Deinonychus antirrhopus
| Deinonic | |
|---|---|
 | |
| Gama geologică | |
| Starea de conservare | |
Fosil | |
| Clasificare științifică | |
| Domeniu | Eukaryota |
| Regatul | Animalia |
| Phylum | Chordata |
| Superordine | Dinozauria |
| Ordin | Saurischia |
| Subordine | Theropoda |
| Familie | † Dromaeosauridae |
| Subfamilie | † Dromaeosaurinae |
| Tip | † Deinonychus Ostrom , 1969 |
| Nomenclatura binominala | |
| † Deinonychus antirrhopus Ostrom, 1969 | |
| Sinonime | |
Velociraptor antirrhopus Paul , 1988 | |
Deinonic (Deinonychus Ostrom , 1969 ) este un gen de dromaeosaurid coelurosaurid dinozaur care include doar un singur gasit specie, Deinonychus antirrhopus. Această specie, care ar putea ajunge la 3,4 metri în lungime, a trăit în Cretacicul inferior în urmă cu aproximativ 115-108 milioane de ani (de la mijloc Apțianul la inferior Albian ). Rămășițele sale au fost găsite în statele americane Montana , Wyoming , și Oklahoma , în stîncile formarea Clovery și formarea Antlers. Cu toate acestea, dintii care poate să aparțină deinonic au fost găsite în continuare la est , în diferite zone ale Maryland .
Studiile efectuate pe deinonic de paleontologul John Ostrom la sfârșitul anilor 1960 a revoluționat modul în care oamenii de știință perceput dinozauri, creându -se astfel așa-numita „Renașterea dinozaur“ și o dezbatere asupra faptului dacă acestea au fost endotermic (cu sânge cald). Anterior, dinozaurii erau în mod popular descriși ca reptile mari stângace. Ostrom remarcat faptul că, contrar acestei imagini, cu deinon are un corp mic, o postură orizontală, o paleo- pasăre cum ar fi coloanei vertebrale, și gheare curbate pe picioare, ceea ce presupune că animalul a fost un prădător agil si activ.. [1]
Numele său, care înseamnă „gheară teribilă”, se referă la gheara uriașă curbată de pe degetul secund al piciorului din spate. Deoarece ghearele moderne archosaurs (păsări și crocodili) sunt alungite cu un capac de cheratină, Ostrom a estimat că, în viață, Gheara deinonic ar putea ajunge la 120 mm în lungime. [1] Specia nume înseamnă antirrhopus „împotriva echilibrului“, referindu -se la ipoteza lui Ostrom privind funcția cozii. Ca și alte dromaeosaurids, coada vertebre posedat o serie de tendoane osificate și alungite apofize . Inițial, acest lucru a condus la ideea că coada era un echilibru rigid, dar o rudă a Velociraptor mongoliensis ( IGM 100/986) are o coadă curbată lateral în formă de S. Acest lucru indică faptul că, în starea vie, coada ar putea curbați flexibil către laturi. [2]
În ambele formațiunile Cloverly și coarne de cerb, dinții deinon au fost găsite în apropierea sau asociate cu resturi fosile ale ornithopozi Tenontosaurus , indicând astfel că deinon l vânat sau hrănit pe carcasele sale.
Jurassic Park Velociraptorii și ei de film omonim au fost modelate după Deinonychus. [3] [4]
Descriere
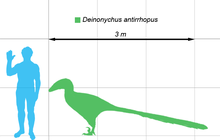
Pe baza unor exemplare mature, [5] se estimează că Deinonychus ar putea ajunge la 3,4 metri lungime, cu un craniu lung de 410 mm, o înălțime în spate de 0,87 metri și o greutate de 73 de kilograme (dar unii experți propun o greutate de 100 de kilograme). [6] [7] Craniul a fost furnizat de mandibulele puternice cu aproximativ șaptezeci dinți ascuțite. Reconstituiri craniu timpuriu (bazate pe resturi incomplete) asemănat că a Allosaurus , dar rămâne mai complete a descoperit mai târziu a arătat că palatului era mai spațioasă decât se credea anterior, indicând faptul că botul era mai îngust, în timp ce pomeții protruded sensibil, indicând o vedere stereoscopică.[8] Atât craniul și maxilarul inferior au deschideri care au redus greutatea craniului. În Deinonychus, fereastra antorbital (deschiderea dintre ochi și nări) a fost deosebit de mare.[8] Similar cu Dromaeosaurus , craniul lui Deinonychus era mai robust decât cea a Velociraptor , și poseda un os nazal superior. [9]

Deinonychus poseda picioare din fata mari echipate cu trei degete dotate cu gheare. Primul deget a fost cel mai scurt, iar al doilea a fost cel mai lung. Fiecare picior din spate era echipat cu patru degete, dintre care primul era foarte mic și redus la un pinten, în timp ce al doilea deget era foarte dezvoltat și susținea o gheară curbată în formă de seceră, care ar fi putut fi folosită în timpul prădării. Această gheară a fost ținută constant ridicată de la sol, pentru a preveni deteriorarea acesteia în timpul alergării. Celelalte două degete susțineau greutatea animalului. [10]
Deși nu a fost găsit nici un impresii ale pielii de Deinonychus, există rămășițe fosile ale unor dromaeosaurids, din care face parte deinonic, cu urme ale prezenței pene. [11] Genul Microraptor este mai veche și mai primitiv filogenetic decât deinonic, și face parte din aceeași familie. [12] Multe fosile Microraptor arată prezența moderne de pasăre penaj pe brațe, picioare și coadă. [11] Velociraptor este un gen mai puțin străvechi decât deinonic, dar este mai mult legat de acesta decât Microraptor, și cel puțin un specimen arată prezența papilelor osoase pe ulna compatibil cu atașarea obligeana a remiges . [13]
Clasificare

Deinonychus antirrhopus reprezintă una dintre cele mai studiate dromaeosaurid specii, [14] și este o rudă apropiată a Velociraptor , găsit mai recente cretacice târzii sedimente din Asia Centrală. [15] [16] Împreună, acestea formează Velociraptorinae clade . Subfamilia Numele Velociraptorinae a fost folosit pentru prima dată de Rinchen Barsbold în 1983 [17] să conțină numai Velociraptor. Mai târziu, Phil Currie a inclus cele mai multe cunoscute dromaeosaurids în ea. [18] Două Cretacicul superior genuri, Tsaagan Mongoliei [15] și Saurornitholestes din America de Nord [6] ar putea fi legate, dar aceasta din urmă prezintă dificultatea de a fi clasificate. [15] Velociraptor și aliații săi se crede că au gheare cele mai utilizate în vânătoare , mai degrabă decât pe dinți, spre deosebire de dromaeosaurids , cum ar fi Dromaeosaurus , care au cranii mai puternice. [19] Împreună cu troodontids , dromaeosaurids formeaza Deinonychosauria clade, care reprezintă o soră taxon de Aves . Filogenetic , Deinonychosauria reprezintă grupul de dinozauri non-aviare cel mai strâns legate de păsări. [20]
Următoarea cladogramă se bazează pe analiza de Robert DePalma, David Burnham, Larry Martin, Peter Larson si Robert Bakker (2015), folosind date actualizate de la Grupul de lucru theropod. [21]
| Dromaeosauridae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Descoperire
Rămășițele fosilizate ale Deinonychus au fost recuperate de la formarea Cloverly de Montana și Wyoming , [1] și Oklahoma Formațiunea de coarne de cerb, [22] în America de Nord . De Cloverly Datele de constituire din Late Apțianul la Upper Albian a cretacicului de Jos , în urmă cu aproximativ 115-108 de milioane de ani. [23] [24] Se poate întâmpla ca dinții descoperite în formarea Arundel din Maryland pot fi atribuite genului. [25]
Primele Rămășițele au fost descoperite în 1931 în sudul Montana lângă Billings . Șeful echipei de paleontologie, Barnum Brown, s-a concentrat pe găsirea rămășițelor ornitopodului Tenontosaurus , dar raportul său scris către Muzeul American de Istorie Naturală a inclus o încuviințare către un mic dinozaur carnivor de lângă Tenontosaurus , ca fiind „încorporat în matrice și dificil. de pregătit ". [26] El a numit agilis Daptosaurus specimen, și care este destinat să publice o descriere și de a restabili scheletul său, specimen AMNH 3015, dar nu a putut să termine treaba. [27] Brown , mai târziu , a adus înapoi de la formarea Cloverly scheletul unui theropod mai mici cu dinți aparent supradimensionate pe care a numit informal „Megadontosaurus“. John Ostrom, revizuiască toate aceste decenii mai târziu, a dat seama că dinții au venit de la un specimen Deinonychus, în timp ce restul scheletului a venit de la un animal complet diferit. Mai târziu , acest model a fost descris și numit microvenator . [27]

Un pic peste treizeci de ani mai târziu, în august 1964 paleontologul John Ostrom a condus o expediție de la Universitatea Yale Muzeul Peabody, care a descoperit ramasite suplimentare la Bridger . Expedițiile din următorii doi ani au găsit peste 1.000 de oase, inclusiv cel puțin trei persoane. Așa cum a fost aproape imposibil de a clarifica asocierea dintre oasele și de a determina numărul de persoane, Deinonychus holotype YPM 5205 a fost limitată la piciorul stâng intact și piciorul drept parțial care au fost determinate să fie de la același individ. [10] Specimenele rămase au fost catalogate în cincizeci de intrări separate la Peabody Muzeul Yale, deși ele pot să provină din cât mai puține trei indivizi. [10]
Studii mai târziu de către Ostrom și Grant E. Meyer a arătat că Brown „Daptosaurus“ și Deinonychus făceau parte din același gen. Ostrom a publicat concluziile sale în februarie 1969, care denumesc rămâne Deinonychus antirrhopus. Numele specific, antirrhopus, deriva din ἀντίρροπος grecesc (contra echilibru), cu referire la funcția probabilă a cozii rigide. [28] Cinci luni mai târziu, Ostrom a publicat o analiză detaliată monografie asupra animalului. [10]
În ciuda descoperirii unei nenumărate de oase în 1969, multe dintre ele s-au pierdut sau s-au dovedit dificil de interpretat. Constatarile continute orice materiale postcranici, incomplete vertebre, și nimic femure , sacru , forcule (altfel cunoscut sub numele de os de dorințe), și sternul . Reconstrucția scheletice a Deinonychus de Ostrom a inclus un apartament, trapezoidal pubis foarte diferită de cea a celorlalte theropods, dar care a arătat aceeași lungime ca ischiomului. [10]
Alte descoperiri

În 1974, Ostrom a publicat o altă monografie pe umărul Deinonychus', în care a dat seama că «pubis» a primei sale de reconstrucție a fost de fapt un os coracoid. [29] În același an, un alt specimen Deinonychus (MCZ 4371) a fost descoperit în Montana , de Steven Orzack în timpul unei Universitatea Harvard expediție. Specimenul a inclus diverse elemente noi, inclusiv femure, pubis, sacrum, un iliac mai bine conservate, și unele piese ale picioarelor și metatarsiene , . Ostrom și-a modificat reconstrucția, încorporând un pubis mai lung, despre care Ostrom a remarcat că era la fel de retrovertit ca și cel al păsărilor. [30]
În 2000, au fost examinate oase izolate și fragmente osoase depozitate în Muzeul American. Examinările au dezvăluit o caracteristică trecută cu vederea; lung, oase subțiri, mai întâi identificate ca tendoanele coada osificate, sa dovedit a fi de fapt gastralia . Chiar mai remarcabil a fost descoperirea de fragmente de ou în vecinătatea specimenul originale Deinonychus. [31]
Un raport ulterior a identificat fragmente de ou ca și cele ale Deinonychus, reprezentând astfel ouăle dromaeosaurid mai întâi identificate pozitiv. [26] De asemenea, sa observat că fragmentele au fost în contact cu gastralia, sugerând astfel că Deinonychus incubat ouăle și a fost endotermic ca păsările actuale. [32] Un alt studiu a arătat că individul era 13-14 ani la momentul morții, și că a finalizat ciclul său de creștere. [33]
Implicații
Descrierea Ostrom a Deinonychus în 1969 a fost laudata ca fiind cea mai importanta descoperire in dinozaur paleontologie al XX-lea. [34] Descoperirea acestui pradator activ și agil a schimbat percepția științifice și populare despre imaginea dinozauri, și a declanșat o dezbatere asupra faptului dacă dinozaurii erau cu sânge cald. Această dezvoltare a fost numită așa-numita „Renașterea dinozaurilor”. Câțiva ani mai târziu, Ostrom a observat asemănările dintre membrele anterioare Deinonychus' și cele ale păsărilor, o observație care l -au dus să reînvie ipoteza că păsările trag din dinozauri. [35] Patruzeci de ani mai târziu, această idee este aproape universal acceptată.
Din cauza sale foarte păsări cum ar fi anatomie, și înrudirea sale aproape de alte dromaeosaurids, paleontologii speculează că Deinonychus a fost acoperit de pene. [36] [37] [38] există dovezi de netăgăduit de pene moderne pentru diferite Dromaeosaurids legate de Deinonychus, inclusiv Velociraptor și Microraptor , cu toate că o astfel de dovadă nu a fost încă descoperit pentru Deinonychus. [11] [39] Atunci când studiază discipline , cum ar fi abilitățile motorii, paleontologii cum ar fi Phil Senter ține cont de prezența probabilă a pene . [40]
Paleobiologie
Comportamente de vânătoare
Dintii Deinonychus au fost descoperite în număr mare de fosile ale ornithopozi Tenontosaurus în formarea Cloverly. Două cariere au fost descoperite, care conțin fosile aproape complet Deinonychus aproape de fosile Tenontosaurus. Primul, cariera Yale din Montana , contine mai multi dinti si fosile de patru pentru adulți și unul tineri Deinonychus. Proximitatea acestor specimene la Tenontosaurus sugerează posibilitatea ca Deinonychus au fost devorarea animalul, și poate fi chiar ucis. Ostrom și Maxwell au speculat că această constatare a indicat că Deinonychus trăit și vânate în pachete. [42] A doua carieră provine din formațiunea Coarne din Oklahoma . Site - ul conține șase schelete parțiale ale Tenontosaurus, inclusiv un număr mare de dinți Deinonychus. Amprentele dinților Deinonychus "au fost descoperite pe humerusului unuia dintre schelete de Tenontosaurus. Deoarece greutatea corporală Deinonychus' a fost de numai 70-100 de lire sterline, este puțin probabil ca un specimen solitar ar putea păcăli un 1-4 tone Tenontosaurus, făcând vânătoare grup mai probabil. [22]
Un studiu din 2007 de către Roach și Brinkman a contestat problema vânătorii de grup în Deinonychus, pe baza comportamentului carnivorelor moderne și taphonomy de site - uri care conțin Tenontosaurus. Modern archosaurs (păsări și crocodili ) și dragoni Komodo nu vânează în mod cooperativ. In schimb, ei sunt vânători solitari sau necrofagi . În al doilea scenariu, conflictele dintre animalele din aceeași specie sunt frecvente. Un bun exemplu se referă la dragonii Komodo; în situațiile în care consumă împreună o carcasă, exemplarele mai mari mănâncă mai întâi și le atacă pe cele mai mici care se apropie prea mult. În cazul în care un mic specimen este ucis, este cannibalized . Atunci când aceste informații se aplică la constatările de la site - uri Tenontosaurus, ele par să fie în concordanță cu crocodil Deinonychus' sau comportament Komodo dragon stil. Deinonychus ramasite descoperite în aceste zone provin din exemplare tinere, cu piese lipsă în concordanță cu canibalismul de specimene pentru adulți. [43] Pe de altă parte, au fost descrise urme de multiple Deinonychus paralele și cu măsurători la distanță ale pasului identice, indicând astfel comportamente de cooperare. [44]

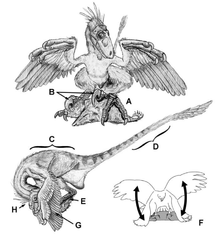
În 2011, Denver Fowler si colegii sai au sugerat o nouă metodă în care Deinonychus și celelalte dromaeosaurids ar captura prada. [41] Acest model, numit „reținere pradă raptor“ (RPR) model, propune ca Deinonychus a ucis prada într - un mod similar cu accipitrid păsărilor de pradă ; Deinonychus ar sări peste prada, ținându - l cu greutatea corpului său și agățându strâns să - l cu ghearele sale curbate. La fel ca accipitridele, dromeosauridul ar începe să se hrănească în timp ce prada era încă în viață, până când a sângerat până la moarte. Acest model se bazează în principal pe morfologia și proporțiile picioarelor dromeosauridelor, care sunt comparabile cu cele ale rapitorilor. Fowler a remarcat că picioarele și picioarele cele mai multe dromaeosaurids sunt foarte similare cu cele ale vulturi și șoimi vrabie , mai ales atunci când a doua gheare extinse și degetul manevrabilitatea sunt luate în considerare. Scurt metatars și forța de prindere a degetelor, cu toate acestea, au fost mai asemănătoare cu cele ale bufnițe . Metoda RPR ar fi , de asemenea , în concordanță cu alte aspecte anatomice ale Deinonychus, cum ar fi morfologia mandibulele și a brațelor. Este posibil ca brațele să fie acoperite cu pene lungi folosite, împreună cu coada rigidă, pentru a echilibra prădătorul în timp ce micșorau prada. Mandibulele sale, care sunt considerate a fi relativ slab, [45] poate să fi fost utilizate într - un ferăstrău mișcare ca modern monitorul Komodo face. [46]
Forța mușcăturii

Estimările de forță mușcătură Deinonychus au fost dezvoltate în 2005, pe baza musculatură mandibular reconstruit. Acest studiu a ajuns la concluzia că , probabil , Deinonychus a avut o muscatura egal cu aproximativ 15% , puterea pe care a aligator american . [45] Un studiu efectuat cinci ani mai târziu , a estimat puterea mușcăturii din nou prin observarea directă impresiile dinților de pe oasele unui specimen de Tenontosaurus. Aceste amprente au venit dintr - o mare specimen, și cu condiția ca prima dovada ca adult mușcături Deinonychus ar putea penetra oase. Pe baza acestor urme, sa ajuns la concluzia că puterea musca a fost mai mare decat estimat anterior, cu o estimare a 4,100-8,200 newtoni , depășind puterea oricărui moderne mamifer carnivor , inclusiv hienă și comparabil cu un aligator. De dimensiuni similare. [47] Oamenii de știință implicați în acest studiu a remarcat , totuși , că amprentele dentare Deinonychus "sunt relativ rare , în comparație cu cele ale theropods oxyphragic , cum ar fi Tyrannosaurus , conducând astfel la probabilitatea ca Deinonychus nu consuma oasele în mod obișnuit, folosind în loc de muscatura sa formidabil la apărare sau pentru a prinde prada. [47]
Funcția membrelor
Spate

Deși gheara este semnul distinctiv al Deinonychus, forma sa variază între exemplare. Specimenul descris de Ostrom în 1969 are o gheară foarte curbată, în timp ce un exemplar din 1976 avea o gheară mai puțin curbată similară celorlalte gheare de pe picior. [30] Ostrom presupus că aceste diferențe ar putea fi din cauza vârstei sau sexul animalelor.
Există dovezi anatomice [1] și urme [48] arată că gheara a avut loc într - o poziție verticală în timp ce dinozaur mers pe a treia și a patra degetele de la picioare.
Ostrom a sugerat că ar putea lovi cu piciorul Deinonychus cu gheara să se rupă pradă în afară. [1] Unii cercetători au sugerat chiar că gheara ar putea gut mari ceratopsids . [49] Alte studii sugerează că ghearele au fost folosite pentru a înjunghia pradă. [50] Manning și colab. (2005) au construit o replică robotică a piciorului posterior dromeosaurid cu ciocuri hidraulice care i-au permis să lovească o carcasă de porc. Testele au arătat că ghearele nu puteau să străpungă carnea decât să o taie. Autorii studiului au sugerat că ghearele au fost mai eficiente ca instrumente de cățărat decât ca instrumente de vânătoare. [51]
Ostrom comparativ cu Deinonychus struț și cassowary . El a menționat că ambele păsări pot provoca răni severe cu gheara degetului secundar mărită. [1] cassowary are gheare acea măsură de 125 mm lungime. [52] Ostrom citat Gilliard (1958) , în care descrie modul în care loviturile livrate de către aceste păsări pot dezmembra sau spinteca un om. [53]
Kofron (1999 și 2003) a studiat 241 de atacuri de casuar documentate și a constatat că un om și doi câini au fost uciși, dar nu au găsit dovezi de dezmembrare sau eviscerare. [54] [55] seriema are de asemenea o a doua gheare extinse, care se folosește pentru dezmembra pradă mici înainte de hrănire pe ea. [56] Un studiu 2011 a sugerat că ghearele vor fi folosite pentru a micsora prada in loc de a dezmembra ei. [57]
Față


În 2002, studiile biomecanice efectuate de Ken Carpenter a confirmat că cel mai probabil functia de membrele anterioare în actul de ruinare a fost de a se agăța de pradă. Osul coracoid masiv și alungit, înzestrat în mod clar cu mușchi puternici la nivelul taliei, întărește această interpretare. [58] Aceleași studii biomecanice indică faptul că Deinonychus nu a putut îndoi brațele la corpul ca păsările.
Studiile efectuate de Phil Senter în 2006 a indicat că Deinonychus ar putea folosi brațele nu numai să se agațe, dar pentru a transporta obiecte aproape de piept. Dacă Deinonychus avea degetele ca niste pene, penele ar fi mișcările brațului limitate ușor. De exemplu, atunci când Deinonychus a extins brațul înainte, mâna rotit într - o poziție de palmier în sus. Acest lucru ar fi făcut ca una dintre aripi să o blocheze pe cealaltă dacă ambele brațe ar fi întinse în același timp. Acest lucru a condus la ipoteza că Deinonychus ar putea deține obiecte la piept cu un singur braț , la un moment dat. Prezența penelor ar fi limitat și mișcarea degetelor; de exemplu, numai al treilea deget al mâinii ar fi putut fi folosit pentru a explora crăpăturile pentru prada mică și numai într-o poziție perpendiculară pe braț. [40] Alan Gishlick , într - un studiu din 2001 privind biomecanica de arme Deinonychus ", a constatat că prezența pene nu ar împiedica capacitatea animalului de a ține pe obiecte. Apucarea ar fi făcută într-o poziție perpendiculară pe aripă, iar obiectele ar fi ținute în ambele mâini într-o poziție de „îmbrățișare”. Aceste descoperiri au fost confirmate ulterior cu studiile lui Carpenter și Senter. [59]
Parsons și Parsons au arătat că exemplare tinere Deinonychus prezintă diferențe morfologice la adulți. De exemplu, brațele minorilor sunt relativ mai lungi decât cele ale adulților, sugerând astfel comportamente diferite. [60] Se referă ghearele piciorului. Parsons și Parsons au sugerat că curbura ghearelor a fost mai pronunțată la exemplarele tinere, permițându-le astfel să urce în copaci înainte de a susține o viață pur terestră a adulților. [61] Aceasta se bazează pe ipoteza că anumite dromaeosaurids mici folosite ghearele picioarelor lor să urce. [51]
Viteză

Dromaeosaurids, în special Deinonychus, sunt adesea reprezentate în cultura populară ca animale foarte repede. Ostrom însuși speculat în descrierea originală că Deinonychus era un animal deosebit de rapid. [10] Cu toate acestea, specimenul originale nu au picioare intacte și estimări ale Ostrom lui femurului lungime au fost dovedit ulterior a fi supraestimate. Într - un studiu ulterior, Ostrom remarcat faptul că relația dintre femur la tibie nu este relevant în determinarea vitezei animalului ca raportul dintre lungimea tibiei la picior. La păsările moderne care aleargă, cum ar fi struții, relația dintre picior și tibie este de .95. În dinozauri analogi, cum ar fi Struthiomimus , relația este 0.68, dar în Deinonychus este doar .48. Ostrom a declarat că singura concluzie logică a fost că Deinonychus nu a fost extrem de rapid în comparație cu alți dinozauri, și cu siguranță nu mai repede decât alergători moderni. [30]
Scurtimea piciorului în raport cu tibia se datorează în parte genoflexiunii metatarsiene , care sunt , de fapt , mai mult , în exemplare tinere. Ostrom a sugerat că metatarsianele scurte au fost legate de funcția ghearelor, pe baza faptului că se scurtează de-a lungul anilor. El a interpretat aceste caracteristici ca sprijin pentru teoria că picioarele din spate erau folosite ca arme. El a sugerat că metatars scurt a redus presiunea asupra tibiei în timpul atacurilor, și , de asemenea interpretat ca atașament musculare neobișnuit animalului a demonstrat că , în timpul unui atac, Deinonychus utilizate alte scopuri decât cele pentru mersul pe jos mușchii. [30]
Într - un studiu pe piste de dinozauri din Canada, Richard Kool a estimat viteza de mers a unei posibile Deinonychus la 10.1 kilometri pe oră. [62]
Ou
Identificarea în 2000 a unui ou Deinonychus probabil în specimenele originale a permis compararea cu alte theropods în ceea ce privește structura, îngrijirea și producția de ouă. În 2006, Grellet-Tinner și Makovicky investigat posibilitatea ca ouăle au fost mâncat de Deinonychus, sau că au fost cu animalul purecoincidence. Primul scenariu a fost discreditat atunci când s-a observat că fragmente de coajă se află între coastele ventrale și picioarele anterioare, mai degrabă decât în regiunea stomacului. S-a mai arătat că s-au format fracturi de coajă în timpul fosilizării. Ipoteza că apropierea dintre ouă și dinozaur a fost întâmplătoare a fost infirmată de faptul că oasele din jurul ouălor erau aproape intacte și nu erau împrăștiate. Cei doi savanți ajuns la concluzia că Deinonychus a fost eclozare ouăle înainte de a fi îngropat, arătând astfel că a avut comportamente materne similare cu cele deduse pentru troodontids și oviraptorids sau că ouăle erau încă în oviduct când animalul a murit. [26]
Examinarea microstructura Confirma ou Deinonychus că a aparținut unei theropod, deoarece împărtășește caracteristici cu cele ale altor theropods, și demonstrează diferențele față de cele ale ornithischians și Sauropozii . Comparativ cu ouă de alte maniraptors , de ou , care se aseamănă Deinonychus din oviraptorids mai mult decât troodontids, deși acesta din urmă grup este mai mult legate de dromaeosaurids. Deși oul a fost prea sfărâmat pentru a-și determina mărimea la nivelul taliei, Grellet-Tinner și Makovicky au estimat că are un diametru de 7 cm pe baza lățimii canalului pelvin al mamei. Acest lucru arată că ouăle de Deinonychus comparabile în mărime cu cele ale oviraptorid Citipati . [26]
Paleoecologie

Dovezi geologice sugerează că Deinonychus locuite luncilor și zone mlăștinoase . [14] Mediul Formațiunii Cloverly și d'Antlers, unde au fost descoperite rămășițele Deinonychus, a constat din paduri tropicale, deltele râurilor , și lagune, similare cu cele moderne Louisiana . [63] [64] Dintre animalele care au trăit în același timp cu Deinonychus erau dinozauri ierbivore precum sauropelta și ornithopoda Zephyrosaurus și Tenontosaurus . În Oklahoma, ecosistemul Deinonychus a inclus marele terapod Acrocanthosaurus , The sauropod Sauroposeidon , crocodil Goniopholis, iar peștele Caiman Lepisosteus . [64] În cazul în care dinții descoperite în Maryland a aparținut Deinonychus ar însemna că a fost , de asemenea , contemporan cu Astrodon și Pricodon nodosaur. [65]
În cultura de masă
I Velociraptor di Jurassic Park e il suo film omonimo furono modellati su Deinonychus , che a quel tempo era stato temporaneamente ribattezzato Velociraptor antirrhopus da Gregory Scott Paul . [19] Sia nel romanzo sia nel film, i resti di Velociraptor vengono trovati in Montana , dove viveva Deinonychus . Il vero Velociraptor infatti viveva nell'Asia centrale. [66] Nel film, la taglia e la forma del muso sono molto più caratteristici di Deinonychus che al Velociraptor vero. [3] [4] Appaiono anche come nemici nel film Alla ricerca della Valle Incantata 7 - La pietra di fuoco freddo . Compaiono anche nel videogioco Jurassic World Evolution ma vengono erroneamente mostrati con una cresta sulla testa.
Note
- ^ a b c d e f JH Ostrom, Stratigraphy and paleontology of the Cloverly Formation (Lower Cretaceous) of the Bighorn Basin area, Wyoming and Montana , in Bulletin of the Peabody Museum of Natural History , vol. 35, 1970, pp. 1-234.
- ^ Mark A. Norell e Makovicky, Peter J., Important features of the dromaeosaurid skeleton II: information from newly collected specimens of Velociraptor mongoliensis , in American Museum Novitates , vol. 3282, 1999, pp. 1-45.
- ^ a b Jody Duncan, The Winston Effect , London, Titan Books, 2006, p. 175, ISBN 1-84576-365-3 .
- ^ a b Robert T. Bakker , Raptor Red , New York, Bantam Books, 1995, p.4 , ISBN 0-553-57561-9 .
- ^ William L. Parsons e Kristen M. Parsons, Further descriptions of the osteology of Deinonychus antirrhopus (Saurischia, Theropoda) , in Bulletin of the Buffalo Society of Natural Sciences , vol. 38, 2009.
- ^ a b GS Paul, Predatory Dinosaurs of the World , New York, Simon and Schuster, 1988, pp. 366 –369, ISBN 0-671-61946-2 .
- ^ Nicolas E. Campione, David C. Evans, Caleb M. Brown e Matthew T. Carrano, Body mass estimation in non-avian bipeds using a theoretical conversion to quadruped stylopodial proportions , in Methods in Ecology and Evolution , vol. 5, n. 9, 2014, DOI : 10.1111/2041-210X.12226/abstract .
- ^ a b WD Maxwell e LM Witmer, New Material of Deinonychus (Dinosauria, Theropoda) , in Journal of Vertebrate Paleontology , vol. 16, n. 3, 1996, pp. 51A.
- ^ Lawrence M. Witmer e William D. Maxwell, The skull of Deinonychus (Dinosauria:Theropoda): New insights and implications , in Journal of Vertebrate Paleontology , vol. 16, n. 3, 1996, pp. 73A.
- ^ a b c d e f JH Ostrom, Osteology of Deinonychus antirrhopus , an unusual theropod from the Lower Cretaceous of Montana , in Peabody Museum of Natural History Bulletin , vol. 30, 1969, pp. 1-165.
- ^ a b c X. Xu, Z. Zhou, X. Wang, X. Kuang, F. Zhang e X. Du, Four-winged dinosaurs from China , in Nature , vol. 421, n. 6921, 2003, pp. 335-340, DOI : 10.1038/nature01342 , PMID 12540892 .
- ^ SH Hwang, MA Norell, Q. Ji e K. Gao, New Specimens of Microraptor zhaoianus (Theropoda: Dromaeosauridae) from Northeastern China , in American Museum Novitates , vol. 3381, 2002, p. 44, DOI : 10.1206/0003-0082(2002)381<0001:nsomzt>2.0.co;2 .
- ^ AH Turner, Makovicky, PJ e Norell, MA, Feather quill knobs in the dinosaur Velociraptor , in Science , vol. 317, n. 5845, 2007, p. 1721, DOI : 10.1126/science.1145076 , PMID 17885130 .
- ^ a b Norell, MA, Makovicky, PJ, Dromaeosauridae , in Weishampel, DB, Dodson, P., Osmólska, H. (a cura di), The Dinosauria , 2nd, Berkeley, University of California Press, 2004, pp. 196 –210, ISBN 0-520-24209-2 .
- ^ a b c MA Norell, Clark, JM, Turner, AH, Makovicky, PJ, Barsbold, R. e Rowe, T., A new dromaeosaurid theropod from Ukhaa Tolgod (Ömnögov, Mongolia) ( PDF ), in American Museum Novitates , vol. 3545, 2006, pp. 1-51, DOI : 10.1206/0003-0082(2006)3545[1:ANDTFU]2.0.CO;2 . URL consultato il 7 luglio 2007 .
- ^ AS Turner, Hwang, SH e Norell, MA, A small derived theropod from Öösh, Early Cretaceous, Baykhangor Mongolia ( PDF ), in American Museum Novitates , vol. 3557, 2007, pp. 1-27, DOI : 10.1206/0003-0082(2007)3557[1:ASDTFS]2.0.CO;2 . URL consultato il 29 marzo 2007 .
- ^ R. Barsbold, Carnivorous Dinosaurs from the Cretaceous of Mongolia , in The Joint Soviet–Mongolian Palaeontological Expedition, Transactions , vol. 19, 1983, pp. 1-117.
- ^ PJ Currie, New information on the anatomy and relationships of Dromaeosaurus albertensis (Dinosauria: Theropoda) , in Journal of Vertebrate Paleontology , vol. 15, n. 3, 1995, pp. 576-591, DOI : 10.1080/02724634.1995.10011250 . ( abstract Archiviato il 27 settembre 2007 in Internet Archive .)
- ^ a b GS Paul, Predatory dinosaurs of the world : a complete illustrated guide , New York, Simon and Schuster, 1988, pp. 358 , ISBN 0-671-61946-2 .
- ^ MJ Benton, Vertebrate Palaeontology (Third Edition) , Blackwell Publishing, 2004, p. 472, ISBN 978-0-632-05637-8 . URL consultato l'11 marzo 2015 (archiviato dall' url originale il 19 ottobre 2008) .
- ^ Robert A. DePalma, David A. Burnham, Larry D. Martin, Peter L. Larson e Robert T. Bakker, The First Giant Raptor (Theropoda: Dromaeosauridae) from the Hell Creek Formation. , in Paleontological Contributions , n. 14, 2015, DOI : 10.17161/paleo.1808.18764 .
- ^ a b DL Brinkman, RL Cifelli e NJ Czaplewski, First occurrence of Deinonychus antirrhopus (Dinosauria: Theropoda) from the Antlers Formation (Lower Cretaceous: Aptian–Albian) of Oklahoma , in Oklahoma Geological Survey Bulletin , vol. 146, 1998, pp. 1-27.
- ^ Z.-Q. Chen e S. Lubin, A fission track study of the terrigenous sedimentary sequences of the Morrison and Cloverly Formations in northeastern Bighorn Basin, Wyoming , in The Mountain Geologist , vol. 34, 1997, pp. 51-62.
- ^ D. Burton, BW Greenhalgh, BB Britt, BJ Kowallis, WS Elliott e R. Barrick, New radiometric ages from the Cedar Mountain Formation, Utah and the Cloverly Formation, Wyoming: implications for contained dinosaur faunas , in Geological Society of America Abstracts with Programs , vol. 38, n. 7, 2006, p. 52.
- ^ TR Lipka, The Affinities of the Enigmatic Theropods of the Arundel Clay Facies (Aptian), Potomac Formation, Atlantic Coastal Plain of Maryland , in Lucas, SG, Kirkland, JI and Estep, JW (a cura di), Lower and Middle Cretaceous Terrestrial Ecosystems , New Mexico Museum of Natural History and Science Bulletin, 14, Albuquerque, New Mexico Museum of Natural History and Science, 1998, pp. 229-234, OCLC 40283894 .
- ^ a b c d G. Grellet-Tinner e Makovicky, P., A possible egg of the dromaeosaur Deinonychus antirrhopus : phylogenetic and biological implications , in Canadian Journal of Earth Sciences , vol. 43, n. 6, 2006, pp. 705-719, DOI : 10.1139/E06-033 .
- ^ a b MA Norell, Gaffney, ES e Dingus, L.,Discovering Dinosaurs in the American Museum of Natural History , New York, Knopf, 1995, pp. 126 –130, ISBN 0-679-43386-4 .
- ^ Ostrom, JH, 1969, "A new theropod dinosaur from the Lower Cretaceous of Montana", Postilla 128 : 1–17
- ^ John H. Ostrom, The Pectoral Girdle and Forelimb Function of Deinonychus (Reptilia: Saurischia) : A Correction , in Postilla, Peabody Museum of Natural History Bulletin , vol. 165, 1974, pp. 1-11.
- ^ a b c d JH Ostrom, On a new specimen of the Lower Cretaceous theropod dinosaur Deinonychus antirrhopus , in Breviora , vol. 439, 1976, pp. 1-21.
- ^ PJ Makovicky e Grellet-Tinner, G., Association between a specimen of Deinonychus antirrhopus and theropod eggshell , in Bravo, AM and T. Reyes (a cura di), First international symposium on dinosaur eggs and babies,Isona i Conca Dellà Catalonia, Spain, 23–26 September 1999 , 2000, pp. 123-128.
- ^ Gerard Grellet-Tinner, Oology And The Evolution Of Thermophysiology In Saurischian Dinosaurs: Homeotherm And Endotherm Deinonychosaurians? , in Papeis Avulsos de Zoologia , vol. 46, n. 1, 2006, pp. 1-10, DOI :10.1590/S0031-10492006000100001 . URL consultato il 7 luglio 2007 (archiviato dall' url originale il 29 settembre 2007) .
- ^ Gregory M. Erickson , Curry Rogers, Kristina, Varricchio, David J., Norell, Mark A. e Xu, Xing, Growth patterns in brooding dinosaurs reveals the timing of sexual maturity in non-avian dinosaurs and genesis of the avian condition ( PDF ) [ collegamento interrotto ] , in Biology Letters , vol. 3, n. 5, 2007, pp. 558-61, DOI : 10.1098/rsbl.2007.0254 , PMC 2396186 , PMID 17638674 . URL consultato il 26 luglio 2007 .
- ^ Fastovsky, DE, Weishampel, DB, Theropoda I: Nature Red in Tooth and Claw , in Fastovsky, DE, Weishampel, DB (a cura di), The Evolution and Extinction of the Dinosaurs , 2nd, Cambridge, Cambridge University Press, 2005, pp. 265-299, ISBN 0-521-81172-4 .
- ^ JH Ostrom,Archaeopteryx and the origin of birds , in Biological Journal of the Linnean Society , vol. 8, n. 2, 1976, pp. 91-182, DOI : 10.1111/j.1095-8312.1976.tb00244.x .
- ^ RT Bakker , The Dinosaur Heresies , Kensington Publishing , 1986, p. 310, ISBN 0-8065-2260-7 .
- ^ Long, JA ; Schouten, P., Deinonychus , in Feathered Dinosaurs: The Origin of Birds , Oxford University Press , 2008, pp. 142-143, ISBN 0-19-537266-2 .
- ^ Dougal Dixon , Fast Hunters , in The Illustrated Encyclopedia of Dinosaurs , Lorenz Books, 2007, pp. 160-161, ISBN 0-7548-1573-0 .
- ^ AH Turner, Makovicky, PJ e Norell, MA, Feather quill knobs in the dinosaur Velociraptor ( PDF ), in Science , vol. 317, n. 5845, 2007, p. 1721, DOI : 10.1126/science.1145076 , PMID 17885130 .
- ^ a b P. Senter, [897:COFFBD 2.0.CO;2 Comparison of Forelimb Function Between Deinonychus And Bambiraptor (Theropoda: Dromaeosauridae) ], in Journal of Vertebrate Paleontology , vol. 26, n. 4, 2006, pp. 897-906, DOI : 10.1671/0272-4634(2006)26[897:COFFBD]2.0.CO;2 .
- ^ a b Denver W. Fowler, The Predatory Ecology of Deinonychus and the Origin of Flapping in Birds , 14 dicembre 2011, DOI : 10.1371/journal.pone.0028964 .
- ^ WD Maxwell e Ostrom, JH, Taphonomy and paleobiological implications of Tenontosaurus – Deinonychus associations , in Journal of Vertebrate Paleontology , vol. 15, n. 4, 1995, pp. 707-712, DOI : 10.1080/02724634.1995.10011256 . ( abstract Archiviato il 27 settembre 2007 in Internet Archive .)
- ^ BT Roach e DL Brinkman, [103:AROCPH 2.0.CO;2 A reevaluation of cooperative pack hunting and gregariousness in Deinonychus antirrhopus and other nonavian theropod dinosaurs ], in Bulletin of the Peabody Museum of Natural History , vol. 48, n. 1, 2007, pp. 103-138, DOI : 10.3374/0079-032X(2007)48[103:AROCPH]2.0.CO;2 .
- ^ Rihui Li, Martin G. Lockley, Peter J. Makovicky, Masaki Matsukawa, Mark A. Norell, Jerald D. Harris e Mingwei Liu, Behavioral and faunal implications of Early Cretaceous deinonychosaur trackways from China , in Naturwissenschaften , vol. 95, n. 3, 2007, pp. 185-91, DOI : 10.1007/s00114-007-0310-7 , PMID 17952398 .
- ^ a b Therrien, F., Henderson, DM and Huff, CB (2005). "Bite me: biomechanical models of theropod mandibles and implications for feeding behavior." pp. 179–237 in Carpenter, K. (ed.), The Carnivorous Dinosaurs . Indianapolis: Indiana University Press.
- ^ DOI : 10.1098/rspb.2010.0794
- ^ a b PM Gignac, PJ Makovicky, GM Erickson e RP Walsh, A description of Deinonychus antirrhopus bite marks and estimates of bite force using tooth indentation simulations , in Journal of Vertebrate Paleontology , vol. 30, n. 4, 2010, pp. 1169-1177, DOI : 10.1080/02724634.2010.483535 .
- ^ Rihui Li, Martin G. Lockley, Peter J. Makovicky, Masaki Matsukawa, Mark A. Norell, Jerald D. Harris e Mingwei Liu, Behavioral and faunal implications of deinonychosaur trackways from China , in Naturwissenschaften , vol. 95, n. 3, 2007, pp. 185-91, DOI : 10.1007/s00114-007-0310-7 , PMID 17952398 .
- ^ Adams, Dawn (1987) "The bigger they are, the harder they fall: Implications of ischial curvature in ceratopsian dinosaurs" pp. 1–6 in Currie, Philip J. and Koster, E. (eds) Fourth symposium on mesozoic terrestrial ecosystems. Tyrrell Museum, Drumheller, Canada
- ^ K. Carpenter, Evidence of predatory behavior by carnivorous dinosaurs , in Gaia , vol. 15, 1998, pp. 135-144.
- ^ a b Phil L. Manning, David Payne, John Pennicott, Paul M. Barrett e Roland A. Ennos,Dinosaur killer claws or climbing crampons? , in Biology Letters , vol. 2, n. 1, 2006, pp. 110-112, DOI : 10.1098/rsbl.2005.0395 , PMC 1617199 , PMID 17148340 .
- ^ SJJF Davies, Ratites and Tinamous , New York, Oxford University Press, 2002, ISBN 0-19-854996-2 .
- ^ ET Gilliard, Living birds of the world , Garden City, NY, Doubleday, 1958.
- ^ Kofron, Chhristopher P. (1999) "Attacks to humans and domestic animals by the southern cassowary ( Casuarius casuarius johnsonii ) in Queensland, Australia
- ^ Christopher P. Kofron, Case histories of attacks by the southern cassowary in Queensland , in Memoirs of the Queensland Museum , vol. 49, n. 1, 2003, pp. 335-338.
- ^ Kent H. Redford e Gustav Peters, Notes on the biology and song of the red-legged seriema (cariama cristata) , in Journal of Field Ornithology , vol. 57, n. 4, pp. 261-269.
- ^ DOI : 10.1371/journal.pone.0028964
- ^ K. Carpenter, Forelimb biomechanics of nonavian theropod dinosaurs in predation , in Senckenbergiana Lethaea , vol. 82, 2002, pp. 59-76, DOI : 10.1007/BF03043773 .
- ^ Gishlick, AD, The function of the manus and forelimb of Deinonychus antirrhopus and its importance for the origin of avian flight , in Gauthier, J. and Gall, LF (a cura di), New Perspectives on the Origin and Early Evolution of Birds , New Haven, Yale Peabody Museum, 2001, pp. 301-318.
- ^ W. Parsons e Parsons, K., Morphology and size of an adult specimen of Deinonychus antirrhopus , (Saurischia, Theropoda) , in Journal of Vertebrate Paleontology , vol. 26, 3 sup., 2006, p. 109A.
- ^ WL Parsons e Parsons, KM, Further descriptions of the osteology of Deinonychus antirrhopus (Saurischia, Theropoda) ( PDF ), in Bulletin of the Buffalo Society of Natural Sciences , vol. 38, 2009, pp. 43-54. URL consultato il 13 marzo 2015 (archiviato dall' url originale il 3 luglio 2010) .
- ^ R. Kool, The walking speed of dinosaurs from the Peace River Canyon, British Columbia, Canada , in Canadian Journal of Earth Science , vol. 18, n. 4, 1981, pp. 823-825, DOI : 10.1139/e81-077 .
- ^ CA Forster, The paleoecology of the ornithopod dinosaur Tenontosaurus tilletti from the Cloverly Formation, Big Horn Basin of Wyoming and Montana , in The Mosasaur , vol. 2, 1984, pp. 151-163.
- ^ a b MJ Wedel e Cifelli, RL, Sauroposeidon : Oklahoma's Native Giant ( PDF ), in Oklahoma Geology Notes , vol. 65, n. 2, 2005, pp. 40-57. URL consultato il 7 luglio 2007 (archiviato dall' url originale il 5 luglio 2008) .
- ^ Z.-Q. Chen e Lubin, S., A fission track study of the terrigenous sedimentary sequences of the Morrison and Cloverly Formations in northeastern Bighorn Basin, Wyoming , in The Mountain Geologist , vol. 34, 1997, pp. 51-62.
- ^ Michael Crichton , Jurassic Park , New York, Alfred A. Knopf, 1990, pp. 117 , ISBN 0-394-58816-9 .
Altri progetti
-
 Wikimedia Commons contiene immagini o altri file su Deinonychus antirrhopus
Wikimedia Commons contiene immagini o altri file su Deinonychus antirrhopus -
 Wikispecies contiene informazioni su Deinonychus antirrhopus
Wikispecies contiene informazioni su Deinonychus antirrhopus
Collegamenti esterni
- ( EN ) Deinonychus antirrhopus , su Fossilworks.org .
- ( EN )
 PBS Eons, The Raptor That Made Us Rethink Dinosaurs , su YouTube , 29 agosto 2019. URL consultato il 28 agosto 2019 .
PBS Eons, The Raptor That Made Us Rethink Dinosaurs , su YouTube , 29 agosto 2019. URL consultato il 28 agosto 2019 .