Briofitele
| Briofiți | |
|---|---|
 | |
| Clasificare științifică | |
| Domeniu | Eukaryota |
| Regatul | Plantae |
| Divizia | Briofitele Schimp. |
| Clase | |
Briofitele ( Schimp. ) Sunt un grup de embriofite fără țesut vascular . Acesta este cel mai divers grup de plante non-vasculare, cu aproximativ 24.000 de specii [1] . Sunt cunoscute sub denumirea de mușchi.
Briofitele sunt caracterizate în general de țesuturi vasculare ne lignificate: absorbția și transportul apei și substanțelor dizolvate este în general necesară pentru acțiunea capilară și afectează întreaga suprafață a plantei. La unele specii sunt prezente corzile celulelor cu funcție conductivă: aceste structuri nu sunt foarte eficiente și nu au funcție de susținere [2] . Lipsa unui sistem de conducere împiedică dezvoltarea în înălțime: briofitele sunt plante de dimensiuni destul de mici și în general creștere orizontală.
Adaptările speciale dezvoltate de briofite atât la nivel vegetativ, cât și la nivel reproductiv sunt de mare interes pentru a înțelege studiul evoluției plantelor terestre [3] . Briofitele, care par să provină din algele verzi, sunt împărțite în trei grupe: Anthocerotopsida (sau antoceri), Hepatopsida (sau ficatul) și briopsida (sau mușchiul) [4] . Studiile moleculare au arătat că un grup de briofite sunt monofiletice [5] .

Anthoceros agrestis

Antheridiofori Marchantia


Sphagnum palustre
Caracteristici

Se deosebesc de plantele vasculare, deoarece nu au structuri vasculare complet diferențiate și lignificate . Absorbția și transportul apei și substanțelor dizolvate se realizează în mare parte prin capilaritate și afectează întreaga suprafață a plantei. La unele specii sunt prezente structurile ( hidroizi și leptoidi ) analogi țesuturilor vasculare xilem și floem, dar niciodată lignificate (deci fără funcție de susținere) și mai puțin eficiente în transportul pe apă.
Ciclul ontogenetic este aplodiplonte cu prevalență gametofitică . Sporofitul nu este autonom, dar trăiește în gametofit cheltuiește fotosintetic . Dintr-un spor haploid unicelular ( meiospora ) se dezvoltă protonema, cu dimensiuni diferite în funcție de specie. Pe protoneme, haploide, sunt formate din mugurii din care se dezvoltă până la maturitate gametofiți care poartă gametangia (antheridia și arhegonia) care conțin respectiv gameti feminin și masculin. Datorită prezenței apei, gametii masculi ajung la gametul feminin rămas în interiorul archegoniului. Acesta este locul în care gamy rezultă în dezvoltarea ' embrionului diploid . Din embrion, conținut și hrănit în archegonium (care face parte din gametofit), sporofitul adult nu se dezvoltă în mod normal. Pe sporofit, în interiorul sporangiului, apare meioza , cu înjumătățirea numărului de cromozomi și apare meiosporul.

Briofitele au, de asemenea, un început de cutinizare pentru a limita transpirația ( stomatele ).
Acestea sunt importante pentru colonizarea unor medii noi, foarte rezistente (reviviscente). Cele mai cunoscute forme sunt mușchii.
Lipsa țesuturilor de transport limitează mărimea briofitelor: înălțimea este mai mică la majoritatea speciilor la 6 centimetri. Cel mai mare briofit cunoscut este un mușchi din Australia și Noua Zeelandă : dawsonia superbă . Această specie atinge 75 de centimetri înălțime. Pe de altă parte, cele mai mici briofite au mai puțin de un milimetru.
Ecologie și distribuție
O cerință fundamentală pentru creșterea briofitelor este, prin urmare, prezența apei sau, în orice caz, a condițiilor umede pentru perioade lungi. Briofitele au nevoie, de asemenea, de un mediu de creștere relativ stabil, de o expunere adecvată la soare și de un interval de temperatură favorabil pentru creștere. Multe briofite reușesc să supraviețuiască perioadelor de secetă sau îngheț printr-un mecanism care nu este încă clar înțeles.
Mediul de creștere preferat este solul, unde briofitele cresc adesea pe rămășițele generațiilor anterioare sau alte plante; alte substraturi comune sunt roci goale, medii acvatice și canapele. Unele briofite cresc pe rămășițe animale în descompunere, la suprafața frunzelor, în săruri, la baza pietricelelor de cuarț sau în medii bogate în cupru. Altele, cum ar fi speciile din genul funaria , sunt capabile să crească pe terenul ars ca urmare a incendiului.
Modelele de distribuție ale briofitelor sunt similare cu cele ale plantelor vasculare. Unele specii sunt cosmopolite, multe cresc în zone limitate. Briofitele sunt distribuite peste tot în lume: singurele excepții sunt regiunile aride, unde totuși pot crește în micro-medii umede și medii cu apă sărată. Unele briofite trăiesc în zona intertidală . Briofitele sunt mai abundente în regiunile cu un climat constant umed în regiunile tropicale și subtropicale; în regiunile arctice sau alpine se pot forma covoare foarte extinse care influențează tipul de vegetație și dinamica acesteia.
Reproducerea sexuală și ciclul de viață

Faza dominantă a ciclului de viață al briofitelor este reprezentată de gametofit . Aceasta poate realiza fotosinteza și, prin urmare, este independentă din punct de vedere nutrițional. Sporofitul poate fi în schimb fotosintetic sau nu: în orice caz este întotdeauna dependent de gametofit și rămâne fixat stabil de acesta.
Sporofitul produce pentru meioza sporilor haploid unicelular ( meiospor ). Acești spori pot rămâne latenți mult timp înainte de a aștepta condiții optime de germinare. În momentul germinării din spori dezvoltă o structură filamentoasă și protonema menționată slab diferențiată. Ulterior, gametofitul adult se dezvoltă din protonemă. Pe gametofit există structuri asemănătoare părului uni- sau multicelulare, numite rizoizi. Funcția rizoizilor este de a ancora gametofilul pe substrat. La multe specii gametofitul este micorrizzati [6] .
Gametofitul diferă de organele sexuale sau gametangia . Femela se spune gametangio archegonium, în timp ce masculul este numit anteridio . Există briofite omotalliche sau monoice și briofite eterotalliche sau dioice . În primii, gametofiții poartă arhegonia și antheridia asupra aceluiași individ; în acesta din urmă există indivizi care poartă arhegoniul și alții care poartă antheridia.
O singură celulă de ou este conținută în archegonium. Arhegoniul are forma unui balon, cu un gât alungit și o bază menționată în pântece bombată : în această parte se păstrează celula de ou.
Nell'anteridio au produs un număr mare de gameti masculi cu doi flageli. La maturitate, gametii masculi sunt eliberați și, folosind un văl de apă, ajung în archegoniu, atrași de stimuli de natură chimică. Necesitatea prezenței unui văl de apă pentru a permite supraviețuirea gametului masculin în afara antheridiumului și în timpul călătoriei sale până la archegonium este unul dintre factorii care limitează răspândirea briofitelor în medii umede. Odată ajunși la archegoniu, gametii masculi pătrund în interiorul acestuia prin gât: intrarea lor determină producerea unei substanțe gelatinoase. Această substanță permite sosirea gametului masculin în abdomen, unde are loc fertilizarea .
Zigotul rezultat din fertilizare și embrion ulterior crește nell'archegonio. Sporofitul adult se dezvoltă apoi din embrion. Aceasta este în general formată dintr-un picior, o mătase și o capsulă.
Piciorul este o structură inserată în burtica arhegoniului: funcția sa este de a permite trecerea nutrienților de la gametofit la sporofit. Mătasea este un peduncul neramificat care permite sporofitului să se dezvolte în înălțime. Capsula este constituită în esență din sporangiu : în interior se formează spori haploizi. La mușchi, capsula poate conține câteva milioane de spori. La maturitate, capsula se deschide cu mecanisme de control, în general higroscopice, eliberând sporii de la care va începe următoarea generație.
Sporofitul unor specii de briofite nu este cunoscut: sunt probabil specii care și-au pierdut capacitatea de reproducere sexuală.
Limite
La briofiți, fiecare gametofit haploid este: în fiecare an se formează apoi gameți cu genetici întotdeauna identici. La briofite omotalliche este, de asemenea, foarte probabil ca gametii masculi să fertilizeze arhegonia, același gametofit generând sporofiți homozigoti . Aceste condiții au ca rezultat o limitare majoră a posibilității de a dezvolta noi personaje: în aceste cazuri, singura posibilitate de apariție a noilor personaje este legată de mutații spontane.
Reproducere asexuată
Briofitele se pot reproduce și prin asexuare (sau asexuată) pentru fragmentarea talului sau prin pietre prețioase . Acestea din urmă sunt conținute, la unele specii, în structuri speciale în formă de bol mic dispuse pe suprafața gametofitului.
Dovezile fosile
Înregistrările fosile atribuite briofitelor sunt foarte slabe [7] . Una dintre dificultățile majore în recunoașterea briofitelor în fosile este lipsa conservării sporofitelor și similitudinea gametofitelor cu unele plante vasculare. Primele dovezi fosile din roci datorate briofitelor provin din „ Argentina și datează din prima parte a„ Ordovicianului (488-444 Ma). Este vorba despre elemente similare spori legați de ficat [8] . Aceste mărturii sunt considerate de unii cercetători drept cele mai vechi dovezi ale prezenței plantelor terestre [9] . Primele dovezi ale mușchilor datează din Permian (299-251 Ma). Majoritatea fosilelor de briofite sunt atribuite „ Eocenului (55,8-33,9 Ma). Acestea sunt plante conservate în chihlimbar cu structuri foarte asemănătoare cu cele actuale, astfel încât să permită recunoașterea multor specii conservate. Același lucru se aplică fosilelor de briofite datând din paleogen și „ neogenă ”: în acest caz este posibil să se recunoască cele mai multe specii [10] . Prin urmare, liniile evolutive din briofite s-au născut și s-au stabilizat în timpuri foarte vechi.
Clasificare
Sistematică tradițională
Clasificarea tradițională include grupul de briofite din divizia Bryophyta, care este împărțit în trei clase: briopsida, Marcanthiopsida și Anthocerotopsida.
- Mușchii (Bryopsida), cu peste 10.000 de specii, sunt cel mai răspândit grup de briofite. Aceștia fac cormoide cu tulpină și frunze adesea multistratificate.
- Eubrydae: Capsulă cu inel, prezența ligninei în peristom (nell'opercoli), deosebit de răspândită.
- Andreaeobrydae: Originea gametofitică a mătăsii
- Sphagnobrydae: Sphagnum, practic nemuritor (partea inferioară se degradează și apoi dă secole de turbă și carbonat-fosil); protonema este laminară, nu există peristom.
- Ficatele (Marchantiopsida), includ aproximativ 7.000 de specii, cu o distribuție cosmopolită cu o preferință pentru mediile umede. Le lipsește protonemele, au rizoizi unicelulari, capsulă mai simplă fără columelă, printre meiosporii care dezvoltă elateri pentru a favoriza dispersia sporilor.
- Antocerotele (Anthocerotopsida) includ o sută de specii. Au un gametifitico ridat format din folii subțiri, gametofite cu stomate și cloroplast, cu picioare care străpung sporofitul, nu complet autonome. Din care se crede că au apărut ferigi cu Rhyniophyte dispărute în Silurian. Deci, ele sunt legătura cu plantele vasculare superioare, deoarece ne gândim la ipoteza că sporofitul său ar fi putut trăi independent de gamentofit și că inversarea ciclului haplodiplon a avut loc în favoarea sporofitului pe gametofit, ca în majoritatea plantelor vasculare superioare plecând de la ferigi.
Filogenia și clasificarea modernă
Studii recente privind embriofitul au arătat că grupul de briofite nu este monofiletic, ci parafiletic , deoarece include strămoșul comun și doar unele dintre grupurile derivate din acesta. Polisporangiatul , din care coboară toate plantele vasculare, este de fapt exclus. Acest lucru a dus la ridicarea fiecărei clase individuale la rangul de diviziune:
- Briofite care conțin mușchi și alge
- Marchantiophyta , aparțin ficatului
- Hornwort , cu hornworts
Există două ipoteze probabile despre istoria evoluției plantelor terestre:
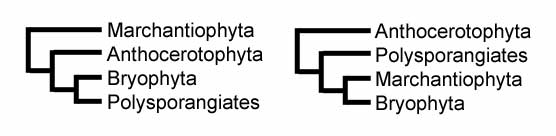
Conform primei ipoteze, ficatele au fost primele care diverg, urmate de antocerote, în timp ce mușchii sunt relativ mai apropiați de grupul polisporangiatului. În cea de-a doua schemă, pe de altă parte, antocerotele diverg mai întâi, urmate de plantele vasculare, în timp ce mușchii și ficatele sunt mai apropiate filogenetic.
Divizia Bryophyta (mușchi)
- Clasa Hepatopsida (ficat)
- Ordinul Calobryales
- Ordinul Metzgeriales
- Ordinul Jungermanniales
- Ordinul sferocarpale
- Ordinul Monocleales
- Ordinul Marchantiales
- Clasa Anthocerotopsida (sau antoceri)
- Ordinul Anthocerotales
- Clasa briopsida (mușchi)
- Subclasa Andreaeidae
- Subclasa Sphagnidae
- Subclasa Tetraphidae
- Subclasa Polytrichidae
- Subclasa Buxbaumiidae
- Subclasa Bryidae
- Subclasa Archidiidae
Briofiți în Italia
Conform celei mai recente liste de verificare [11] [12] Flora italiană include 1130 briofite. Bogăția deosebită a florei este evidentă atunci când considerați că în Europa sunt raportate briofite în 1690 [13] . Bogăția deosebită a florei briologice italiene se datorează poziției sale geografice și variabilității mediilor sale.
Există (sau a existat) reliquato al unei specii fosile , prezent până la ultima glaciație : Paludella squarrosa , în mlaștina Santa Caterina Valfurva . Până la începutul anilor optzeci a fost prezent pe o suprafață de câțiva metri pătrați, în mijlocul sfagnului . De atunci nu mai știi, dar sectorul construcțiilor turistice, care a avansat ocupând mlaștina, ar fi putut să-și fi anulat definitiv existența. [14]
Notă
- ^ Aleffi M., 2007: Briofiți. În: C. Blasi, Boitani L., Post S., Manes F. & M. Marchetti, Biodiversity in Italy. Contribuția la Strategia Națională pentru Biodiversitate: 162-171.
- ^ Aceste corzi sunt formate din celule lipsite de maturitate a protoplasmei și înconjurate de alte celule cu nuclee degenerate și perforate ale pereților transversali.
- ^ Shaw J. și K. Renzaglia, 2004: Filogenia și diversificarea briofitelor. American Journal of Botany 91 (10): 1557-1581. Pentru inspecție la: http://www.amjbot.org/content/91/10/1557.full.pdf+html Depus la 8 martie 2012 la Internet Archive . (verificat la 2 decembrie 2011).
- ^ Aleffi M., op. cit.
- ^ Nishiyama T., Wolf PG, Kugita M., Sinclair RB, Sugita M., Sugiura C., Wakasugi T., Yamada K., K. Yoshinaga, Yamaguchi K., Ueda k. & H. Mitsuyasu, 2004: Cloroplastul de filogenie indică faptul că sunt briofitele monofiletice. Biologie moleculară și evoluție 21 (10): 1813-1819. Pentru inspecție la: https://mbe.oxfordjournals.org/content/21/10/1813.full.pdf+html (a avut loc la 2 decembrie 2011).
- ^ Pentru a aprofunda relația ciuperci / briofite vezi: S. Pressel, Bidartondo MI, Ligrone R. & Duckett JG, 2009: Simbioze fungice în briofite: noi perspective asupra secolului XXI. Phytotaxa 9: 238-253. Pentru inspecție la: http://www.mapress.com/phytotaxa/content/2010/f/pt00009p253.pdf (Verificat 4 decembrie 2011).
- ^ Smoot EL & TN Tayolor, 1986: plante fosile conservate structural din Antarctica: II. Un mușchi permian din Munții Transantartici. American Journal of Botany 73 (12): 1683-1691. Pentru inspecție la: Copie arhivată (PDF) pe paleobotany.bio.ku.edu. Adus pe 3 decembrie 2011 (depus de 'url original 18 iunie 2010). (Verificat 4 decembrie 2011).
- ^ Archangelsky S. & L. Villar de Seone, 1996 Palynological Estudios de la formacion Baquero (Cretaceous), provincia Santa Cruz, Argentina. Ameghiniana 35: 7-19.
- ^ Vezi: http://www.uky.edu/KGS/education/geologictimescale.pdf (Verificat la 3 decembrie 2011).
- ^ Vezi, de exemplu: RH Zander, 2009: analiza volutionay a cinci familii de briofite folosind fosile virtuale. Anales del Jardin Botanico de Madrid 66 (2): 263-277. Disponibil la: http://redalyc.uaemex.mx/redalyc/pdf/556/55612913005.pdf [ link rupt ] (Verificat la 3 decembrie 2011).
- ^ Aleffi M. și R. Schumacher, 1995: Listă de verificare și listă roșie a meritelor vii (marchantiophyta) și hornworts (Anthrocerotophyta) din Italia. Flora Mediterranea 5: 73-161.
- ^ Cortini Pedrotti C., 2001: Nouă listă de verificare a mușchilor din Italia. Flora Mediterranea 11: 23-107. Pentru inspecție la: http://www.herbmedit.org/flora/11-023.pdf (a avut loc la 2 decembrie 2011).
- ^ Aleffi M., 2007: op.cit.
- ^ Raportarea Paludella squarrosa (PDF), pe museocivico.rovereto.tn.it.
Bibliografie
- AA. VV., 2003: Număr special: Briofit. Fremontia 31 (3): 1-44. Pentru inspecție la: https://web.archive.org/web/20061224155818/http://www.uam.es/informacion/asociaciones/SEB/divulgacion/Fremontia_Vol31-No3.pdf (a avut loc la 2 decembrie 2011).
- Glyme JM, 2006: Bryophyte Ecology. Volumul 1. Ecologie fiziologică. Sponsorizat de Michigan Technological University; Societatea Botanică din America , Asociația Internațională a Briologilor. Publicat online: http://www.bryoecol.mtu.edu/ (a avut loc la 2 decembrie 2011).
- WH Purves, D. Sadawa, Orians GH & HC Heller, 2001: Biologie. Evoluția diversității. Zanichelli, Bologna. ISBN 88-08-09664-5 .
- Konrat von M., Shaw & KS Renzaglia AJ (eds), 2010: Briofite: Cele mai apropiate rude vii ale primelor plante terestre. Phytotaxa 9: 1-278. Pentru inspecție la: http://www.mapress.com/phytotaxa/content/2010/pt00009.htm (Verificat 4 decembrie 2011).
Elemente conexe
Alte proiecte
-
 Wikimedia Commons conține imagini sau alte fișiere pe briofit
Wikimedia Commons conține imagini sau alte fișiere pe briofit
linkuri externe
- (EN) Bryophyta , of Encyclopedia Britannica , Encyclopædia Britannica, Inc.
- (EN) Bryophyta , pe Fossilworks.org.
- Repertoriu al florei protejate italiene pe minambiente.it (depus de „Adresa URL originală 9 iulie 2011).
| Controlul autorității | Tesauro BNCF 4230 · LCCN (EN) sh85017380 · GND (DE) 4040211-3 · BNF (FR) cb11964814z (dată) · NDL (EN, JA) 00.566.261 |
|---|