Ribozom

Ribozomii sunt complexe macromoleculare, cufundate în citoplasmă sau ancorate la reticulul endoplasmatic dur sau conținute în alte organite ( mitocondrii și cloroplaste ), responsabile de sinteza proteinelor . Funcția lor este de a citi informațiile conținute în lanțul de ARN messenger (ARN-m).
Au fost scoase în 1953 la microscopul electronic de către biologul român George Emil Palade [1] , descoperirea care i-a adus premiul Nobel . În schimb, termenul de ribozom a fost propus în 1958 de Richard B. Roberts [2] .
Structura

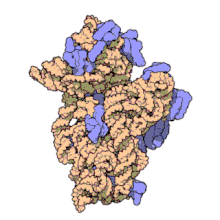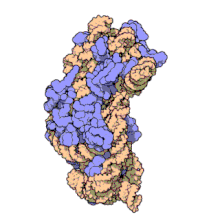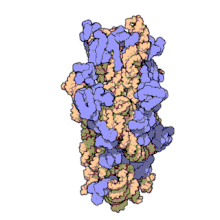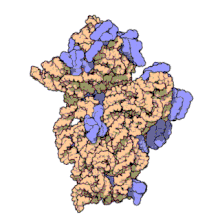
Ribozomii sunt sintetizați în nucleol. Sunt formate din patru molecule de ARN ribozomal și proteine care se asociază pentru a forma două subunități de dimensiuni diferite. Ribozomii bacteriilor , archaea și eucariotele diferă semnificativ între ele atât pentru structură, cât și pentru secvențele ARN .
Un ribozom bacterian are o masă de aproximativ 2700 kDa , un diametru de aproximativ 20 nm și un coeficient de sedimentare de 70 S.
Poate fi împărțit în două părți sau subunități, una mai mare și una mai mică:
- o subunitate mare de 50 S având cel puțin 34 de proteine (L1-L34) și două molecule de ARN (23 S și 5 S),
- o mică subunitate 30 S conținând cel puțin 21 de proteine (S1-S21) și un ARN 16 S.
Pe de altă parte, ribozomul celulei eucariote, cu excepția celor conținute în mitocondriile numite mitoribozomi și cloroplaste, care este de 70 S ca în procariote , este mai mare și are o masă moleculară de 4000 kDa, un diametru de 23 nm și un coeficient de sedimentare de 80 S.
De asemenea, este compus din două subunități, mai mari de 60 S și mai mici de 40 S:
- o subunitate majoră este compusă din trei molecule de ARNr , un 28 S, un S până la 5,8, până la un S 5 final și aproximativ 45 de proteine .
- cel mai mic este format dintr-un singur lanț de ARNr 18 S și 33 de proteine.
În general, ajungem la aproximativ 80 de proteine.
Moleculele unice de ARNr, cu excepția 5 S, sunt sintetizate în nucleoli ca ARN 45 S. ADN-ul conținut în nucleol este transcris de ARN polimeraza I pornind de la mai multe puncte ale lanțului ADN, în structuri (cistronii) care sunt chemați la „pom de Crăciun”: trunchiul ar fi reprezentat de ADN, ramurile de numeroasele lanțuri de ARNr care sunt transcrise în același timp. ARNr 45 S proaspăt transcris se numește pre-ARNr; ca rezultat al tăierilor, va da naștere la 18 S ARNr pentru subunitatea mică a ribozomului și S 32, care vor fi tăiate în continuare în 28 S și 5,8 S. În pre-ARNr sunt prezente pseudouridina și bazele azotate metilate . Se crede că funcția acestor modificări în bazele ambelor evită tăierea enzimei sau promovează interacțiunile ARN interne ale lanțului sau cu alte molecule.
Aranjament
Aranjamentul lor în interiorul celulei variază în funcție de tipul celulei și este legat de funcția acesteia din urmă:
- dacă celula secretă proteina produsă, numai ea posedă ribozomi atașați la reticulul endoplasmatic, care ocupă cea mai mare parte a citosolului și a membranei nucleare;
- dacă celula stochează aceste proteine, are ribozomi liberi în citoplasmă.
Ribozomii liberi
Ribozomii care sunt liberi în citoplasmă sau care fac parte din mitocondrii sau cloroplaste se numesc așa. În general, acestea sunt atribuite sintezei proteinelor care vor fi eliberate și utilizate în citoplasmă sau în partea interioară a membranei organelor unde sunt prezenți ribozomi și, de asemenea, în peroxizomi . Mai mulți ribozomi sunt capabili să se unească folosind legătura ca o aceeași moleculă de ARN mesager, prin care se deplasează de la codonul inițial până la cel final în direcția 3 ', pentru a forma un poliribosom (sau polizom ).
Ribozomi legați de membrane
Acestea se găsesc legate de membranele care constituie nucleul celular sau reticulul endoplasmatic dur. Aceștia sunt responsabili să sintetizeze și să elibereze proteine în membranele acestor structuri, unde pot fi transportate la destinația lor finală, care pot fi intra- sau extracelulare, caz în care sunt adesea expulzați prin exocitoză . Ribozomii aderați la membrane sunt o secțiune a setului de ribozomi liberi, o secțiune care este apoi recrutată imediat ce încep să sintetizeze proteinele specifice.
Procesul de traducere
Cele două subunități ale ribozomului se unesc între ele și lucrează împreună pentru a traduce un ARN mesager într-un lanț polipeptidic în timpul sintezei proteinelor. Cea mai importantă parte a ribozomului, în acest sens, sunt moleculele de ARN ribozomal care sunt capabile de ele însele, deși încet, să opereze procesul de translație, comportându-se ca un ribozim . Acest lucru sugerează că componenta proteică acționează în așa fel încât să sporească și să accelereze acțiunea ARN-ului ribozomal.
Înainte ca subunitățile să se unească pentru a începe translatarea moleculei de ARNm, este nevoie ca complexul de pre-inițiere să se lege de subunitatea inferioară; în acest moment ribozomul se poate conecta la molecula de ARN mesager și, odată ce subunitatea minoră recunoaște tripletul de inițiere, factorii de pre-inițiere se detașează, cele două subunități se conectează și ribozomul poate începe să sintetizeze proteina. Subunitatea superioară este compusă din trei situri, în ordine: situsul A ( aminoacid ), P ( peptididico ) și situsul E („ieșire”, ieșire). Amintiți-vă că subunitatea inferioară a pregătit doar un loc de atașament pentru ARNm .
Primul ARN de transfer (ARNt) este inserat în situsul A, în acest moment ajunge un al doilea ARNt care va fi poziționat în situsul P. Acum, ribozomul curge de un triplet, aducând aminoacizii traduși din situsul A, pentru a se lega la situsul P Tripletul codificat de P trece acum la site-ul E, unde va fi lansat. În concluzie:
- Site-ul A este site-ul care așteaptă sosirea de noi triplete care vor fi codificate;
- Site-ul P menține fermă secvența polipeptidică, eliberând-o numai atunci când site-ul A a tradus un aminoacid suplimentar;
Ipoteza semnalului
Ribozomii liberi și legați de membrană sintetizează două clase diferite de proteine. Cum poate celula să facă o astfel de distincție? Ipoteza semnalului, dezvoltată în 1971, afirmă că informațiile care permit acest proces să funcționeze sunt conținute în secțiunea N-terminală a lanțului polipeptidic, care este prima care iese din ribozom. Ulterior acestei ipoteze s-au adăugat câteva date justificative care au permis să se ajungă la următorul model.
Catenă ARN mesager care urmează a fi tradusă ajunge la ribozom care în acel moment este liber și conținut în citoplasmă. Odată ce traducerea a început, o secvență de 6-20 aminoacizi nepolari este conținută în partea N-terminală (deși în unele polipeptide este mai internă). Această secvență este recunoscută de o particulă de recunoaștere a semnalului (SRP), o structură formată dintr-o moleculă de ARN (ARN 7SL) și 6 lanțuri polipeptidice, care se leagă de aceasta și blochează plierea prematură și continuarea sintezei proteinelor. Acest lucru face ca ribozomul să se deplaseze către membranele reticulului endoplasmatic dur, în loc să finalizeze traducerea proteinei care altfel ar fi eliberată în citoplasmă. Particula de recunoaștere a semnalului legat acționează ca un ligand care se leagă de doi receptori găsiți pe membrana citoplasmatică a reticulului endoplasmatic, unul pentru SRP și celălalt pentru ribozom. Pentru a crea legătura dintre SRP și receptorul său, o moleculă de guanozin trifosfat (GTP) trebuie să se lege de SRP. Ulterior, SRP este deconectat de la secvența semnal și apoi de ribozom, pentru hidroliza GTP la difuzat de guanozină (PIB), în timp ce secvența semnal se leagă de o secțiune a unui canal proteic transmembranar. După detașarea SRP, ribozomul reia sinteza proteinelor, dar lanțul polipeptidic este translocat prin canal și intră în reticulul endoplasmatic (translocație). Ulterior, secvența semnal este tăiată de o enzimă proteolitică, peptidaza semnalului, iar restul proteinei este supus proceselor de repliere adecvate și formării de legături disulfidice.
Notă
Bibliografie
- Easter G., Abbate G., C. Forni, General Botany and Plant Diversity, Ed. Piccin, 2007
Elemente conexe
Alte proiecte
-
 Wikționarul conține intrarea în dicționar „ ribozom ”
Wikționarul conține intrarea în dicționar „ ribozom ” -
 Wikimedia Commons conține imagini sau alte fișiere pe ribozom
Wikimedia Commons conține imagini sau alte fișiere pe ribozom
linkuri externe
- Ribozom pe Treccani.it - enciclopedii online, Institutul Enciclopediei Italiene .
- (EN) Ribosome on Encyclopedia Britannica , Encyclopædia Britannica, Inc.
- (RO) IUPAC Gold Book, „ribozomi” pe goldbook.iupac.org.
- (EN) Nature Structural & Molecular Biology: SRP întâlnește ribozomul , pe nature.com.
- Ribozom în Treccani.it - enciclopedii online, Institutul italian de enciclopedie.
| Controlul autorității | Tezaur BNCF 58308 · LCCN (EN) sh85113856 · GND (DE) 4127731-4 · BNF (FR) cb12254742n (data) · NDL (EN, JA) 00.569.445 |
|---|