Dromaeosauridae
| Dromaeosauridae | |
|---|---|
 | |
| Starea de conservare | |
Fosil | |
| Clasificare științifică | |
| Domeniu | Eukaryota |
| Regatul | Animalia |
| Sub-regat | Eumetazoa |
| Superphylum | Deuterostomie |
| Phylum | Chordata |
| Subfilum | Vertebrate |
| Infraphylum | Gnathostomata |
| Superclasă | Tetrapoda |
| Clasă | Reptilia |
| Subclasă | Diapsid |
| Infraclasă | Arhosauromorpha |
| Superordine | Dinozauria |
| Ordin | Saurischia |
| Subordine | Theropoda |
| Infraordon | Tetanurae |
| Superfamilie | Celurosaurie |
| Familie | † Dromaeosauridae Colbert și Russell, 1969 |
Dromaeosauridae este o familie de dinozauri teropode cu pene care au prosperat în perioada Cretacicului . Numele Dromaeosauridae înseamnă „șopârle care aleargă”, din greaca δρομευς (alergător) și σαυρος (șopârlă). Membrii săi sunt adesea denumiți rapitori în medii informale, [1] un termen popularizat de filmul Jurassic Park . Anumite genuri includ sufixul raptor în numele lor pentru a sublinia trăsăturile lor asemănătoare păsărilor , deoarece termenul în engleză (de origine latină ) este adesea folosit în referință la păsările de pradă.
Fosile dromeosauride au fost găsite în America de Nord , Europa , Africa , Japonia , China , Mongolia , Madagascar , Argentina și Antarctica . [2] Au apărut pentru prima dată în stadiul batonian al jurasicului (acum aproximativ 167 milioane de ani) și au supraviețuit până la stadiul Maastrichtian al Cretacicului superior (acum aproximativ 65 de milioane de ani), existând de peste o sută de milioane de ani până la Cretacic- Extincția în masă a paleocenului . Prezența dromeosauridelor în timpul jurasicului mediu a fost confirmată de descoperirea dinților fosili izolați, deși nu au fost încă descoperite schelete fosile care datează din acea perioadă. [3]
Descriere
Construcția generală a dromeosauridelor a redeschis teoriile conform cărora cel puțin unii dinozauri erau creaturi active, rapide și strâns legate de păsări . Desenul realizat de Robert T. Bakker pentru monografia lui John Ostrom din 1969 [4], care descrie dromeosauridul Deinonychus în actul alergării, se numără printre cele mai influente reconstrucții paleontologice din istorie. [5] Dromeosauridele posedau un craniu relativ mare, dinți zimți, un bot alungit și ochi orientați spre înainte, indicând astfel viziunea binoculară.[6] La fel ca majoritatea teropodelor, dromeosauridele aveau un gât ușor lung în formă de „S” și toracele lor era relativ îndesat și profund. La fel ca toți ceilalți Maniraptorieni, aceștia posedau membrele anterioare lungi care, la anumite specii, se puteau îndoi strâns de corp și trei degete alungite cu gheare mari. [7] Structura șoldului dromeosauridelor a fost caracterizată de un pubis alungit care a continuat sub baza cozii. Picioarele dromeosauridelor erau echipate cu o gheară mare în formă de seceră pe al doilea deget. Coada era subțire cu vertebre alungite și lipsite de apofiză după a paisprezecea vertebră caudală. [7]
Se știe că unele, și probabil toate, dromeosauridele sunt acoperite de pene, inclusiv pene mari de la nivelul membrelor anterioare și pene de coadă. Această dezvoltare, ipotezată mai întâi în anii 1980 și apoi confirmată în 1999, reprezintă o schimbare importantă în modul în care dromeosauridele au fost ilustrate istoric în artă și film. [8]
Picioare


Ca și alți teropode, dromeosauridele erau bipede. Spre deosebire de majoritatea teropodelor, care mergeau pe trei degete, urmele fosilizate ale diferitelor grupuri de paravieni (inclusiv dromeosauridele) arată că au mers cu degetul al doilea ridicat, cu doar degetele al treilea și al patrulea atingând solul. [9] Al doilea deget de la picior avea o gheară imensă curbată, despre care se crede că este folosită pentru prinderea prăzii și urcarea copacilor. Această gheară a fost deosebit de bine dezvoltată la eudromaeosaurii mari. [10] O specie în special, Balaur bondoc , avea un prim deget de la picioare la fel de bine dezvoltat ca al doilea. Atât primul, cât și al doilea degetele de la picioare au fost ținute retrase de la sol. [11]
Coadă
Dromeosauridele aveau cozi lungi. Majoritatea vertebrelor caudale aveau proeminențe osoase și unele specii aveau tendoane osificate. În studiul său despre Deinonychus , Ostrom a emis ipoteza că aceste trăsături indicau o coadă rigidă, pe care animalul nu o putea deplasa decât la rădăcină. [4] Cu toate acestea, un specimen de Velociraptor a arătat o coadă conservată în formă de „S”, indicând faptul că s-ar putea mișca lateral cu o oarecare flexibilitate. [12]
S-a emis ipoteza că coada a fost folosită ca suport sau ca contrapondere în timpul alergării sau al zborului; [12] în Microraptor , vârful cozii a fost caracterizat printr-un ventilator în formă de diamant, care ar fi putut fi folosit ca cârmă în timpul zborului sau la alunecare. [13]
mărimea
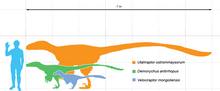
Dromaeosauridae a inclus specii mici și mijlocii; cel mai mic gen era Mahakala , lung de 0,7 metri, în timp ce cel mai mare ( Utahraptor , Achillobator și Dakotaraptor) avea șase metri lungime. [14] [15] Este posibil ca anumite exemplare din Utahraptor să ajungă la 11 metri lungime, deși aceste exemplare merită un studiu suplimentar. [16] Este evident că dimensiunea mare a evoluat convergent cel puțin de două ori în rândul dromeosauridelor; odată printre dromeosaurini (Utahraptor, Achillobator și Dakotaraptor) și din nou printre unenlagiini ( Austroraptor , care măsura cinci metri lungime). Unii dinți izolați găsiți pe Insula Wight par să indice existența unui dromeosaurid de mărimea Utahraptorului ; forma lor indică totuși o relație mai strânsă cu Velociraptorul . [17]
Mahakala a fost cel mai mic și primitiv dromeosaurid găsit. Aceste caracteristici, coroborate cu dimensiunea redusă a altor genuri primitive înrudite, cum ar fi Microraptor și Anchiornis troodontid , indică faptul că strămoșul comun al dromeosauridelor, troodontidelor și păsărilor ar fi putut fi un animal mic, lung de 65 cm și greutate de 600-700 m. grame. [18]
Penaj

„Alegerea este a ta: Utahraptor și știință cu pene sau Raptor și ignoranță scalate”. |
| ( Andrea Cau (2012) [19] ) |
Există multe dovezi că dromeosauridele au fost acoperite de pene. Unele fosile prezintă pene lungi pe membrele anterioare și pe coadă, cu pene pufoase care acoperă corpul. [8] [20] Alte fosile în care nu există urme aparente de penaj arată totuși, pe ulna , papilele osoase care ar fi servit drept ancore pentru penele de zbor . [21] În general, structura și poziționarea penelor în dromeosauridele prezintă similitudini cu cele ale Archeopteryx . [8]
Primul dromeosaurid găsit cu dovezi clare de pene a fost Sinornithosaurus , descoperit în China în 1999. [20] După descoperirea acestuia, au fost găsite și alte exemplare cu corpuri acoperite cu pene, dintre care unele păstrează aripi bine dezvoltate. Microraptorul a fost echipat cu o pereche de aripi secundare pe membrele inferioare. [8] Deși conservarea urmelor de pene este posibilă numai în sedimentele cu granulație fină, s-au găsit dovezi de penaj și în rocile mai aspre, prin prezența papilelor osoase, care sunt prezente și la anumite păsări. Atât Rahonavis , Velociraptor , cât și Dakotaraptor au fost descoperite cu aceste papile, dovedind astfel că au fost pene, deși nu s-au găsit încă urme directe de penaj. Din această cauză, este foarte probabil ca penele să fie prezente și în genurile mari, fără zbor. [21] [22] [23] Deși anumiți oameni de știință din trecut au speculat că dromeosauridele mai mari și-au pierdut penajul, descoperirea papilelor osoase în Velociraptor și Dakotaraptor a fost folosită ca argument pentru a susține că toate dromeosauridele au fost pene. [21] [22] [24]
Paleobiologie
Funcția gheare

Funcția ghearei în formă de seceră pe al doilea deget de la picior este o chestiune de dezbatere. Când John Ostrom a descris-o pentru Deinonychus în 1969, el a speculat că gheara a fost folosită pentru a înghiți prada, cum ar fi dinții canini ai tigrilor cu dinți de sabie . Dawn Adams a sugerat în 1987 că gheara a fost folosită pentru eviscerarea dinozaurilor ceratopside . [25] Această interpretare a fost aplicată tuturor dromeosauridelor. Cu toate acestea, în 2005, s-a sugerat că gheara a fost folosită în primul rând pentru cățărare, dar că ar putea fi folosită și pentru a înjunghia, mai degrabă decât a pradă intestinală. [26]

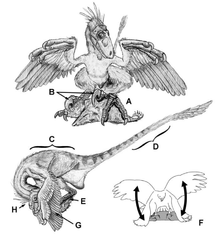
În descrierea lui Deinonychus , Ostrom l-a comparat cu struțul și casuarul , observând cum ambele păsări sunt prevăzute cu gheare letale pe degetul doi. [4] Casuarul are în special o gheară lungă de 125 mm. [28] Ostrom l-a citat pe Gilliard (1958) în descrierea modului în care casuarul este capabil să mutileze și să distrugă un om. [29] Cu toate acestea, studiile efectuate în 1999 și 2003 au constatat că din 241 de atacuri de casuar înregistrate, au pierit doar o persoană și doi câini. Nu există dovezi că victimele au fost dezmembrate sau eviscerate. [30] [31] Casuarele își folosesc de fapt ghearele pentru a se apăra, pentru a ataca amenințări și pentru a-și amenința rivalii. [28] Seriema este o altă pasăre cu o gheară mărită pe al doilea deget; îl folosește pentru dezmembrarea prăzilor mici înainte de a le consuma. [32]
Un studiu realizat în 2009 pentru a testa funcționalitatea ghearelor din spate și din față ale Velociraptorului a arătat paralele structurale cu cele ale bufniței de vultur , ale cărei gheare sunt utilizate în primul rând pentru alpinism. Oamenii de știință implicați în studiu au remarcat că vârful celei de-a doua gheare din spate este ideală pentru găurire și prindere. [33] Comparând ghearele dromeozauridelor cu cele ale anumitor animale de astăzi, s-a observat că nivelul curburii din gheare poate fi folosit ca o indicație a stilului de viață, unde ghearele mai curbate indică o viață mai mult sau mai puțin arbore. Ghearele din spate ale lui Deinonychus au de fapt o curbură de 160 de grade, un nivel tipic al animalelor arborice. Ghearele din față prezintă, de asemenea, o curbură similară. [33]
Paleontologul Peter Mackovicky a comentat acest studiu, concluzionând că gheara trebuie să aibă, de asemenea, o altă funcție, deoarece forma curbă a fost găsită și în genuri mari, cum ar fi Achillobator , prea mare pentru a urca. Macovicky a sugerat ca ghearele să fie folosite pentru a se agăța de flancurile prăzilor mari. [34]
În 2009, Phil Senter a publicat un studiu asupra degetelor de la picioare dromeosauride, demonstrând modul în care flexibilitatea lor este compatibilă cu excavarea cuiburilor de insecte. El a emis ipoteza că genurile mai mici, precum Rahonavis și Buitreraptor , erau suficient de mici pentru a fi insectivore, în timp ce cele mai mari, precum Deinonychus și Neuquenraptor , își vor folosi ghearele pentru a captura prada vertebratelor în cuiburile de insecte. [35]
În 2011, Denver Fowler și colegii săi au sugerat o nouă metodă prin care Deinonychus și celelalte dromeosauride ar captura prada. [27] Acest model, supranumit modelul „restricția prăzii rapitoare” (RPR), propune ca Deinonychus să-și ucidă prada într-un mod similar cu păsările de pradă accipitride ; Deinonychus ar sări peste pradă, ținându-l cu greutatea corporală și agățându-se strâns de ea cu ghearele curbate. La fel ca accipitridele, dromeosauridul ar începe să se hrănească în timp ce prada era încă în viață, până când a sângerat până la moarte. Acest model se bazează în principal pe morfologia și proporțiile picioarelor dromeosauridelor, care sunt comparabile cu cele ale rapitorilor. Fowler a remarcat faptul că membrele inferioare și picioarele majorității dromeosauridelor sunt foarte asemănătoare cu cele ale vulturilor și șoimilor de vrabie , mai ales atunci când se ia în considerare manevrabilitatea a doua gheară mărită și degetul. Cu toate acestea, metatarsul scurt și rezistența la prindere a degetelor erau mai asemănătoare cu cele ale bufnițelor . Metoda RPR ar fi, de asemenea, compatibilă cu alte aspecte anatomice ale dromeozauridelor mari, cum ar fi morfologia mandibulei și a membrelor anterioare. Este posibil ca membrele anterioare să fie acoperite cu pene lungi folosite, împreună cu coada rigidă, pentru a echilibra prădătorul în timp ce strângea prada. Mandibulele sale, despre care se crede că sunt relativ slabe, [36] ar fi putut fi folosite într-o mișcare de ferăstrău , așa cum o face dragonul modern Komodo . [37]
Comportamente sociale

Unele fosile Deinonychus au fost găsite în asociere cu rămășițele marelui erbivor Tenontosaurus . Acest lucru a fost interpretat ca o dovadă că dromeosauridele au vânat în grupuri precum unele mamifere moderne [39] Această concluzie nu a fost acceptată universal; de fapt, un studiu realizat în 2007 de Roach și Brinkman sugerează că, în loc să fie un vânător social, Deinonychus ar fi fost în schimb un oportunist singuratic care ar fi profitat de prezența altor conspecifici pentru a lansa atacuri dezorganizate asupra prăzilor mari. Diapsidele moderne, inclusiv păsările și crocodilii (cele mai apropiate rude vii de dromeosauridele) demonstrează un comportament de vânătoare puțin cooperant. În schimb, sunt de obicei prădători solitari care sunt ocazional atrași în masă de carcase, unde izbucnesc ocazional conflicte între membrii aceleiași specii. De exemplu, în dragonii Komodo , indivizii mai mari mănâncă mai întâi și îi pot ataca pe cei mai mici încercând să se hrănească în avans. Dacă cel mai mic exemplar cedează în luptă, acesta este canibalizat. Astfel de comportamente sunt în concordanță cu ceea ce a fost descoperit în zonele care demonstrează presupuse dovezi ale comportamentelor de vânătoare de cooperare la dromeosauride. Exemplarele de Deinonychus găsite în aceste zone sunt de fapt toți adolescenții cu membrele lipsă și alte semne de mutilare. [40]
În 2007, oamenii de știință au descris primele urme de dromeosauride găsite în China. Pe lângă confirmarea ipotezei că au mers cu gheara în formă de seceră retrasă de la sol, urmele (lăsate de animale de mărimea lui Achillobator ) arată dovezi ale prezenței a șase indivizi care se deplasează împreună în aceeași direcție de-a lungul unui țărm. Indivizii păstrau o distanță de doar un metru între ei și mergeau într-un ritm destul de lent. Oamenii de știință care au descoperit aceste urme au sugerat că acest lucru dovedește că anumite dromeosauride erau sociale, dar acest lucru nu este suficient pentru a concluziona că au vânat și împreună. [9]
Zburați și alunecați
S-a speculat că cel puțin cinci specii de dromeosauride au reușit să zboare sau cel puțin să alunece. Primul, Rahonavis (o dată clasificat eronat ca o pasăre primitivă)[6] [41] ar fi putut fi capabil de zbor îmbunătățit, după cum indică membrele anterioare echipate cu papile osoase pentru atașarea penelor de zbor.[42] Aripile lui Rahonavis erau mai robuste decât cele ale Archaeopteryx și arată prezența ligamentelor necesare pentru bătutul aripilor. După aceasta, Luis Chiappe a concluzionat că Rahonavis ar putea zbura, dar într-un mod mai incomod decât păsările de astăzi. [43]

O altă specie, Microraptor gui , ar fi putut aluneca folosind atât aripile posterioare, cât și cele din față. Un studiu realizat de Sankar Chatterjee în 2005 a indicat că aripile Microraptorului funcționau ca un biplan și că alunecau în stil „ phugoid ”, aruncându-se de pe un copac și apoi ridicându-se în aer deschizându-și aripile și penele cozii pentru a se agăța de alt copac. . Chatterjee a ajuns la concluzia că Microraptor a arătat și adaptările necesare pentru un zbor îmbunătățit limitat. [13]
Changyuraptor yangi , o rudă apropiată a Microraptor , se crede, de asemenea, că a fost un zburător limitat sau cel puțin o rindelă datorită membrelor sale echilibrate. Cu toate acestea, Changyuraptor era mult mai mare, comparabil cu un curcan obișnuit , făcându-l unul dintre cei mai mari dinozauri zburători non-aviari.
O altă specie, Deinonychus antirrhopus , prezintă o capacitate limitată de a zbura. De fapt, exemplarele tinere au fost prevăzute cu membre superioare mai lungi decât adulții, cu brâuri de umăr mai robuste. Aceste trăsături sunt comparabile cu cele ale altor teropode zburătoare și indică faptul că ar putea zbura când sunt tineri, doar pentru a pierde această abilitate odată ce au crescut. [44]
Simțurile
Studiile comparative asupra inelelor sclerotice ale diferitelor dromeosauride ( Microraptor , Sinornithosaurus și Velociraptor ) și ale păsărilor și reptilelor actuale indică faptul că unele dromeosauride (cum ar fi Microraptor și Velociraptor ) erau prădători nocturni, în timp ce se poate deduce că Sinornithosaurus era catemeral (activ în intervale scurte în timpul zilei). [45] Descoperirea penajului irizat în Microraptor a pus la îndoială ipoteza nocturnă, deoarece nicio pasăre nocturnă din prezent nu prezintă această caracteristică. [46]
Alte studii efectuate pe bulbii olfactivi ai dromeosauridelor au arătat că aceștia aveau o capacitate olfactivă similară cu cea a tiranozauridelor și a păsărilor de astăzi cu un simț al mirosului ridicat, cum ar fi vulturul cu gâtul roșu . Acest lucru arată cât de important a fost acest sens în viața de zi cu zi a dromeosauridelor. [47] [48]
Paleopatologie
În 2001, Bruce Rothschild a studiat fracturile de stres și fracturile de avulsie la scheletele diferitelor teropode. Deoarece fracturile de stres sunt cauzate de traume repetate, se poate deduce că la dromeosauride au fost cauzate de un comportament regulat. Astfel de fracturi au fost de fapt descoperite pe o gheară din față a unui dromeosaurid, probabil suferită în timpul unei lupte cu o pradă. [49]
Clasificare
Relația cu păsările

Dromeosauridele au multe trăsături comune cu păsările primitive. Natura specifică a înrudirii lor cu păsările a fost sursa diverselor studii, diferite ipoteze fiind propuse și respinse de noi descoperiri. Din 2001, Mark Norell și colegii săi au analizat pe larg fosilele diferiților celurozauri , ajungând la concluzia provizorie că dromeosauridele erau dinozaurii non-aviari cei mai înrudiți cu păsările actuale și că troodontidele (de asemenea, cu trăsături similare) față de păsări) erau o linie mai îndepărtată. Au propus chiar că familia Dromaeosauridae era parafileptică în raport cu păsările. [50] În 2002, Hwang și colegii săi au ajuns la concluzia, prin lucrările lui Norell, și noi descoperiri că dromeosauridele și troodontidele erau veri de păsări. [14] Conform paleontologilor, totuși, nu există încă suficiente dovezi pentru a determina dacă dromeosauridele ar putea zbura sau dacă acestea ar fi descendenți din strămoși volatili. [51]
Teorii alternative
Dromeosauridele erau atât de asemănătoare cu păsările, încât unii cercetători au propus clasificarea lor ca păsări adecvate. Fiind înzestrați cu pene, dromeozauridele (împreună cu alți alți celurozauri ) sunt tehnic „păsări” dacă se utilizează definiția tradițională. Alții, precum Lawrence Witmer, au afirmat că clasificarea unui teropod precum Caudipteryx ca pasăre bazată exclusiv pe prezența penelor ar face inutil și lipsit de sens termenul „pasăre”. [52]

S-a propus de mai multe ori că dromeosauridele au fost descendenți de la strămoși volatili. Astfel de ipoteze sunt adesea denumite Birds Came First (BCF), sau „păsările au venit pe primul loc” [53] În lucrările sale, Gregory S. Paul a remarcat numeroase trăsături fiziologice în sprijinul acestei ipoteze, propunând că dromeosauridul ancestral era similar cu Archaeopteryx . În acest caz, dromeozauridele de dimensiuni mari erau comparabile cu struții moderni, care erau și creaturi terestre descendente de la păsări. [23]
În 2002, a fost descoperit Microraptor , cel mai primitiv dromeosaurid găsit la acea vreme. [14] Un an mai târziu, trăsăturile primitive ale Microraptorului , împreună cu aripile și penele sale, au fost folosite ca dovadă că dromeosauridul ancestral era un planer. În acest caz, dromeosauridele mai mari ar pierde treptat această abilitate pe parcursul evoluției lor. [8] Alți cercetători precum Larry Martin susțin că dromeosauridele și alți maniraptorieni (inclusiv păsările) în general nu sunt deloc dinozauri. [54]

În 2005, un specimen de Archaeopteryx a fost descris atât de bine conservat încât s-a putut deduce că era mai aproape de teropodele non-aviare decât se credea anterior. Mai exact, s-a descoperit că Archeopteryxx este prevăzut cu un os palatin primitiv, un deget mare neinversat și un al doilea deget posterior hipertrofiat. Această analiză filogenetică a fost controversată, deoarece a arătat că Confuciusornis era mai mult legat de Microraptor decât de Archaeopteryx , făcând astfel Avialae un taxon parafileptic. S-a propus că paravianul ancestral era capabil să zboare și să alunece și că dromeosauridele și troodontidele erau descendenți care pierduseră aceste abilități. [56] [57] Corfe și Butler au criticat această lucrare pe baza metodologiei. [58]
Aceste ipoteze alternative au fost testate când Turner și colegii săi au descris un nou dromeosaurid în 2007; Mahakala , un gen și mai vechi și mai primitiv al Microraptorului . Mahakala avea membrele superioare prea scurte pentru a-i permite să zboare sau să alunece. Cercetătorii implicați au ajuns la concluzia că norma ancestrală a dromeozauridelor nu includea capacitatea de a zbura și că singurii dinozauri zburători adevărați sunt conținuți în clada Avialae (care include și păsările moderne). [59]
Cu toate acestea, un studiu suplimentar realizat în 2012 a stabilit că cel mai vechi dromeosaurid a fost Xiaotingia , un gen foarte asemănător cu Archaeopteryx , indicând astfel că cel puțin acest membru străvechi al familiei ar putea zbura. [60]
Deinonicosaurie
Din anii 1960 , dromeosauridele au fost adesea clasificate împreună cu troodontidele într-o cladă numită Deinonichosaurie , pe baza prezenței celui de-al doilea deget de la picior posterior hipertrofiat (o trăsătură despre care se știe că este prezentă și la păsările primitive). Numele Deinonychosauria a fost inventat de Ned Colbert și Dale Russell în 1969 și a fost definit ca o cladă (încorporând toate teropodele mai mult legate de dromeosauride decât de păsări) de Jaques Gauthier în 1986. La începutul anilor 2000, consensul dintre paleontologi a fost că dromeosauridele au fost cele mai apropiate rude ale troodontidilor și că împreună cu ei au fost cei mai înrudiți dinozauri cu păsările. [61] În 2012, o analiză filogenetică a 474 de caracteristici prezente în 111 genuri a arătat că Deinonichosauria este un grup monofiletic . [62] Studii mai recente și cu date mai detaliate indică totuși că este probabil ca troodontidele să fie cele mai înrudite cu păsările. [63][64] Întrucât Deinonychosauria a fost inițial definită ca toate animalele mai legate de dromeosauride decât păsările (fără o referință specifică la troodontide), Deinonychosauria ar trebui considerată în mod necesar un sinonim al Dromaeosauridae dacă troodontidele sunt cu adevărat mai mult legate de păsări.[64]
Taxonomie
Familia Dromaeosauridae a fost propusă pentru prima dată de William Diller Matthew și Barnum Brown , care au definit-o ca o subfamilie (Dromaeosaurinae) a familiei Deinodontidae în 1922, conținând doar genul Dromaeosaurus . [65]

Subfamiliile Dromaeosauridae sunt adesea reclasificate pe baza unor noi analize, dar constau de obicei din grupurile enumerate mai jos. Unele tipuri de dromeosauride nu au fost atribuite unei anumite subfamilii, fie pentru că sunt prea slab conservate pentru a fi clasificate cu certitudine, fie adesea pentru că sunt primitive în ceea ce privește subdiviziunile Dromeosauridelor ( Mahakala, de exemplu, este printre cele mai primitive genuri și nu corespund unui subgrup nominalizat până acum). Cea mai primitivă subfamilie este adesea reprezentată de Unenlagiinae. [18] Acest grup enigmatic a fost inclus în familie provizoriu și este probabil ca unii dintre membrii săi să nu fie chiar dromeosaurizi. [67] Genurile de pământ mai mari, cum ar fi Buitreraptor și Unenlagia , prezintă adaptări clare pentru o viață volatilă, deși erau prea mari pentru a lua zborul. Un mic membru al acestui grup, Rahonavis , a fost prevăzut cu aripi bine dezvoltate, cu papile osoase care ancorau penele de zbor, indicând astfel că ar putea zbura. A doua cea mai primitivă cladă este Microraptoria, un grup care include specii mai mici, unele cu adaptări clare la viața arbore. Toate urmele de țesătură ale membrilor acestui grup arată prezența penajului. La fel ca unii unenlagiini, este probabil ca unii microraptorieni să poată zbura. Subfamilia Velociraptorinae a inclus în mod tradițional Velociraptor , Deinonychus și Saurornitholestes și, deși descoperirea lui Tsaagan a susținut această grupare, includerea Saurornitholestes este încă incertă. Subfamilia Dromaeosaurinae este formată din genuri de dimensiuni medii și mari, cu cranii dreptunghiulare (celelalte subfamilii au de obicei boturi mai înguste). [68]




Cea mai mare parte a acestei clasificări se bazează pe Holtz (2011). [68]
- Familia Dromaeosauridae
- Luanchuanraptor
- Nuthetes
- Pamparaptor
- Ornithodesmus
- Variraptor
- Piroraptor
- Shanag
- Zhenyuanlong
- Subfamilia Halszkaraptorinae
- Subfamilia Unenlagiinae
- Subfamilia Microraptorinae
- Nod Eudromaeosauria [10]
- Subfamilia Saurornitholestinae
- Deinonic [70]
- Sottofamiglia Velociraptorinae
- Sottofamiglia Dromaeosaurinae
Storia dei generi

Filogenia
Dromaeosauridae fu prima definito come un clade da Paul Sereno nel 1998 come il gruppo più inclusivo contenente Dromaeosaurus , ma non Troodon , Ornithomimus o Passer . Le varie "sottofamiglie" sono state ridefinite come clade, come tutte le specie più imparentate al genere omonimo che a Dromaeosaurus o gli altri omonimi degli altri sotto-clade (Makovicky per esempio definì Unenlagiinae come tutti i dromaeosauridi più imparentati a Unenlagia che a Velociraptor ). Microraptoria rappresenta l'unico sotto-clade non convertito da una sottofamiglia. [61] [72] L'analisi cladistica di Turner et al. (2012) sostenne la monofilia di Dromaeosauridae. [73]
Questo cladogramma è basato su un'analisi del 2012 da parte di Turner, Makovicky e Norell. [73]
| Dromaeosauridae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Sempre del 2012, questo cladogramma alternativo riflette i risultati degli studi dei paleontologi Phil Senter, James I. Kirkland, Donald D. DeBlieux, Scott Madsen e Natalie Toth. [60]
| Dromaeosauridae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Ricostruzioni

Ricostruzione museale di Deinonychus

Ricostruzione museale di Dromaeosaurus, la posizione prona degli arti anteriori è messa in discussione in alcuni studi

Ricostruzione di Dakotaraptor [74]

Ricostruzione di Velociraptor [75]



Nella cultura di massa
Il dromaeosauride Velociraptor è stato reso famoso per il suo ruolo nel romanzo del 1990 Jurassic Park e dal film omonimo del 1993. Questi "raptor" furono modellati sul dromaeosauride imparentato Deinonychus , che a quel tempo fu rinominato Velociraptor antirrhopus da Gregory Scott Paul . [76] Infatti, i personaggi nel romanzo e nel film scoprono i resti d'un "raptor" in Montana , che è compatibile col raggio geografico di Deinonychus , ma non di Velociraptor , che viveva nell'Asia centrale. [77]
Nel film, l'anatomia e la grandezza dei Velociraptor corrisponde a quella di Deinonychus . [78] [79] Dato che la conferma della presenza di piumaggio in Velociraptor avvenne dopo la distribuzione di Jurassic Park e il suo primo sequel , nel film i "raptor" sono raffigurati con squame simile a quelli dei rettili moderni. In Jurassic Park III (2001), i maschi della specie sono raffigurati con spine calamose sulla testa e la nuca , malgrado il fatto che una tale struttura non sia compatibile con il piumaggio soffice e ben sviluppato degli uccelli moderni. [21] Altre critiche rivolte ai "raptor" dei film includono la posizione prona degli arti anteriori (anatomicamente impossibile nell'animale reale), la loro intelligenza elevata, la loro velocità nella corsa, e il loro comportamento di caccia sociale e cursorio, di cui ci sono poche prove nel Velociraptor propriamente detto. [19]
Note
- ^ Acorn, J., Deep Alberta: Fossil Facts and Dinosaur Digs , University of Alberta Press, 2007, pp. 13 , ISBN 0-88864-481-7 .
- ^ Case, JA, Martin, JE, and Reguero, M. (2007). "A dromaeosaur from the Maastrichtian of James Ross Island and the Late Cretaceous Antarctic dinosaur fauna." Pp. 1–4 in Cooper, A., Raymond, C., and Team, IE (eds.), Antarctica: a Keystone in a Changing World – Online Proceedings for the Tenth International Symposium on Antarctic Earth Sciences , US Geological Survey Open-File Report 2007-1047, SRP 083. US Geological Survey, Washington, DC
- ^ Metcalf, SJ, Vaughan, RF, Benton, MJ, Cole, J., Simms, MJ and Dartnall, DL, A new Bathonian (Middle Jurassic) microvertebrate site, within the Chipping Norton Limestone Formation at Hornsleaslow Quarry, Gloucestershire , in Proceedings of the Geologists' Association , vol. 103, n. 4, 1992, pp. 321-342, DOI : 10.1016/S0016-7878(08)80129-0 .
- ^ a b c JH Ostrom, Osteology of Deinonychus antirrhopus , an unusual theropod from the Lower Cretaceous of Montana ( PDF ), in Peabody Museum of Natural History Bulletin , vol. 30, 1969, pp. 1-165.
- ^ Paul, Gregory S. (2006) "Drawing Dinosaurs"(2006). "Geotimes", January 2006. Copia archiviata , su geotimes.org . URL consultato il 18 dicembre 2007 (archiviato dall' url originale il 2 maggio 2008) .
- ^ a b Peter J. Makovicky, Sebastián Apesteguía e Federico L. Agnolín, The earliest dromaeosaurid theropod from South America , in Nature , vol. 437, n. 7061, 2005, pp. 1007-1011, Bibcode : 2005Natur.437.1007M , DOI : 10.1038/nature03996 , PMID 16222297 .
- ^ a b Norell, MA, and Makovicky, PJ, Dromaeosauridae , in Weishampel, DB, Dodson, P., Osmólska, H. (a cura di), The Dinosauria , 2nd, Berkeley, University of California Press, 2004, pp. 196 –210, ISBN 0-520-24209-2 .
- ^ a b c d e Xu Xing, Four-winged dinosaurs from China , in Nature , vol. 421, n. 6921, 2003, pp. 335-340, Bibcode : 2003Natur.421..335X , DOI : 10.1038/nature01342 , PMID 12540892 .
- ^ a b Rihui Li, Lockley, MG, Makovicky, PJ, Matsukawa, M., Norell, MA, Harris, JD e Liu, M., Behavioral and faunal implications of Early Cretaceous deinonychosaur trackways from China , in Naturwissenschaften , vol. 95, n. 3, 2007, pp. 185-91, Bibcode : 2008NW.....95..185L , DOI : 10.1007/s00114-007-0310-7 , PMID 17952398 . URL consultato il 17 luglio 2015 (archiviato dall' url originale il 9 febbraio 2020) .
- ^ a b NR Longrich e PJ Currie,A microraptorine (Dinosauria–Dromaeosauridae) from the Late Cretaceous of North America , in PNAS , vol. 106, n. 13, 2009, pp. 5002-7, Bibcode : 2009PNAS..106.5002L , DOI : 10.1073/pnas.0811664106 , PMC 2664043 , PMID 19289829 .
- ^ Csiki Z., Vremir, M., Brusatte, SL e Norell, MA,An aberrant island-dwelling theropod dinosaur from the Late Cretaceous of Romania , in Proceedings of the National Academy of Sciences of the United States of America , vol. 107, n. 35, 2010, pp. 15357-61, Bibcode : 2010PNAS..10715357C , DOI : 10.1073/pnas.1006970107 , PMC 2932599 , PMID 20805514 . Supporting Information
- ^ a b Mark A. Norell e Makovicky, Peter J., Important features of the dromaeosaurid skeleton II: information from newly collected specimens of Velociraptor mongoliensis , in American Museum Novitates , vol. 3282, 1999, pp. 1-45.
- ^ a b S. Chatterjee e RJ Templin, Biplane wing planform and flight performance of the feathered dinosaur Microraptor gui ( PDF ), in Proceedings of the National Academy of Sciences , vol. 104, n. 5, 2007, pp. 1576-1580, Bibcode : 2007PNAS..104.1576C , DOI : 10.1073/pnas.0609975104 , PMC 1780066 , PMID 17242354 .
- ^ a b c SH Hwang, MA Norell, Q. Ji e K. Gao, New Specimens of Microraptor zhaoianus (Theropoda: Dromaeosauridae) from Northeastern China , in American Museum Novitates , n. 3381, 2002, pp. 44pp.
- ^ A. Perle, MA Norell e J. Clark, A new maniraptoran theropod – Achillobator giganticus (Dromaeosauridae) – from the Upper Cretaceous of Burkhant, Mongolia , in Contributions of the Mongolian-American Paleontological Project , vol. 101, 1999, pp. 1-105.
- ^ Britt, Chure, Stadtman, Madsen, Scheetz e Burge, New osteological data and the affinities of Utahraptor from the Cedar Mountain Fm. (Early Cretaceous) of Utah , in Journal of Vertebrate Paleontology , vol. 21, n. 3, 2001, p. 36A, DOI : 10.1080/02724634.2001.10010852 .
- ^ Naish, D. Hutt, and Martill, DM (2001). "Saurischian dinosaurs: theropods." in Martill, DM and Naish, D. (eds). Dinosaurs of the Isle of Wight . The Palaeontological Association, Field Guides to Fossils. 10, 242–309.
- ^ a b AS Turner, Hwang, SH e Norell, MA, A small derived theropod from Öösh, Early Cretaceous, Baykhangor Mongolia ( PDF ), in American Museum Novitates , vol. 3557, 2007, pp. 1-27, DOI : 10.1206/0003-0082(2007)3557[1:ASDTFS]2.0.CO;2 . URL consultato il 29 marzo 2007 .
- ^ a b Andrea Cau, Theropoda volume II: Deinonychosauria , Amazon Media EU S.à rl, 2012.
- ^ a b Xu, X., Wang, X.-L., and Wu, X.-C., A dromaeosaurid dinosaur with a filamentous integument from the Yixian Formation of China , in Nature , vol. 401, n. 6750, 1999, pp. 262-266, Bibcode : 1999Natur.401..262X , DOI : 10.1038/45769 .
- ^ a b c d AH Turner, Makovicky, PJ e Norell, MA, Feather quill knobs in the dinosaur Velociraptor ( PDF ), in Science , vol. 317, n. 5845, 2007, p. 1721, Bibcode : 2007Sci...317.1721T , DOI : 10.1126/science.1145076 , PMID 17885130 .
- ^ a b Robert A. DePalma, David A. Burnham, Larry D. Martin, Peter L. Larson and Robert T. Bakker (2015). "The first giant raptor (Theropoda: Dromaeosauridae) from the Hell Creek Formation" . Paleontological Contributions 14: 1–16.
- ^ a b Paul, Gregory S. (2002). Dinosaurs of the Air: The Evolution and Loss of Flight in Dinosaurs and Birds. Baltimore: Johns Hopkins University Press. 472 pp.
- ^ R. Prum e AH Brush, The evolutionary origin and diversification of feathers , in The Quarterly Review of Biology , vol. 77, n. 3, 2002, pp. 261-295, DOI : 10.1086/341993 , PMID 12365352 .
- ^ Adams, Dawn (1987) "The bigger they are, the harder they fall: Implications of ischial curvature in ceratopsian dinosaurs" pg 1–6 in Currie, Philip J. and Koster, E. (eds) Fourth symposium on mesozoic terrestrial ecosystems. Tyrrell Museum, Drumheller, Canada
- ^ Manning, PL, Payne, D., Pennicott, J., Barrett, PM, and Ennos, RA, Dinosaur killer claws or climbing crampons? , in Biology Letters , vol. 2, n. 1, 2005, pp. 110-112, DOI : 10.1098/rsbl.2005.0395 , PMC 1617199 , PMID 17148340 (archiviato dall' url originale il 23 gennaio 2012) .
- ^ a b Denver W. Fowler, The Predatory Ecology of Deinonychus and the Origin of Flapping in Birds , 14 dicembre 2011, DOI : 10.1371/journal.pone.0028964 .
- ^ a b Davies, SJJF (2002) "Ratites and Tinamous" Oxford University Press. New York, USA
- ^ Gilliard, ET (1958) "Living birds of the world" Doubleday, garden City, NY. USA. 400p.
- ^ Christopher P. Kofron,Attacks to humans and domestic animals by the southern cassowary (Casuarius casuarius johnsonii) in Queensland, Australia , in Journal of Zoology , vol. 249, n. 4, 1999, pp. 375-381, DOI : 10.1111/j.1469-7998.1999.tb01206.x .
- ^ Christopher P. Kofron, Case histories of attacks by the southern cassowary in Queensland , in Memoirs of the Queensland Museum , vol. 49, n. 1, 2003, pp. 335-338.
- ^ Kent H. Redford e Gustav Peters, Notes on the biology and song of the red-legged seriema (cariama cristata) , in Journal of Field Ornithology , vol. 57, n. 4, 1986, pp. 261-269, JSTOR 4513154 .
- ^ a b PL Manning, L. Margetts, MR Johnson, PJ Withers, WI Sellers, PL Falkingham, PM Mummery, PM Barrett e DR Raymont, Biomechanics of dromaeosaurid dinosaur claws: Application of X-ray microtomography, nanoindentation, and finite element analysis , in The Anatomical Record: Advances in Integrative Anatomy and Evolutionary Biology , vol. 292, n. 9, 2009, pp. 1397-1405, DOI : 10.1002/ar.20986 .
- ^ Velociraptor's 'killing' claws were for climbing , in New Scientist , n. 2725, settembre 2009. URL consultato il 15 settembre 2009 .
- ^ Senter, P., Pedal function in deinonychosaurs (Dinosauria: Theropoda): a comparative study , in Bulletin of the Gunma Museum of Natural History , vol. 13, 2009, pp. 1-14.
- ^ Therrien, F., Henderson, DM and Huff, CB (2005). "Bite me: biomechanical models of theropod mandibles and implications for feeding behavior." pp. 179–237 in Carpenter, K. (ed.), The Carnivorous Dinosaurs . Indianapolis: Indiana University Press.
- ^ DOI : 10.1098/rspb.2010.0794
- ^ A. Mudroch, U. Richter, U. Joger, R. Kosma, O. Idé e A. Maga, Didactyl tracks of paravian theropods (Maniraptora) from the ?Middle Jurassic of Africa , in PLoS ONE , vol. 6, n. 2, 2011, p. e1462, DOI : 10.1371/journal.pone.0014642 . Figure 4.
- ^ WD Maxwell e Ostrom, JH, Taphonomy and paleobiological implications of Tenontosaurus - Deinonychus associations , in Journal of Vertebrate Paleontology , vol. 15, n. 4, 1995, pp. 707-712, DOI : 10.1080/02724634.1995.10011256 .
- ^ BT Roach e DL Brinkman, [103:AROCPH 2.0.CO;2 A reevaluation of cooperative pack hunting and gregariousness in Deinonychus antirrhopus and other nonavian theropod dinosaurs ], in Bulletin of the Peabody Museum of Natural History , vol. 48, n. 1, 2007, pp. 103-138, DOI : 10.3374/0079-032X(2007)48[103:AROCPH]2.0.CO;2 .
- ^ MA Norell, JM Clark, AH Turner, PJ Makovicky, R. Barsbold e T. Rowe, A new dromaeosaurid theropod from Ukhaa Tolgod (Omnogov, Mongolia) , in American Museum Novitates , vol. 3545, 2006, pp. 1 –51, DOI : 10.1206/0003-0082(2006)3545[1:ANDTFU]2.0.CO;2 .
- ^ Catherine A. Forster, Sampson, Scott D., Chiappe, Luis M. e Krause, David W., The Theropod Ancestry of Birds: New Evidence from the Late Cretaceous of Madagascar , in Science , vol. 279, n. 5358, 1998a, pp. 1915-1919, Bibcode : 1998Sci...279.1915F , DOI : 10.1126/science.279.5358.1915 , PMID 9506938 . . (HTML abstract).
- ^ LM Chiappe, Glorified Dinosaurs: The Origin and Early Evolution of Birds , Sydney, UNSW Press, ISBN 0-471-24723-5 .
- ^ William L. Parsons e Parsons, Kristen M., Morphological Variations within the Ontogeny of Deinonychus antirrhopus (Theropoda, Dromaeosauridae) , in PLoS ONE , vol. 10, n. 4, 2015, DOI : 10.1371/journal.pone.0121476 , e0121476.
- ^ Schmitz, L.; Motani, R. e Motani, Nocturnality in Dinosaurs Inferred from Scleral Ring and Orbit Morphology , in Science , vol. 332, n. 6030, 2011, pp. 705-8, Bibcode : 2011Sci...332..705S , DOI : 10.1126/science.1200043 , PMID 21493820 .
- ^ Quanguo Li, K.-Q. Gao, Q. Meng, JA Clarke, MD Shawkey, L. d'Alba, R. Pei, M. Ellison, MA Norell e J. Vinther, Reconstruction of Microraptor and the Evolution of Iridescent Plumage , in Science , vol. 335, n. 6073, 9 marzo 2012, pp. 1215-1219, Bibcode : 2012Sci...335.1215L , DOI : 10.1126/science.1213780 , PMID 22403389 .
- ^ Zelenitsky, DK; Therrien, F.; Kobayashi, Y.,Olfactory acuity in theropods: palaeobiological and evolutionary implications , in Proceedings of the Royal Society B , vol. 276, n. 1657, 2009, pp. 667-673, DOI : 10.1098/rspb.2008.1075 , PMC 2660930 , PMID 18957367 .
- ^ Zelenitsky, DK; Therrien, F.; Ridgely, RC; McGee, AR; Witmer, LM, Evolution of olfaction in non-avian theropod dinosaurs and birds , in Proceedings of the Royal Society B , in press, n. 1725, 2011, pp. 3625-3634, DOI : 10.1098/rspb.2011.0238 .
- ^ Rothschild, B., Tanke, DH, and Ford, TL, 2001, Theropod stress fractures and tendon avulsions as a clue to activity: In: Mesozoic Vertebrate Life, edited by Tanke, DH, and Carpenter, K., Indiana University Press, p. 331-336.
- ^ Norell, M. Clark, JM, Makovicky, PJ (2001). " Phylogenetic relationships among coelurosaurian theropods. " New Perspectives on the Origin and Evolution of Birds: Proceedings of the International Symposium in Honor of John H. Ostrom , Yale Peabody Museum: 49–67
- ^ Kevin Padian e Kenneth P. Dial, Origin of flight: Could 'four-winged' dinosaurs fly? , in Nature , vol. 438, n. 7066, 2005, pp. E3; discussion E3–4, Bibcode : 2005Natur.438E...3P , DOI : 10.1038/nature04354 , PMID 16292258 .
- ^ Witmer, LM (2005) "The Debate on Avian Ancestry; Phylogeny, Function and Fossils.” In "Mesozoic Birds: Above the Heads of Dinosaurs”, pp. 3–30. ISBN 0-520-20094-2
- ^ Olshevsky, George. (1994). "The birds first? A theory to fit the facts — evolution of reptiles into birds". Omni , June 1994. Volume 16 No. 9
- ^ LD Martin, A basal archosaurian origin for birds ( PDF ), in Acta Geologica Sinica , vol. 50, n. 6, 2004, pp. 978-990.
- ^ National Geographic News- Earliest Bird Had Feet Like Dinosaur, Fossil Shows – Nicholas Bakalar, December 1, 2005, Page 2. Retrieved 2006-10-18.
- ^ G Mayr, A Well-Preserved Archaeopteryx Specimen with Theropod Features , in Science , vol. 310, n. 5753, 2005, pp. 1483-1486, Bibcode : 2005Sci...310.1483M , DOI : 10.1126/science.1120331 , PMID 16322455 .
- ^ G Mayr e Peters, SD, Response to comment on 'A Well-Preserved Archaeopteryx Specimen with Theropod Features' , in Science , vol. 313, n. 5791, 2006, pp. 1238c, Bibcode : 2006Sci...313.1238M , DOI : 10.1126/science.1130964 .
- ^ IJ Corfe e Butler, RJ, Comment on 'A Well-Preserved Archaeopteryx Specimen with Theropod Features' , in Science , vol. 313, n. 5791, 2006, pp. 1238b, Bibcode : 2006Sci...313.1238C , DOI : 10.1126/science.1130800 , PMID 16946054 .
- ^ Alan H. Turner, Diego Pol, Julia A. Clarke, Gregory M. Erickson e Mark Norell, A basal dromaeosaurid and size evolution preceding avian flight , in Science , vol. 317, n. 5843, 2007, pp. 1378-1381, Bibcode : 2007Sci...317.1378T , DOI : 10.1126/science.1144066 , PMID 17823350 .
- ^ a b c P. Senter, JI Kirkland, DD DeBlieux, S. Madsen e N. Toth, New Dromaeosaurids (Dinosauria: Theropoda) from the Lower Cretaceous of Utah, and the Evolution of the Dromaeosaurid Tail , in PLoS ONE , vol. 7, n. 5, 2012, p. e36790, DOI : 10.1371/journal.pone.0036790 , PMC 3352940 , PMID 22615813 .
- ^ a b Phil Senter, R. Barsbold, Brooks B. Britt e David B. Burnham, Systematics and evolution of Dromaeosauridae (Dinosauria, Theropoda) , in Bulletin of the Gunma Museum of Natural History , vol. 8, 2004, pp. 1-20.
- ^ Turner, AH, Makovicky, PJ, and Norell, MA 2012. A review of dromaeosaurid systematics and paravian phylogeny. Bulletin of the American Museum of Natural History 371: 1–206.
- ^ DOI : 10.1038/nature12168
- ^ a b Mortimer, M. (2012) The Theropod Database: Phylogeny of taxa Archiviato il 16 maggio 2013 in Internet Archive .. Retrieved 2013-AUG-15.
- ^ WD Matthew e B. Brown, The family Deinodontidae, with notice of a new genus from the Cretaceous of Alberta , in Bulletin of the American Museum of Natural History , vol. 46, 1922, pp. 367-385.
- ^ Michael Pittman, Rui Pei, Xing Xu The first dromaeosaurid (Dinosauria:Theropoda) from the Lower Cretaceous Bayan Gobi Formation of Nei Mongol, China PeerJ PrePrints 3:e1654
- ^ Agnolin, FL; Novas, FE, Unenlagiid theropods: are they members of the Dromaeosauridae (Theropoda, Maniraptora)? ( PDF ), in Anais da Academia Brasileira de Ciências , vol. 83, 2011, pp. 117-162, DOI :10.1590/S0001-37652011000100008 . URL consultato il 23 aprile 2011 .
- ^ a b Holtz, Thomas R. Jr. (2011) Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages, Winter 2011 Appendix.
- ^ SJ Czerkas, Feathered Dinosaurs and the Origin of Flight , in The Dinosaur Museum Journal , vol. 1, Blanding, 2002.
- ^ a b Longrich, NR & Currie, PJ,A microraptorine (Dinosauria-Dromaeosauridae) from the Late Cretaceous of North America , in Proceedings of the National Academy of Sciences , vol. 106, n. 13, 2009, pp. 5002-5007, DOI : 10.1073/pnas.0811664106 , PMC 2664043 , PMID 19289829 .
- ^ Sweetman, SC, The first record of velociraptorine dinosaurs (Saurischia, Theropoda) from the Wealden (Early Cretaceous, Barremian) of southern England , in Cretaceous Research , vol. 25, n. 3, 2004, pp. 353-364, DOI : 10.1016/j.cretres.2004.01.004 .
- ^ Sereno, PC 2005. Stem Archosauria—TaxonSearch Archiviato il 15 gennaio 2009 in Internet Archive . [version 1.0, November 7, 2005
- ^ a b DOI : 10.1206/748.1
- ^ Robert A. DePalma, David A. Burnham, Larry D. Martin, Peter L. Larson, Robert T. Bakker, The first giant raptor (Theropoda: Dromaeosauridae) from the Hell Creek Formation , Paleontological Contributions, n. 14, The University of Kansas, Paleontological Institute, 30 October , 2015 Lawrence, Kansas, USA ISSN 1946-0279
- ^ Senter P, Robins JH (2015) Resting Orientations of Dinosaur Scapulae and Forelimbs: A Numerical Analysis, with Implications for Reconstructions and Museum Mounts . PLoS ONE 10(12): e0144036. doi:10.1371/journal.pone.0144036
- ^ Gregory Scott Paul (1988) Predatory Dinosaurs of the World , Simon and Schuster, pp. 362-363
- ^ Michael Crichton , Jurassic Park , New York, Alfred A. Knopf, 1990, pp. 117 , ISBN 0-394-58816-9 .
- ^ Jody Duncan, The Winston Effect , London, Titan Books, 2006, p. 175, ISBN 1-84576-365-3 .
- ^ Robert T. Bakker , Raptor Red , New York, Bantam Books, 1995, p.4 , ISBN 0-553-57561-9 .
Bibliografia
- Czerkas, SA, Zhang, D., Li, J., and Li, Y., Flying Dromaeosaurs , in Czerkas, SJ (a cura di), Feathered Dinosaurs and the Origin of Flight: The Dinosaur Museum Journal 1 , Blanding, The Dinosaur Museum, 2002, pp. 16-26.
- Colbert, E. and DA Russell, The small Cretaceous dinosaur Dromaeosaurus , in American Museum Novitates , vol. 2380, 1969, pp. 1-49.
- Hopson, JA, Relative brain size in dinosaurs: implications for dinosaurian endothermy , in American Association for the Advancement of Science Symposium , vol. 28, 1980, pp. 287-310.
- Stefano Broccoli, Marco Castiello, Marco Lampugnani, In dino veritas , 2011.
- Qiang Ji, Mark A. Norell, Ke-Qin Gao, Shu-An Ji and Dong Ren, Distribution of integumentary structures in a feathered dinosaur , in Nature , vol. 410, 26 April 2001, pp. 1084-1088, DOI : 10.1038/35074079 .
- Kirkland, JI, R. Gaston, et al., A large dromaeosaur (Theropoda) from the Lower Cretaceous of Eastern Utah , in Hunteria , vol. 2, 1993, pp. 1-16.
- Makovicky, Apesteguía and Agnolín, The earliest dromaeosaurid theropod from South America , in Nature , vol. 437, 2005, pp. 1007-1011.
- Metcalf SJ, Vaughan RF, Benton MJ, Cole J, Simms MJ & Dartnall DL, A new Bathonian (Middle Jurassic) microvertebrate site, within the Chipping Norton Limestone Formation at Hornsleaslow Quarry, Gloucestershire , in Proceedings of the Geologists' Association , vol. 103, 1992, pp. 321-342.
- Norell, Mark A. and PJ Makovicky, Dromaeosauridae , in DB Weishampel, P. Dodson and H. Osmolska (a cura di), The Dinosauria , Berkeley, University of California Press, 2004, pp. 196-209.
- JH Ostrom , Osteology of Deinonychus antirrhopus, an unusual theropod from the Lower Cretaceous of Montana , in Peabody Museum of Natural History Bulletin , vol. 30, 1969, pp. 1-165.
- Paul, GS,Dinosaurs of the Air: The Evolution and Loss of Flight in Dinosaurs and Birds , Baltimore, Johns Hopkins University Press, 2002.
- Perle, A., M. Norell and J. Clark, A new maniraptoran theropod Achillobator giganticus (Dromaeosauridae) from the Upper Cretaceous of Burkhant, Mongolia , in Contributions of the Department of Geology, National University of Mongolia, Ulaanbaatar , vol. 101, 1999, pp. 1-105.
- Prum, R. & Brush AH, The evolutionary origin and diversification of feathers , in The Quarterly Review of Biology , vol. 77, 2002, pp. 261-295.
- Senter, P., R. Barsold, BB Britt and DA Burnham, Systematics and evolution of Dromaeosauridae (Dinosauria, Theropoda) , in Bulletin of the Gunma Museum of Natural History , vol. 8, 2004, pp. 1-20.
- Sereno, PC, The logical basis of phylogenetic taxonomy , in Systematic Biology , vol. 51, 2005, pp. 1-25.
- Xing Xu, Xiao-Lin Wang and Xiao-Chun Wu, A dromaeosaurid dinosaur with a filamentous integument from the Yixian Formation of China , in Nature , vol. 401, 16 settembre 1999, pp. 262-266, DOI : 10.1038/45769 .
- Xing Xu, Zhonghe Zhou, Xiaolin Wang, Xuewen Kuang, Fucheng Zhang and Xiangke Du, Four winged dinosaurs from China , in Nature , vol. 421, 23 gennaio 2003, pp. 335-340, DOI : 10.1038/nature01342 .
Altri progetti
-
 Wikimedia Commons contiene immagini o altri file su Dromaeosauridae
Wikimedia Commons contiene immagini o altri file su Dromaeosauridae -
 Wikispecies contiene informazioni su Dromaeosauridae
Wikispecies contiene informazioni su Dromaeosauridae
Collegamenti esterni
- The Dromaeosauridae: The Raptors! , from the University of California Berkeley Museum of Paleontology.