritrovano ugualmente tracce neandertaliane.
- Grotte di Toirano , tra Albenga e Savona , molto vaste e concrezionate. Nella Grotta delle streghe, o della Basura, si rinviene un'impronta fossile di un piede giovane [52] , (ma alcune datazioni radiometriche correlate alle impronte hanno stimato un'età di 12 000 anni, nel qual caso sarebbero allo stato attuale delle conoscenze, attribuibili solamente a Homo sapiens sapiens) [ senza fonte ] e molti resti animali ( Ursus spelaeus ). Le grotte e le loro testimonianze fossili, frequentate sicuramente dall' Homo heidelbergensis sino ai sapiens del neolitico, sono tuttora allo studio.
Veneto
- Riparo Tagliente , Monti Lessini, Verona : denti [53] . [54]
- Grotta di Fumane , Verona : denti.
- Grotta San Bernardino Vicenza : falange, denti.
Toscana
- Buca del Tasso , presso Camaiore Toscana nord-occidentale ( Lucca ). La presenza dei resti scheletrici neandertaliani è documentata dal femore attrbiutio ad un bambino di circa 9 anni di età. [55] [56] .
Lazio
- Monte Circeo , Grotta Guattari , ( Latina ). Il ritrovamento il 24 febbraio 1939 da parte del prof. Alberto Carlo Blanc fece a lungo discutere per l'interpretazione fortemente discussa di un cranio con segni di aggressione e svuotamento encefalico. Per mezzo secolo si discusse sull'antropofagia rituale della specie. Studi tafonomici successivi spiegarono lo svuotamento encefalico come azione di iene . Scavi archeologici tra il 2019 e il 2021 hanno riportato alla luce resti di altri 9 individui, portando ad 11 il numero di Homo neanderthalensis ritrovati nel sito. [57]
- Monte Circeo , ( Latina ), Grotta del Fossellone : mandibola.
- Monte Circeo , ( Latina ), Grotta Breuil : al suo interno, gli scavi del 1986 hanno rinvenuto una porzione postero-inferiore di parietale, una corona di primo molare inferiore di adulto e il terzo molare inferiore di un individuo giovane (circa 13 anni) [ senza fonte ] .
- Roma , Cava di Saccopastore . Ritrovamento nel 1929 e 1935 di due soggetti , di tipo arcaico databili circa 120 000 anni [58] .
Abruzzo
Campania
- Grotta Taddeo, Marina di Camerota , ( Salerno ): denti.
- Il Molare, a Scario , dove si ritrova una mandibola.
- Grotta di Vallicelli, a Monte San Giacomo , industria litica.
Puglia
- Sulla costa del Salento (Grotta Romanelli).
- A Capo di Leuca , ( Lecce ), la Grotta delle Tre Porte. Nella sala denominata 'Antro del bambino' insieme a resti di fauna pleistocenica si ritrova il molare fossile appartenuto a un bimbo Neandertal di circa dieci anni [59] [60] [61] .
- Grotta di Santa Croce , Bisceglie , ( Bari ): femore e frammento cranico.
- Grotta di Lamalunga , nei pressi di Altamura ( Bari ): scheletro completo, denominato Uomo di Altamura ).
- Grotta del Cavallo, Nardò , ( Lecce )
- Grotta-Riparo Uluzzo , Nardò , ( Lecce ): denti.
- Fondo Cattìe, Maglie , ( Lecce ) [62] , [63] .
- Cava Nuzzo, Melpignano ( Lecce ): dente.
Calabria
- Contrada Ianni di San Calogero, Nicotera , ( Vibo Valentia ): porzione cranica.
- San Francesco di Archi , ( Reggio Calabria ): mandibola.
Solamente reperti litici
Lombardia
- Grotta dell'Orso (Monte Generoso – San Fedele Intelvi – CO), piccoli reperti in selce. [64]
- Cascina Barbabella di Bagaggera ( Parco regionale di Montevecchia e della Valle del Curone – Rovagnate – LC), resti di focolari e concentrazioni di manufatti in selce [ senza fonte ]
- Buco del piombo [65] ( Erba – CO)
Toscana
Nella cultura di massa
- Nella serie a fumetti Martin Mystère , creata da Alfredo Castelli ed edita dalla Bonelli , Java , il compagno di avventure del protagonista, è un uomo di Neandertal appartenente a una tribù della Mongolia, unica sopravvissuta all'estinzione della specie. Le caratteristiche fisiche sono leggermente differenti dalla realtà, in particolare è più alto e molto prestante fisicamente, mentre permane la caratteristica del mutismo, infatti Java si esprime con suoni gutturali ea gesti, aiutandosi anche con la lingua dei segni .
- In Far Cry Primal , gli Udam sono descritti come neandertaliani, benché questi fossero già estinti.
- In Topolino all'età della pietra ( The Land of Long Ago ), riduzione a fumetti del romanzo Il mondo perduto di Arthur Conan Doyle , il professor Ossivecchi ha come assistente Oompa, un uomo di Neanderthal, che si esprime a gesti e al quale il professore ha insegnato qualche parola. [67]
Nella letteratura e nel cinema
Numerosi romanzi e racconti vedono tra i protagonisti uno o più Neandertaliani:
- Jean M. Auel , Ayla figlia della Terra .
- L. Sprague De Camp , L'uomo nodoso (The gnarly man) , 1939.
- Lester Del Rey , Il giorno è compiuto (The Day Is Done) , 1939.
- William Golding , Uomini nudi (The inheritors) , 1955.
- Michael Crichton , Mangiatori di morte (Eaters of the Dead) , 1976.
- Robert J. Sawyer , Trilogia dei Neanderthal (Neanderthal Parallax) , 2002 - 2003 .
- Isaac Asimov e Robert Silverberg , Il figlio del tempo (The Ugly Little Boy) , 1992 , basato sull'omonimo racconto di Asimov del 1958
- Stanis Mulas , L'uomo dei Balzi Rossi , 1988.
- Ben Bova , Orion , 1984.
Nel cinema:
- La guerra del fuoco (La guerre du feu) di Jean-Jacques Annaud ( 1981 )
- Il 13º guerriero di John McTiernan ( 1999 ), adattamento cinematografico di Mangiatori di morte di Crichton
- L'era glaciale di Chris Wedge ( 2002 )
- Una notte al museo di Shawn Levy ( 2006 )
- Una notte al museo 2 - La fuga di Shawn Levy ( 2009 )
- Ao, l'ultimo dei Neanderthal di Jacques Malaterre ( 2010 )
- I Croods di Kirk DeMicco e Chris Sanders ( 2013 )
- Una notte al museo 3 - Il segreto del faraone di Shawn Levy ( 2014 )
- Il viaggio di Arlo di Peter Sohn ( 2015 )
Note
- ^ Il termine Neanderthal , scritto con "h", è corretto solo nel nome latino Homo neanderthalensis in quanto, secondo l'ortografia tedesca attuale, la parola Tal (valle) ha perso la "h" che aveva prima delle regolamentazioni linguistiche sancite dalla Conferenza ortografica del 1901Thal è un vecchio modo di scrittura di Tal. Questo, in seguito alle decisioni prese dalla Conferenza ortografica del 1901, non è più corretto. (Thal ist eine alte Schreibweise von Tal. Sie ist gemäß den Beschlüssen der Orthographischen Konferenz von 1901 seither nicht mehr korrekt.) ( DE ) Thal - Wiktionary , su de.wiktionary.org . URL consultato il 14 marzo 2017 .
- ^ Neanderthal Museum, Storia del ritrovamento
- ^ Richard E. Green et al, A Draft Sequence of the Neanderthal Genome , in Science , vol. 328, n. 5979, pp. 710–722, DOI : 10.1126/science.1188021 .
- ^ Paul Rincon, Neanderthal genes 'survive in us' , su BBC News , BBC , 6 maggio 2010. URL consultato il 7 maggio 2010 .
- ^ Le ossa lunghe sono quelle che si sviluppano in lunghezza e sono, in particolare:
- ^ H. Helmuth, Body height, body mass and surface area of the Neanderthals. , in Z Morphol Anthropol , vol. 82, n. 1, 1998, pp. 1-12, PMID 9850627 .
- ^ Froehle, Andrew W; Churchill, Steven E (2009), Energetic Competition Between Neandertals and Anatomically Modern Humans , (PDF), PaleoAnthropology: 96–116, 2009; collegamento consultato il 31 ottobre 2011.
- ^ ( EN ) MS Ponce de León, L. Golovanova, V. Doronichev, G. Romanova, T. Akazawa, O. Kondo, H. Ishida e CP Zollikofer, Neanderthal brain size at birth provides insights into the evolution of human life history , in PNAS , n. 105, 2008, pp. 13764-13768.
- ^ ( EN ) Carles Lalueza-Fox et al. , A Melanocortin 1 Receptor Allele Suggests Varying Pigmentation Among Neanderthals. , su sciencemag.org , 25 ottobre 2007.
- ^ Articolo di sintesi [ collegamento interrotto ] , su repubblica.it .
- ^ ( EN ) Laurent Excoffier, Neanderthal Genetic Diversity: A Fresh Look from Old Samples Current Biology , vol. 16, 22 agosto 2006, pp. R650-R652.
- ^ ( EN ) Carles Lalueza-Fox, In Neanderthals, there was probably the whole range of hair colour we see today in modern European populations, from dark to blond right through to red .
- ^ ( EN ) V. Fabre, S. Condemi e A. Degioanni, Genetic Evidence of Geographical Groups among Neanderthals , in PLoS ONE , 4(4), 2009, p. e5151, DOI : 10.1371/journal.pone.0005151 .
- ^ ( EN ) Jean-Jacques Hublina, Sahra Talamo, Michèle Julien, Francine David, Nelly Connetb, Pierre Bodub, Bernard Vandermeerschd e Michael P. Richardsa, Radiocarbon dates from the Grotte du Renne and Saint-Césaire support a Neanderthal origin for the Châtelperronian , in PNAS , vol. 109, n. 46, 29 ottobre 2012, pp. 18743-18748, DOI : 10.1073/pnas.1212924109 .
- ^ ( EN ) Jacques Jaubert, Sophie Verheyden e Dominique Genty, Early Neanderthal constructions deep in Bruniquel Cave in southwestern France , in Nature , advance online publication, 25 maggio 2016, DOI : 10.1038/nature18291 . URL consultato il 26 maggio 2016 .
- ^ Bob Fink, Neanderthal Flute , Greenwich, 1997, ISBN 0-912424-12-5 . WorldCat.org )
- ^ JJ Hublin et al. , Radiocarbon dates from the Grotte du Renne and Saint-Cesaire support a Neanderthal origin for the Chatelperronian , in Proc Natl Acad Sci US A. , vol. 109, n. 46, 13 novembre 2012, pp. 18743-8, DOI : 10.1073/pnas.1212924109 .
- ^ ( EN ) Schmidt, P., Blessing, M., Rageot, M., Iovita, R., Pfleging, J., Nickel, KG; Righetti, L. & Tennie, C., Birch tar extraction does not prove Neanderthal behavioral complexity , in PNAS , 19 agosto 2019, DOI : 10.1073/pnas.1911137116 .
- ^ ( FR ) J.-J. Hublin, Y. Coppens e P. Picq, Origine et évolution des Néandertaliens , in Aux origines de l'humanité , vol. 1, Fayard , 2001.
- ^ ( FR ) J. Jaubert, Chasseurs et artisans du Moustérien , in La Maison des Roches , 1999. .
- ^ Anche i Neandertal seppellivano i loro morti , su lescienze.it , 9 dicembre 2020. URL consultato il dicembre 2020 .
- ^ ( EN ) JJ Hublin, C. Barroso Ruiz, P. Medina Lara, M. Fontugne e J.-L. Reyss, The Mousterian site of Zafarraya (Andalucia, Spain): dating and implications on the palaeolithic peopling processes of Western Europe , in Comptes rendus de l'Académie des sciences , 321 (IIa), 1995, pp. 931-937.
- ^ ( EN ) J.-J Hublin e E. Trinkaus, The Mousterian human remains from Zafarraya (Andalucia, Spain) , in American Journal of Physical Anthropology , supplemento 26, 1998, pp. 122-123.
- ^ ( EN ) FH Smith, E. Trinkaus, PB Pettitt, I. Karavanić e M. Paunovic, Direct radiocarbon dates for Vindija G1 and Velika Pećina Late Pleistocene hominid remains , in PNAS , vol. 96, n. 22, 26 ottobre 1999, pp. 12281-12286.
- ^ T. Higham, CB Ramsey, I. Karavanić, FH Smith e E. Trinkaus, Revised direct radiocarbon dating of the Vindija G1 Upper Paleolithic Neanderthals , in PNAS , vol. 103, n. 3, 17 gennaio 2006, pp. 553-557.
- ^ ( EN ) C. Finlayson et al. , Late survival of Neanderthals at the southernmost extreme of Europe , in Nature , 13 settembre 2006.
- ^ ( EN ) D. Brill, Neanderthal's last stand , in [email protected] , 13 settembre 2006.
- ^ Johannes Krause et al. The complete mitochondrial DNA genome of an unknown hominin from southern Siberia , Nature advance online publication 24 March 2010 | doi:10.1038/nature08976
- ^ Red-Heads and Neanderthals , su gktgazette.com , maggio 2001. URL consultato il 28 ottobre 2005 (archiviato dall' url originale il 18 ottobre 2005) .
- ^ Nicole's hair secrets , su arts.telegraph.co.uk , 10 febbraio 2002. URL consultato il 2 novembre 2005 (archiviato dall' url originale il 24 febbraio 2006) .
- ^ Roxanne Khamsi, Neanderthal DNA illuminates split with humans , su newscientist.com , 11 ottobre 2006. URL consultato il 28 agosto 2014 .
- ^ Michael Shermer, Our Neandertal Brethren: Why They Were Not a Separate Species , su scientificamerican.com , 1º luglio 2010. URL consultato il 28 agosto 2014 .
- ^ ( EN ) Neanderthal bone gives DNA clues , su cnn.com .
- ^ ( EN ) Scientists decode Neanderthal genes , su msnbc.msn.com .
- ^ ( EN ) Humans and Neanderthals interbred , su cosmosmagazine.com . URL consultato il 25 maggio 2007 (archiviato dall' url originale il 22 febbraio 2009) .
- ^ Elizabeth Pennisi, Ancient DNA: No Sex Please, we're Neanderthals , in Science , vol. 316, n. 5827, 18 maggio 2007, p. 967, DOI : 10.1126/science.316.5827.967a . URL consultato il 18 maggio 2007 .
- ^ Sriram Sankararaman et al. , The date of interbreeding between Neanderthals and modern humans , in Plos genetics , vol. 8, n. 10, 1º ottobre 2012, DOI : 10.1371/journal.pgen.1002947 . URL consultato il 26 novembre 2012 .
- ^ Tom Higham, Christopher Bronk Ramsey, Ivor Karavanic, Fred H. Smith e Erik Trinkaus, Revised direct radiocarbon dating of the Vindija G1 Upper Paleolithic Neanderthals , in PNAS , vol. 103, n. 3, 17 gennaio 2006, pp. 553-557.
- ^ ( EN ) New Machine Sheds Light on DNA of Neanderthals , su nytimes.com , New York Times .
- ^ ( EN ) Jason Shogren, R. Horan e E. Bulte, How Trade Saved Humanity from Biological Exclusion: An Economic Theory of Neanderthal Extinction. , in Journal of Economic Behavior and Organization , vol. 58, 2005, pp. 1-29.
- ^ Steven L. Kuhn e Mary C. Stiner, What's a Mother to Do? The Division of Labor among Neanderthals and Modern Humans in Eurasia , in Current Anthropology , vol. 47, n. 6, dicembre 2006.
- ^ Cfr. JET Channell, L. Vigliotti, "The role of geomagnetic field intensity in late Quaternary evolution of humans and large mammals", Reviews of Geophysics , Vol. 57, 29 May 2019 [1] , [2] .
- ^ ( EN ) Neanderthal exploitation of marine mammals in Gibraltar , in PNAS , n. 105, 22 settembre 2008, pp. 14241-14242, DOI : 10.1073/pnas.0807931105 .
- ^ ( EN ) Neanderthal diet like early modern human's , su nhm.ac.uk .
- ^ MP. Richards, PB. Pettitt; E. Trinkaus; FH. Smith; M. Paunović; I. Karavanić, Neanderthal diet at Vindija and Neanderthal predation: the evidence from stable isotopes. , in Proc Natl Acad Sci USA , vol. 97, n. 13, Jun 2000, pp. 7663-6, DOI : 10.1073/pnas.120178997 , PMID 10852955 .
- ^ <Bo Live, magazine dell'Università di Padova, 5 marzo 2020
- ^ Da William L. Straus, Jr. e AJE Cave, Quarterly Review of Biology , vol. 32, n. 4, dicembre 1957.
Citato in ( EN ) Oxford Dictionary of Modern Quotations , Oxford University Press, 2007, ISBN 0-19-920895-6 . - ^ A. Leroi-Gourhan 1965, pp. 56-57, figg. 223-225; J. Kozlowski 1992, p. 37 e ss. e figg. 21-25; M. Lorblanchet 1999, p. 203 e ss.
- ^ ( FR ) Bernard Heuvelmans e Boris F. Porchnev, L'homme de Néanderthal est toujours vivant , in Paris: Plon , 1974.
- ^ G. Giacobini, L. Umley e A. Dem, Les fossiles humains de la Caverna delle Fate (Finale, Ligurie Italienne) et la definition des caracteres Néandertaliens au debut du Würm , in L'Homme de Néandertal , 3: L'Anatomie, Liegi, M. Otte ed E.Trinkaus, 1988, pp. 53-65..
- ^ Museo Archeologico del Finale , su museoarcheofinale.it .
- ^ G. Giacobini e F. D'Errico, La fauna. In Atti Tavola Rotonda Internazionale sull'interpretazione delle impronte umane della Grotta di Toirano , in Rivista di Studi Liguri , vol. 51, n. 4, 1985, pp. 345-352.
- ^ G. Villa, G. Giacobini e Perettoc., thun hohenstein u. 2001, Neanderthal teeth from the Mousterian levels of the Riparo Tagliente (Verona – NE Italy) , in Atti del XIII Congresso degli Antropologi Italiani , Roma, pp. 45-49. . Z
- ^ Grotta di Fumane
- ^ ( FR ) S. Cotrozzi, Francesco Mallegni e AM Radmilli, Fémur d'un enfant néandertalien dans la Buca del Tasso à Metato, Alpi Apuane (Italie) , in L'Anthropologie , n. 89, Parigi, 1985, pp. 111-116.
- ^ G. Stefanini, Condizioni fisiche e stratigrafiche della Buca del Tasso , in Archivio per l'Antropologia e l'Etnologia , LII, 1922, pp. 4-9.
- ^ "Neanderthal, dalla Grotta Guattari al Circeo nuove incredibili scoperte" sul sito del Ministero Beni Culturali
- ^ I Paleantropi di Saccopastore di CA Blanc ( PDF ), su quartaer.eu . URL consultato il febbraio 2016 .
- ^ Giampiero Dantoni, I livelli musteriani con strumenti su valva di Callista (Callista)chione (L.) nel Salento
- ^ Roberto Filograna, La grotta del cavallo e la sua preistoria
- ^ Arturo Palma di Cesnola, Il Paleolitico della Puglia .
- ^ ( EN ) Silvana M. Borgognini Tarli, A neanderthal lower molar from Fondo Cattìe (Maglie, Lecce) , in Journal of Human Evolution , vol. 12, n. 8, 1983 (archiviato dall' url originale il 3 maggio 2014) .
- ^ Silvana Borgognini Tarli, Studio di un molare neanderthaliano (Maglie 1) proveniente dal Fondo Cattie (Maglie, Lecce) , in Quaderni; pubblicazione scientifica del Museo di Paleontologia di Maglie , n. 1, 1983.
- ^ Monte Generoso , su montegeneroso.ch . URL consultato il 25 luglio 2011 (archiviato dall' url originale l'8 aprile 2012) .
- ^ Parco Curone , Ana , UbiGreen FondazioneCariplo. Relazione archeologica .
- ^ Grosseto, trovate tracce dell'uomo di Neanderthal: «Viveva nella cava di La Pietra»
- ^ Pagina dedicata a "Topolino All'Età Della Pietra" su INDUCKS , su coa.inducks.org , 21 agosto 2014.
Bibliografia
- ( EN ) Richard E. Green, Anna-Sapfo Malaspinas, Johannes Krause, Adrian W. Briggs, Philip LF Johnson, Caroline Uhler, Matthias Meyer, Jeffrey M. Good, Tomislav Maricic, Udo Stenzel, Kay Prüfer, Michael Siebauer, Hernán A. Burbano, Michael Ronan, Jonathan M. Rothberg, Michael Egholm, Pavao Rudan, Dejana Brajković, Željko Kućan, Ivan Gušić, Mårten Wikström, Liisa Laakkonen, Janet Kelso, Montgomery Slatkin e Svante Pääbo, A Complete Neanderthal Mitochondrial Genome Sequence Determined by High-Throughput Sequencing [ collegamento interrotto ] , in Cell , vol. 134, pp. 416-426.
- ( EN ) Richard E. Green, Johannes Krause, Susan E. Ptak, Adrian W. Briggs, Michael T. Ronan, Jan F. Simons, Lei Du, Michael Egholm, Jonathan M. Rothberg, Maja Paunovic e Svante Pääbo, Analysis of one million base pairs of Neanderthal DNA ( PDF ), in Nature , n. 444, 16 novembre 2006, pp. 330-336.
- AA. VV., I neanderthaliani. Comparsa e scomparsa di una specie , in Systema Nature , vol. 7, Pavia, Edizioni Altravista, 2009.
- Svante Pääbo L'uomo di Neanderthal, Alla ricerca dei genomi perduti , Einaudi , ISBN 978-88-06-20566-9 , 2014.
Voci correlate
Altri progetti
-
 Wikiquote contiene citazioni sull' Homo neanderthalensis
Wikiquote contiene citazioni sull' Homo neanderthalensis -
 Wikimedia Commons contiene immagini o altri file sull' Homo neanderthalensis
Wikimedia Commons contiene immagini o altri file sull' Homo neanderthalensis -
 Wikispecies contiene informazioni sull' Homo neanderthalensis
Wikispecies contiene informazioni sull' Homo neanderthalensis
Collegamenti esterni
- ( EN ) Homo neanderthalensis , su Enciclopedia Britannica , Encyclopædia Britannica, Inc.
- ( EN ) Homo neanderthalensis , su Fossilworks.org .
- Il Neanderthal non era 'inferiore' al Sapiens , su repubblica.it , la Repubblica , 27 agosto 2008.
- Il Neaderthal si ornava di piume , su repubblica.it , la Repubblica , 25 febbraio 2011.
| Controllo di autorità | Thesaurus BNCF 32365 · LCCN ( EN ) sh85090493 · GND ( DE ) 4171356-4 · BNF ( FR ) cb12474629x (data) · BNE ( ES ) XX540103 (data) · NDL ( EN , JA ) 00568063 |
|---|