Tyrannosaurus rex
| Tiranosaur | |
|---|---|
 Reconstrucția scheletului Tyrannosaurus rex | |
| Gama geologică | |
| Starea de conservare | |
Fosil | |
| Clasificare științifică | |
| Domeniu | Eukaryota |
| Regatul | Animalia |
| Sub-regat | Eumetazoa |
| Ramură | Bilateria |
| Superphylum | Deuterostomie |
| Phylum | Chordata |
| Subfilum | Vertebrate |
| Infraphylum | Gnathostomata |
| Superclasă | Tetrapoda |
| Clasă | Sauropsida |
| Subclasă | Diapsid |
| Infraclasă | Arhosauromorpha |
| Superordine | Dinozauria |
| Ordin | Saurischia |
| Subordine | Theropoda |
| Infraordon | Celurosaurie |
| Superfamilie | † Tyrannosauroidea |
| Familie | † Tyrannosauridae |
| Subfamilie | † Tyrannosaurinae |
| Trib | † Tiranosaurini |
| Tip | † Tiranosaur Osborn, 1905 |
| Specii | Tyrannosaurus rex |
| Nomenclatura binominala | |
| † Tyrannosaurus rex Osborn , 1905 | |
| Sinonime | |
Tyrannosaurus rex (al cărui nume înseamnă literalmente „rege șopârlă tirană”), prescurtat adesea în T. rex (diminutiv științific) sau colocvial T-Rex , este o specie de dinozaur teropod care a trăit în Cretacicul târziu , cu aproximativ 70-65 milioane de ani în urmă ( Maastrichtian ), aparținând familiei tiranosauridelor . Denumirea italiană comună pentru acest gen de dinozauri este „tiranosaur”. A trăit în America de Nord , care pe atunci era un continent izolat pe nume Laramidia . Tiranosaurul a fost mult mai răspândit din punct de vedere geografic decât celelalte tiranozauride. Fosilele sale se găsesc într-o varietate de formațiuni care datează din epoca Maastrichtiană din Cretacicul superior , cu aproximativ 70-65 de milioane de ani în urmă. [1] A fost una dintre speciile ultimilor dinozauri numită în mod corespunzător atunci când a avut loc extincția în masă a Cretacic-Paleocen , care a determinat dispariția acestuia.
La fel ca celelalte tiranosauride, Tiranosaurul era un carnivor biped cu un craniu mare, masiv, echilibrat de o coadă lungă și grea. În comparație cu picioarele sale puternice, membrele anterioare ale Tiranosaurului erau scurte, dar relativ puternice și echipate cu două degete cu gheare. Deși depășit ca dimensiune de Spinosaurus , Tiranosaurul a fost cel mai mare dintre tiranosauride și unul dintre cei mai mari și mai puternici prădători terestri cunoscuți.
Cel mai complet exemplar fosil (descoperit în 1992 și denumit „Sue”) măsoară aproximativ 12,3 metri lungime, [2] 4,5 metri înălțime, [3] și se estimează că cântărește până la 6,8 tone [4] , totuși, conform descoperiri foarte recente, artefactul numit „Scotty”, descoperit în 1991, dar recuperat complet în urmă cu doar câțiva ani, din cauza unor dificultăți deosebite în săpătură, s-ar dovedi a fi cel mai mare, cel mai masiv și mai vechi Tiranosaur găsit vreodată. [5] [6] Estimările exacte, luate dintr-un schelet aproape complet, raportează că specimenul ar măsura mai mult de 13 metri în lungime și ar cântări aproximativ 8,8 tone, depășind astfel recordul anterior stabilit de „Sue”. [5] [7] . S-au estimat și alte exemplare mai incomplete: „C. Rex”, descoperit de Jack Horner în 2000, ar fi, de asemenea, mai lung și mai robust decât „Sue”, făcând specia să atingă 14 metri lungime [8] și să conducă din nou animalul care concurează pentru titlul de cel mai mare dinozaur carnivor din toate timpurile.
Unii paleontologi au emis ipoteza că T. rex a fost un prădător în vârful piramidei alimentare a vremii, hrănindu-se cu hadrosauri , ceratopside , anchilosauride și probabil și cu sauropode ; [9] Alții, totuși, au sugerat că animalul era un scavenger . Problema face obiectul unei îndelungate dezbateri în paleontologie , în care majoritatea paleontologilor cred că Tiranosaurul era un carnivor oportunist, adică se hrănea atât cu pradă vie, cât și cu carcase . [10] Se estimează că avea una dintre cele mai puternice maxilare din regnul animal. [11] [12]
De-a lungul anilor au existat numeroase simulări și publicații concepute pentru a estima puterea mușcăturii Tyrannosaurus rex, iar în 2019 au fost efectuate noi studii asupra craniului marelui prădător. Ian N. Cost de la Universitatea Columbia, folosind o echipă variată, după ce a reconstituit o hartă 3D complexă a craniului reptilei și a analizat dinamica acesteia, de asemenea, cu tehnici de inginerie, evaluând setul de țesuturi și ligamente de colagen, a ajuns la concluzia că Tyrannosaurus rex avea o structură craniană rigidă (ca cea a crocodililor și a hienelor) și inflexibilă (precum păsările și șerpii). [13] În plus, analizele au arătat că palatul T. rex era deosebit de rigid și capabil să reziste la solicitări mecanice foarte puternice în timpul mușcăturilor. Toate acestea ar fi garantat reptilei să exercite o putere de 64.000 N. în mușcătură, egală cu 7,1 tone, făcându-l capabil să zdrobească orice tip de oase și să se hrănească cu măduva hadrosaurilor și ceratopsidelor . [14] Conform diferitelor experimente, Tyrannosaurus rex ar fi putut distruge caroseria unei mașini. [15] În orice caz, puterea dezlănțuită de fălcile T. rex face din acest rex mare oficial cel mai puternic animal terestru mușcat înregistrat vreodată; gândiți-vă doar că un exemplar adult ar fi putut provoca o pradă (cum ar fi un Triceratops ) o rană de 60 cm lungime și 30 de adâncime, ceea ce înseamnă că, odată ajunsă la prada din spatele T.rex, ar fi aterizat imediat. [16]
Au fost găsite peste 50 de rămășițe fosile de Tiranosaur , inclusiv câteva schelete aproape complete. Cel puțin unul dintre aceste exemplare păstrează urme de țesut moale și proteine . Abundența artefactelor a făcut posibilă cercetarea detaliată a aspectelor biologiei Tyrannosaurus , inclusiv a celor legate de biomecanică și creșterea acesteia. Cu toate acestea, comportamentele sale de vânătoare, gradul său de inteligență , fiziologia și alte detalii ale vieții, cum ar fi viteza maximă în timpul unei curse, sunt încă subiecte de dezbatere. Sa taxonomie este , de asemenea , un subiect de controverse, deoarece unii cercetători considera tarbozaur Bataar , găsit în Asia, oa doua specie Tyrannosaurus, în timp ce alții clasifica ca un gen separat. Alte genuri de tiranozauride nord-americane, descrise la momentul descoperirii lor ca fiind noi taxoni , au fost ulterior sinonime cu Tiranosaurul .
Etimologie
« Tiranosaurul este universal cunoscut sub întregul nume specific, Tyrannosaurus rex , care îl ridică deasupra genului anonim al celorlalte specii fosile, aproape toate cunoscute doar sub numele generic, fără a menționa vreodată specia. Menționează cineva vreodată Velociraptor mongoliensis ? Nu, există doar „ velociraptorul ” (sau, mai rău, „rapitorul”). ” |
| ( Andrea Cau (2012) [17] ) |
Numele științific al tiranosaurului se datorează lui Henry Fairfield Osborn și amintește în ambele părți ale sale o idee de dominație: Tiranosaurul este sinteza grecului τύραννος ( týrannos : „ tiran ”) și σαῦρος ( sâuros : „șopârlă”); rex este latină pentru „ rege ”. Prin urmare, denumirea sa științifică completă înseamnă „Tiranul Regelui Șopârlă”.
Anatomie
„ Tiranosaurul a unit în sine dimensiunile unui elefant, violența unui tigru și dinții unui rechin” [18]

Tyrannosaurus rex a fost unul dintre cele mai mari carnivore terestre din toate timpurile; cel mai complet exemplar, și considerat până de curând, cel mai mare descoperit (inventariat ca FMNH PR2081 și cunoscut sub porecla de „Sue”), are un schelet care măsoară 12,3 metri lungime, [2] și are 5,5 metri înălțime. [3] Estimările greutății taliei variază de la 4,5 tone[19] [20] la peste 7,2, [21] deși cele mai recente estimări o situează între 5,4 și 6,8 tone. [4] [22] [23] [24] Un studiu realizat în 2011 a stabilit că greutatea maximă a Sue este estimată a fi între 9,5-18,5 tone. Cu toate acestea, autorii studiului au afirmat că estimările lor minime și maxime se bazează pe modele cu marje substanțiale de eroare și că au considerat estimările fie prea „subțiri”, „grase”, fie „disproporționate”. [2] Packard și colab. (2009) au aplicat metodele utilizate pentru a estima greutatea dinozaurilor pentru a estima greutatea elefanților și au constatat că aceste metode au produs supraestimări; prin urmare, greutatea Tyrannosaurus rex poate fi mai mică decât cea estimată anterior. [25] Alte estimări au concluzionat că exemplarele mai mari de Tiranosaur cântăresc în jur de [26] sau peste 9 tone. [2] [27] [28]
În martie 2019, o echipă de paleontologi, condusă de Scott Pearson, a constatat că descoperirea numită „Scotty” (RSM P2523.8), descoperită în 1991 și recuperată complet în urmă cu doar câțiva ani, din cauza dificultăților deosebite în excavare , s-ar dovedi a fi cel mai mare Tiranosaur găsit vreodată. [5] [6]

Descoperirea este acum considerată completă (aproximativ 70%) și din studiul picioarelor din spate și a femurului, echipa de cercetare a reușit să stabilească cu certitudine că „Scotty” avea mai mult de 13 metri lungime și cântărea 8.845 de tone, depășind astfel record stabilit de „Sue” [5] [6] ; În special, robustețea animalului este surprinzătoare, făcându-l cel mai masiv prădător terestru descoperit vreodată [6], de asemenea, Scotty se dovedește a fi cel mai vechi Tyrannosaurus găsit vreodată care a trăit timp de aproximativ treizeci de ani. [6]
Alți tiranozauri "foarte mari", deși sunt incomplet, sunt specimenul "MOR 008" a cărui greutate a fost estimată la 7 tone pe 14 metri lungime. [29] și descoperirea poreclită „C-REX”, ale cărei estimări, efectuate de Jack Horner, ar atribui lui T. Rex o greutate apropiată de 9 tone și o lungime mai mare de 14 metri. [30]
Cu toate acestea, acestea sunt estimări efectuate pe rezultate incomplete, în special deduse din dimensiunile membrelor inferioare.
La fel ca alte teropode , gâtul Tiranosaurului era curbat în formă de „S”, dar era remarcabil pentru lungimea limitată și musculatura care îi susținea capul masiv. Membrele anterioare au fost prevăzute cu doar două degete, [31] cu un metacarpian diminuat suplimentar reprezentând un al treilea deget rezidual. [32] În schimb, membrele posterioare erau printre cele mai lungi în raport cu corpul oricărui alt teropod. Coada era lungă și robustă, cântărind o jumătate de tonă, conținând uneori mai mult de 40 de vertebre, pentru a echilibra greutatea capului și a pieptului masiv. Multe dintre oase erau goale, explicate ca ușurând animalul fără pierderea forței fizice. [31]
Cap

Capul ca atare era excepțional de masiv, lung de până la 1,5 metri [33] și, în comparație cu alte teropode anterioare și contemporane, avea o formă puternic modificată. Zona posterioară a craniului a fost mult mărită, în timp ce botul s-a îngustat în corespondență cu nările. În consecință, ochii au putut avea un câmp vizual excelent în regiunea anterioară și anterioară / inferioară, oferind animalului o viziune stereoscopică considerată superioară celei a altor specii contemporane. [34] [35]
Oasele nazale, fuzionate împreună, au oferit animalului o structură craniană mai masivă și mai rezistentă. Multe oase erau pneumatice, ca cele ale păsărilor, permițând astfel reducerea greutății și o flexibilitate mai bună. Aceste caracteristici indică o remarcabilă putere de mușcătură care trebuie să fi fost cu mult superioară tuturor celor non-tiranozauride. [11] [36] [37]
Gura T. rex avea treizeci de dinți în arcada superioară și douăzeci și opt în cea inferioară, caracterizată prin heterodonție ridicată (diferență de formă). [31] [38] Lungimea variază de la zece la treizeci de cm. În general, au o secțiune transversală ovală și au o margine fin zimțată. Premaxilarele superioare, pe de altă parte, sunt apropiate și au o secțiune în formă de D. Dinții rămași au fost robusti, mai distanțați și cu nervuri de întărire, îndoite înapoi cu o formă generală de banană care le-a împiedicat să se rupă în timpul mușcăturii și în timp ce sfâșierea cărnii. [39] Dimensiunile dinților ar putea ajunge până la treizeci de cm lungime, inclusiv rădăcina; au fost, prin urmare, cel mai mare dintre dinozaurii carnivori. [3]
Mușcătura tiranosaurului a fost puternică, similară în putere cu cea a actualului crocodil marin cu o estimare de 36.000 - 53.000 N. Cu toate acestea, nu a putut rivaliza cu cea a crocodililor mari, care l-au depășit de mai multe ori în putere și presiune. [40] Cu toate acestea, această caracteristică i-a permis să străpungă și să rupă oasele prăzilor mari, după cum reiese din urmele lăsate de dinți pe unele rămășițe fosile. Maxilarul conform unei simulații făcute de Gregory Erickson, de la Florida State University și Paul Gignac, de la Universitatea de Stat din Oklahoma și publicat în Scientific Reports, ar putea exercita o mușcătură de aproximativ 36.000 N, care a fost apoi ridicată la 53.000 N în studiile ulterioare. Presiunea a fost de ordinul de 700 kg / cm2. Cu toate acestea, această forță a fost extrem de mică decât cea a crocodililor mari, cum ar fi Purussaurus sau Rhamposuchus [41][42] [43] . După cum afirmă Gignac, datorită acestei abilități "T rex a reușit să folosească mai mult carcasele dinozaurilor mari cu coarne și dinozaurii cu coadă de rață, ale căror oase erau bogate în minerale și măduvă".
În 2019, au fost efectuate noi studii asupra craniului și a mușcăturii T. rex; Ian N. Cost (Departamentul de Patologie și Științe Anatomice al Universității Columbia) în fruntea unei echipe variate de cercetători, după ce a reconstituit o hartă 3D complexă a craniului reptilei și a analizat dinamica acesteia și cu tehnici de inginerie, a atins un rol important concluzii: mușcătura Tyrannosaurus rex ar putea elibera forța de 64.000 N. egală cu 7,1 tone, de aproximativ patru ori mai mult decât crocodilul de mare. Acest lucru se datorează faptului că structura osoasă a craniului T. rex era de tip rigid, ca și crocodilii și hienele actuale, și nu era flexibilă, așa cum se teoretiza în deceniile anterioare; în special, palatul reptilei era foarte rigid, capabil să amortizeze tensiunile mecanice de mestecat și presiunea mușcăturii sale ar fi putut distruge o mașină. T. rex a fost cel mai puternic animal terestru care a mușcat vreodată. [16]
Piele: solzi sau pene
Deși nu există dovezi directe pentru prezența penajului în Tiranosaur , mulți paleontologi la începutul celui de-al doilea deceniu al secolului XXI, pe baza prezenței la specii înrudite de dimensiuni similare, [44] cred că este probabil că a avut un corp acoperit.de pene sau cel puțin pe părți ale corpului. [45] . Mark Norell de la Muzeul American de Istorie Naturală a declarat că, în ciuda absenței dovezilor directe, motivul pentru care credem că T. rex a fost pene pentru cel puțin o etapă a vieții sale este același cu cel pentru care credem că australopitul Lucy avea păr . [44]
Primele indicații care sugerează prezența penajului în tiranozauroizi au provenit din specia Dilong paradoxus , descoperită în formațiunea Yixian din China (2004). Scheletul său fosilizat păstrează structuri filamentoase care sunt recunoscute în mod obișnuit ca fiind precursorii penelor. [46] Întrucât până la această descoperire, toate amprentele cutanate uriașe tiranosauroide au arătat semne de solzi, cercetătorii care studiau Dilong au presupus că penele erau prezente la speciile adulte din speciile mai mici și la puii din speciile mai mari, doar pentru a fi pierdute la maturitate. [46] Descoperirile ulterioare și ulterioare au sugerat că anumite tiranozauride uriașe adulte aveau și pene (cum ar fi Yutyrannus huali ) , punând astfel la îndoială ipoteza că prezența penajului era legată de mărime. [47]
Descoperirea lui Yutyrannus huali (aproximativ 9 metri lungime) în 2012, a evidențiat prezența penelor pe diferite părți ale corpului, sugerând astfel că întregul corp era acoperit cu ele [47] și acest lucru a mutat atenția asupra posibilității ca și mai mari tiranozauridele, precum T. rex , ar putea fi acoperite cu ele.
Studiile publicate în 2017 (de asemenea de Biology Letters [48] și National Geographic [49] ), efectuate de paleontologii Peter Larson și Jack Persons, ar oferi dovezi care susțin teza conform căreia T. rex a fost acoperit cu solzi, precum reptilele de astăzi, și nu din pene; de fapt, conform acestui studiu bazat pe amprentele pielii unui tiranosaur, numit de cercetători „Wyrex”, imprimat pe unele roci, amprente ale unor părți ale pelvisului, cozii și gâtului, absența perilor sau a penelor și a unui formare tipică solzoasă reptiliană. [50]
De fapt, corelația inerentă penajului, între Tyrannosaurus rex nord-american și Yutyrannus huali asiatic, în lumina descoperirilor noi și evidente ale pielii solzoase, evidențiată pe diferite genuri de tiranosauride cretacice târzii , precum Gorgosaurus , Tarbosaurus și același T. rex , în prezent se clatină dacă nu cade complet. Comparația directă între aceleași părți ale corpului lui Yutyrannus și celelalte tiranozauride nord-americane, arată discrepanța clară dintre descoperiri: unde urme de penaj apăruseră pe Yutyrannus , nu există dovezi pentru T. rex [51] Toate acestea a sugerat paleontologilor că aceste animale au luat diferite căi evolutive. [51] Cercetătorii sunt convinși că T. rex a pierdut complet sau aproape complet aceste structuri din cauza dimensiunii, termoreglării corpului și habitatului. [49]
Prin urmare, după un deceniu de discuții, paleontologii (inclusiv Philip J. Currie și Robert T. Bakker [48] ) sunt de acord cu privire la reconstrucția aspectului T. rex , la o interpretare mai tradițională și „canonica”, în absența unor dovezi reale privind prezența proto-pene. [50]
În plus, descoperirea unui tegument facial solzos, într-un specimen de Daspletosaurus , ar indica faptul că majoritatea tiranosauridelor , inclusiv Tiranosaurul , dețineau organe senzoriale tegumentare faciale dezvoltate, posibil utilizate pentru contact, modularea mișcărilor maxilarului, percepția temperaturii și detectarea pradă. Cântarele mari plate și largi prezente pe bot (asemănătoare cu cele ale crocodililor ) au garantat, de asemenea, o protecție suplimentară pe bot în timpul capturării prăzii și în timpul luptelor intraspecifice. [52] [53] [54]
Dimorfism sexual

Între anii optzeci și nouăzeci, când a început să aibă un număr suficient de mare de dovezi fosile pentru a efectua analize pe baze statistice , a început să se propună diferențierea sexuală a tiranozaurilor pe baza mărimii. De fapt, există două „tipuri” distincte de schelete de tiranosauri adulți. Un tip este masiv și mare, în timp ce al doilea tip este mai slab și de dimensiuni mai mici. Exemplul izbitor al primei categorii este scheletul numit SUE (în onoarea descoperitorului Sue Hendrickson), cel mai complet (și, de asemenea, cel mai mare, cu o lungime de 13 metri) găsit vreodată.
Mai mulți paleontologi au propus că, la fel ca în păsările de pradă de astăzi, cele mai mari și mai masive schelete aparțineau tiranosaurilor femele, în timp ce cele mai ușoare aparțineau masculilor.
Ipoteza părea să se reflecte în diferența dimensională a chevronului primei vertebre caudale. Osul în cauză a apărut mai dezvoltat pe exemplarele mai mici și mai puțin pe cele mai mari. S-a emis ipoteza că dimensiunea mică era legată de necesitatea de a permite ouălor să treacă mai ușor din tractul reproductiv. [55] Pentru a întări teoria, a fost adus un vechi studiu realizat de Romer asupra crocodililor , care s-ar fi dovedit a fi eronat. [56]
Descoperirea acestei erori de bază în studiul crocodilului, împreună cu descoperirea unui chevron complet dezvoltat în scheletul SUE, au dus la o revizuire radicală a acestor ipoteze în ultimii ani. Singurul individ al cărui sex a fost identificat cu certitudine este MOR 1125 (așa-numitul B-rex găsit în estul Montanei a cărui măduvă ar putea fi analizată, parțial conservată), descoperită de Jack Horner și Mary Schweitzer în 2000. Specimenul nu era deosebit mare, nici nu era foarte complet. Cu toate acestea, o privire în interiorul unuia dintre femururi în 2007 a arătat că era un exemplar unic. În interior erau țesuturi moi, împreună cu structuri asemănătoare vaselor de sânge. Acestea erau similare cu vasele de sânge ale unui struț, oferind dovezi suplimentare ale relației dintre păsări și dinozauri. Dintr-o analiză a țesuturilor conservate s-a dovedit a fi o femeie, care probabil a murit la vârsta de 16 ani, acum aproximativ 68 de milioane de ani. S-a constatat că o parte este țesutul medular specializat, care se găsește numai la păsările femele în timpul ovulației și servește la furnizarea de calciu pentru formarea cojii de ou. [57]
În prezent, nu există, prin urmare, dovezi clare care să indice existența unui dimorfism legat de sexul indivizilor. Având în vedere vasta difuzie teritorială a speciei, este posibil ca diferențele de mărime să fie atribuite variabilității condițiilor de mediu (temperaturi locale, disponibilitate mai mare sau mai mică de hrană etc.). O altă posibilitate este pur și simplu ca cele mai mari exemplare să fie și cele mai vechi. [31]
Postură

Portrete moderne din muzee, artă și film arată T. rex cu corpul paralel cu solul, cu coada extinsă în spatele corpului pentru a echilibra capul. [58]
Vechile reconstrucții, care au durat până în anii optzeci, arată animalul ca un fel de „trepied viu”, aproape ridicat pe picioarele din spate și cu coada care acționează ca un al treilea punct de sprijin, târându-se pe sol. Acest tip de reconstrucție, care în anumite privințe poate fi considerat „antropomorfizat” se datorează indirect lui Joseph Leidy , care și-a reasamblat astfel Hadrosaurus foulkii în 1865 (care a fost primul dinozaur care a fost descris ca biped). [59] Convins că T. rex și-a asumat o postură similară, Henry Fairfield Osborn a întărit acest punct de vedere adoptând teoria și prezentând, în 1915 , primul schelet complet de tiranosaur, asamblat în acest mod la Muzeul de Istorie Naturală din New York. [60]
Cu toate acestea, începând cu anii șaptezeci , s-a înțeles că această postură este absolut nerealistă, deoarece ar fi cauzat slăbirea și dislocarea a numeroase articulații (dintre care cel mai important care ar fi afectat ar fi fost gâtul și pelvisul). [61] Reconstrucțiile anterioare, care deveniseră foarte populare și care inspiraseră numeroase filme și reproduceri, au fost astfel înlocuite cu altele mai moderne care reprezintă postura Tyrannosaurus rex cu corpul aproape paralel cu solul și coada extinsă pentru a echilibrează greutatea capului. [58]
Membrele anterioare

Când a fost găsit primul schelet Tyrannosaurus rex , humerusul era singurul element cunoscut al membrelor anterioare. [62] Osborn a reconstruit tiranosaurul în 1915 luând membrele de la un alosaur (mai lung și cu trei degete). [63]

În 1914, însă, Lawrence Lambe propusese că era mai potrivit să-l echipăm cu anexe similare cu cele ale lui Gorgosaurus , având în vedere afinitatea mai mare a celor două specii. [64] Ceea ce arătau de fapt a fost un mister până în 1989 . În acel an a fost descoperit așa-numitul „Wankel rex” (specimenul MOR 555), completat cu membrele anterioare și aspectul lor a fost sigur, confirmând ipoteza lui Lambe.[65] [66] [67]
Examinarea scheletală a aparatului brahial [68] sugerează că aceste membre nu pot fi considerate doar vestigiale , spre deosebire de ceea ce a ipotezat Paul în 1988. Oasele au suprafețe mari pentru atașarea mușchilor robusti.
Dezbaterea despre care ar putea fi funcția lor rămâne deschisă. Unii paleontologi au propus ca animalul să-l folosească pentru a aduce bucăți de carne la gură. Ipoteza este probabil de eliminat. Îmbinarea cotului și a umărului permite mișcări într-un arc de maximum 40 / 45º, insuficient în acest scop. Pe de altă parte, este posibil ca el să-și fi folosit membrele pentru a-și menține partenerul nemișcat în timpul copulării [69] sau pentru a ține ferm și a limita mișcările prăzii. [68] O ipoteză mai recentă este că acestea aveau funcția de a ajuta animalul să se ridice, așezând palmele pe pământ și aplicând presiune, dându-i astfel o mică apăsare pentru a-l face să se ridice. [61] .
Taxonomie
Tyrannosaurus este genul de tip al superfamiliei Tyrannosauroidea , al familiei Tyrannosauridae și al subfamiliei Tyrannosaurinae; pe scurt, reprezintă standardul prin care paleontologii decid să includă alte specii în grup. Alți membri ai subfamiliei tiranosaurină includ Daspletosaurul nord-american și Tarbosaurul asiatic, [70] [71], ambele fiind uneori sinonime cu Tiranosaurul . [58] In trecut, tyrannosaurids au fost considerate a fi descendenții theropods gigant în vârstă, precum megalosauroids sau carnosaurs , dar cercetările mai recente indică faptul că acestea sunt derivate din coelurosaurs . [72]

În 1955, paleontologul sovietic Evgeny Maleev a numit o nouă specie descoperită în Mongolia ; Tyrannosaurus bataar . [73] În 1965, această specie a fost redenumită Tarbosaurus bataar . [74] În ciuda acestui fapt, multe analize filogenetice arată că Tarbosaurus bataar este cel mai înrudit taxon cu T. rex , [71] și a fost adesea considerată o specie asiatică a acestuia. [72] [75] [76] Studii recente asupra craniului Tarbosaurus bataar au arătat că acesta era mult mai slab decât cel al T. rex și că distribuția forței mandibulare în timpul unei mușcături ar fi mai asemănătoare cu cea a lui Alioramus , un alt Tiranosaur asiatic. [77] O analiză cladistică ulterioară a sugerat că Alioramus , și nu T. rex , era cel mai apropiat taxon cu Tarbosaurus , justificând astfel separarea lor. [70]
Alte tiranosauride descoperite în aceleași formațiuni ca T. rex au fost clasificate mai întâi ca taxoni separați, inclusiv Aublysodon și Albertosaurus megagracilis , [58] ultima denumită Dinotyrannus megagracilis în 1995. [78] În prezent, aceste fosile sunt considerate a aparține tânărului T. rex . [79] Un craniu mic, dar aproape complet descoperit în Montana care măsoară 60 cm ar putea fi o excepție. Questo teschio fu in origine classificato nel 1946 come un Gorgosaurus , [80] ma fu dopo riferito a un nuovo genere, Nanotyrannus . [81] Le opinioni sono divise sulla validità di Nanotyrannus ; molti paleontologi lo considerano un giovane T. rex . [82] Ciononostante, i due generi mostrano piccole differenze, come il numero di denti più elevati in Nanotyrannus , conducendo così alcuni a raccomandare la separazione dei due fino a che ulteriori scoperte non chiariscano la situazione. [71] [83]
Questo cladogramma è basato sull'analisi filogenetica condotta da Loewen et al. nel 2013. [84]
| Tyrannosauridae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Sinonimi

Sono stati riportati i seguenti sinonimi :
- Albertosaurus megagracilis Paul , 1988
- Aublysodon molnari Paul , 1988
- Aublysodon molnaris Paul , 1988
- Dinotyrannus megagracilis Paul , 1988
- Dynamosaurus imperiosus Osborn , 1905
- Manospondylus gigas Cope , 1892
- Nanotyrannus lancensis Gilmore, 1946
- Stygivenator molnari Paul , 1988
- Tyrannosaurus imperiosus Osborn , 1905 [85]
Paleobiologia
Origine asiatica
L'origine evolutiva del Tyrannosaurus è piuttosto intricata e non del tutto chiara. Nel 2016 , un team di studiosi guidati dal paleontologo Steve Brusatte dell' Università di Edimburgo , in Scozia , ha avanzato l'ipotesi che il Tyrannosaurus avesse origini asiatiche. [86] Secondo le teorie dei paleontologi scozzesi, gli antenati dei tyrannosauridi , e di tutti gli altri dinosauri teropodi , erano già presenti sul supercontinente Pangea , circa 200 milioni di anni fa quando il continente si separò dando origine ai continenti i vari dinosauri si adattarono ai loro rispettivi habitat. Questo spiega anche perché i tyrannosauridi nativi del Nord America, come gli albertosaurini abbiano una costituzione diversa da quelle di altri tyrannosauridi più evoluti. Ma circa 70 milioni di anni fa, Asia e Nord America si avvicinarono formando un ponte di terra che univa i due continenti, permettendo ad animali asiatici di "trasferirsi" in Nord America. Ci sono prove che il Tyrannosaurus avesse origini asiatiche, difatti è strettamente imparentato con tyrannosauridi asiatici quali Tarbosaurus e Zhuchengtyrannus . L'arrivo del Tyrannosaurus in Nord America, circa 67 milioni di anni fa, coincide, inoltre, con la graduale estinzione dei tyrannosauridi nativi americani come Albertosaurus e Gorgosaurus . Si pensa che il Tyrannosaurus abbia avuto un ruolo rilevante nella loro estinzione, spodestando i vecchi predatori e salendo in cima alla catena alimentare, come una vera e propria specie invasiva o aliena . [86]
Sviluppo

L'identificazione di certi esemplari come T. rex giovani ha permesso agli scienziati di documentare i cambiamenti ontogenetici della specie, stimare la durata della vita, e determinare il tasso di crescita. Si stima che l'esemplare più piccolo conosciuto ( LACM 28471) avesse in vita una massa corporea di soli 30 chili, mentre è probabile che il più grande ( FMNH PR2081 "Sue") pesasse 5400 chili. Analisi istologiche sulle ossa dell'esemplare LACM 28471 mostrano che morì a due anni, mentre quelle di "Sue" mostrano che morì a 28 anni, probabilmente vicino alla vita massima per la specie. [87]

Ulteriori studi istologici mostrano che gli esemplari giovani pesavano meno di 1800 chili fino a 14 anni, quando la massa corporea incrementava considerevolmente. Durante questa rapida fase di crescita, un giovane T. rex avrebbe aumentato il proprio peso corporeo di 600 chili all'anno per i successivi quattro anni, rallentando verso a 18 anni. [4][88] Un ulteriore studio corroborò l'ultimo, ma scoprì che il tasso di crescita era più veloce, sui 1800 chili all'anno. Ciò indicò che il tasso di crescita di T. rex era tipico d'animali della sua taglia. [2] Il rallentamento del tasso di crescita dopo i 18 anni potrebbe indicare la maturità fisica, un'ipotesi sostenuta dalla scoperta del midollo fossilizzato nel femore d'un esemplare di 16-20 anni scoperto in Montana ( MOR 1125, soprannomminato "B. rex"). Il tessuto midollare si trova solo negli uccelli femmina durante l' ovulazione , indicando così che l'esemplare era sessualmente maturo. [89] Ulteriori studi indicano che questo esemplare morì a 18 anni. [90] Gli altri tirannosauridi mostrano cambiamenti simili ma a livelli inferiori.[91]
Più del 50% degli esemplari di T. rex scoperti sembrano essere morti entro sei anni dopo raggiunta la maturità sessuale, un modello presente in altri tirannosauri e in certi uccelli e mammiferi moderni di taglia grande. Queste specie sono caratterizzate da tassi di mortalità infantile elevati, con un decremento di mortalità negli adolescenti. La mortalità si eleva di nuovo dopo la maturità sessuale, parzialmente dovuta alla concorrenza per la riproduzione. Uno studio suggerisce che la rarità dei fossili di esemplari giovani di T. rex possa essere dovuta a tassi di mortalità infantili bassi; animali di quell'età, non morendo in grandi numeri, non si fossilizzavano. Questa rarità però può anche essere dovuta a una documentazione di fossili incompleta o alla predilezione dei collezionisti di fossili a scartare esemplari piccoli a favore di quelli più spettacolari.[91] In una conferenza del 2013, Thomas Holtz Jr. suggerì che i dinosauri vivessero velocemente e morissero precocemente a causa della loro rapidità riproduttiva, diversamente dai mammiferi il cui arco vitale ha una maggiore estensione perché prendono più tempo per riprodursi.[92] Anche Gregory S. Paul scrisse che il Tyrannosaurus si riproduceva rapidamente e moriva precocemente, ma attribuì il loro breve arco di vita all'esistenza pericolosa che conducevano. [93]
Diffusione
Charles Marshall, direttore del Museo di Paleontologia dell' Università della California , ha stimato in 2,5 miliardi il numero totale di T. rex vissuti e in 20.000 il numero massimo di esemplari presenti sulla terra nello stesso momento (valori medi). Tale cifra è il risultato di un calcolo che considera - tra l'altro - che la specie è vissuta per un periodo compreso tra 1,2 e 3,6 milioni di anni, che la vita media era di circa 28 anni e che quindi le generazioni totali sono state mediamente 127.000. Conoscendo l'areale di diffusione (l'area del Nord America occidentale attualmente occupata dalle Montagne Rocciose e le Grandi Pianure centrali ) ha anche stimato che ogni esemplare avesse mediamente un territorio di 110 km² [94] [95] .
Tessuto molle
Nel 2005 fu annunciato da Science che tessuto molle era stato recuperato nella cavità midollare d'un femore di T. rex da parte del North Carolina State University . [96] Nominato "Museum of the Rockies specimen 1125" (MOR 1125), l'esemplare fu scoperto presso la formazione Hell Creek, e conteneva sia vasi sanguigni flessibili biforcuti che tessuto della matrice ossea. Entrambi contenevano microstrutture simili a cellule . Attualmente non ci sono spiegazioni concrete per la conservazione di questo materiale. [97] Se di seguito si scoprirà che le microstrutture appartengono definitivamente all'esemplare in questione, le proteine superstiti potrebbero essere utilizzate per ipotizzare indirettamente il contenuto del dinosauro in questione, dal momento che ogni proteina è prodotta da un gene specifico. Dopo questa scoperta sono stati scoperti materiali simili a tessuti in due T. rex e in un adrosauro. [96] Ricerche su alcuni di questi tessuti indicano che gli uccelli sono i parenti più stretti del T. rex , rispetto a qualsiasi altro organismo moderno. [98] Studi pubblicati in Science nel 2007 evidenziarono che tracce di collagene trovate nelle ossa di T. rex mostrano somiglianze con quelle di polli, rane e tritoni. [99] Studi ulteriori del 2008 su tessuti molli di T. rex ne confermarono la parentela con gli uccelli moderni. [100]
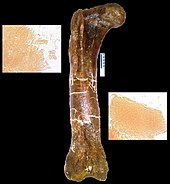
Il presunto tessuto molle fu messo in dubbio nel 2008 da ricercatori dell' University of Washington , che sostennero come il contenuto del femore dell'esemplare di T. rex consistesse in un biofilm prodotto da batteri infiltrati in cavità una volta occupate da cellule. [101] Scoprirono che le strutture inizialmente identificate come frammenti di cellule (a causa della presenza di ferro ) erano invece framboidi (sfere minerali che contengono ferro). Sfere simili sono state scoperte in altri fossili di vari periodi, inclusi in un' ammonite . Nell'ammonite le sfere furono trovate in luoghi dove il ferro che contenevano non poteva avere alcuna correlazione con la presenza di sangue. [102] Tali affermazioni furono contestate per il fatto che non ci sono prove documentate di biofilm che produca tubi cavi e biforcuti come quelli trovati nell'esemplare di T. rex . [103]
Nel 2016 , un team di paleontologi dell' Università della Carolina del Nord ha pubblicato la scoperta di un esemplare di Tyrannosaurus , risalente a 68 milioni di anni fa, morta mentre era incinta . [104] I ricercatori, coordinati dalla paleontologa Mary Schweitzer, hanno individuato nel suo femore un tessuto molto particolare, detto osso midollare, che si trova anche nelle femmine degli uccelli soltanto durante il periodo della deposizione delle uova , e serve a fornire calcio per la formazione dei gusci delle uova. E, in teoria, i resti di questo esemplare potrebbero contenere DNA. [104]
Termoregolazione
Il metodo di termoregolazione del T. rex non è chiaro. Come molti dinosauri, fu prima ritenuto d'avere un metabolismo ectotermico tipico dei rettili a sangue freddo. Questo ragionamento fu contestato negli anni sessanta, durante il cosiddetto "rinascimento dei dinosauri". [105] [106] Fu infatti proposto che T. rex fosse endotermico , indicando così uno stile di vita molto attivo. [20] In seguito, vari paleontologi hanno congetturato sull'attitudine del T. rex di regolare la propria temperatura corporea. Evidenze istologiche d'un elevato tasso di crescita negli esemplari giovani di T. rex potrebbero sostenere l'ipotesi d'un metabolismo elevato. Come in mammiferi e uccelli, lo sviluppo del T. rex si determinava principalmente all'interno dello stadio giovanile, in contrasto ad uno sviluppo più esteso lungo l'arco vitale, osservato nella maggior parte degli altri vertebrati.[88]
Le proporzioni degli isotopi d'ossigeno nelle ossa fossilizzati sono spesso usate per determinare le temperature in cui le ossa furono deposte. In un esemplare, le proporzioni degli isotopi nelle ossa da parti diverse del corpo indicano una differenza di temperatura non più elevata di 4-5 °C tra le vertebre del torace e la tibia . Ciò condusse alla conclusione che T. rex fosse infatti omeotermico , avendo un metabolismo intermediato tra quello dei rettili ectotermici ei mammiferi endotermici. [107] Certi scienziati hanno notato che le proporzioni degli isotopi d'ossigeno nei fossili potrebbero essere state alterate durante o dopo la fossilizzazione.[108] Ciononostante, risultati simili sono stati scoperti nel teropode sudamericano più antico, il Giganotosaurus . [109] Infatti, anche i dinosauri ornitischi mostrano prove d'omeotermia, mentre tali prove sono assenti nei fossili di varani scoperti nelle stesse formazioni. [110] Anche se T. rex mostrava prove d'omeotermia, ciò non significa necessariamente che l'animale fosse endotermico. La sua termoregolazione potrebbe essere spiegata anche dalla gigantotermia , come dimostrato dalle tartarughe marine. [111] [112]
Nel luglio 2019, uno studio pubblicato sulla rivista The Anatomical Record dai ricercatori delle università di Ohio, Florida e Missouri ha ipotizzato che la funzione delle fosse fronto-parietali presenti nella parte alta della scatola cranica del T. rex , considerati finora i punti di ancoraggio dei muscoli della mascella, sarebbe stata in realtà quella di favorire gli scambi di calore tra i vasi sanguigni e l'ambiente esterno, scaldando o raffreddando la testa dell'animale in base alla temperatura circostante proprio come accade negli odierni coccodrilli. [113]
Locomozione
Ci sono due questioni principali riguardo alle abilità locomotorie di T. rex : la sua abilità di girarsi e quale fosse la sua velocità massima in una corsa dritta. Entrambi sono argomenti importanti nel dibattito che si pone come questione se l'animale fosse un predatore attivo o un necrofago.

Può darsi che T. rex fosse lento nel girarsi, impiegando probabilmente 1-2 secondi per girare solo di 45°, un grado che un umano può girare in una frazione di secondo. [115] La causa della sua difficoltà a girarsi velocemente è collegata al momento di inerzia , siccome la maggior parte della massa di T. rex era lontano dal suo centro di gravità, come un umano che porta un tronco. Avrebbe potuto però parzialmente rimediare inarcando la schiena e la coda, e piegando il collo e le braccia presso il corpo, come fanno i pattinatori per girare più velocemente. [116]

Gli scienziati hanno prodotto un certo numero di stime sulla sua velocità massima; la maggior parte concordano a circa 11 metri al secondo, ma alcuni ipotizzano una velocità ancora più elevata di 20 metri al secondo. Queste ambiguità sono dovute al fatto che, malgrado la presenza di impronte di teropodi che camminano, non si sono ancora trovate impronte di teropodi giganti che corrono. [117] Gli scienziati che ritengono che T. rex fosse un abile corridore puntano al fatto che T. rex avesse le ossa cave (alleggerendo così l'animale) e che altri animali come gli struzzi ei cavalli possono correre velocemente con passi lenti ma lunghi. Inoltre, certi esperti ritengono che T. rex avesse proporzionalmente i muscoli delle gambe più grandi di qualsiasi altro animale moderno, permettendogli così di correre a 40-70 chilometri all'ora. [118]
Nel 1993, Jack Horner e Don Lessem dichiararono che T. rex fosse lento, e che probabilmente fosse incapace di correre, siccome le proporzioni del femore e la tibia sono simili a quelli negli elefanti .[65] Nel 1998 però, Holtz notò che gli arti posteriori dei tirannosauridi avevano componenti prossimali e distali (lo stinco, i piedi e le dita) relativamente più lunghe in proporzione al femore degli altri teropodi, e che i loro metatarsi erano molto più compatti, conducendo così alla conclusione che i tirannosauridi fossero i teropodi giganti più veloci. [119] Thomas Holtz Jr. appoggiò questa ipotesi, affermando che i tirannosauridi avevano piedi proporzionalmente più lunghi di quelli degli allosauridi giganti, un tratto che indica velocità superiore.[92]
Uno studio condotto nel 2003 da Eric Snively e Anthony P. Russel condusse alla scoperta che il terzo metatarso dei tirannosauridi ei suoi legamenti elastici avrebbero funzionato come un 'modello tensile a chiave di volta ' per rinforzare i piedi durante la corsa, conferendo così all'animale una stabilità e una velocità potenziale superiore a quella degli altri teropodi. [114]
Uno studio del 1998 stimò che le ossa delle gambe di T. rex non avevano la robustezza di quelli degli elefanti, che sono animali relativamente lenti e incapaci di correre. Fu proposto in base a questa scoperta che la velocità massima di T. rex sarebbe stata 11 metri al secondo, simile a quella d'uno scattista, ma tali stime dipendono da tante supposizioni dubbiose. [120]
Nel 1995, Farlow ei suoi colleghi sostenerono che un esemplare di T. rex pesante circa 6-8 tonnellate sarebbe rimasto gravemente ferito, o persino ucciso, se fosse caduto durante una corsa, siccome il suo torace avrebbe colpito il suolo a una velocità di 60 metri/s, e gli arti anteriori minuscoli non avrebbero potuto attutire l'impatto. [22] Animali come le giraffe però riescono a mettersi al galoppo a 50 chilometri all'ora, malgrado il rischio di cadere, [121] [122] perciò è possibile che T. rex si spostasse velocemente se necessario. [123] [124]
Gregory S. Paul, scrivendo nel 2000 per la rivista Gaia , propose che le ginocchia flesse ei piedi digitigradi d'un T. rex adulto fossero meglio adatti per la corsa di quelli degli elefanti e gli umani, notando che T. rex aveva un ilio enorme e una cresta cnemiale che avrebbe sostenuto muscoli robusti necessari per la corsa. Inoltre, propose che, malgrado i rischi di ferimento durante la corsa a causa della densità delle ossa, tale adattamento valeva la pena per resistere alle ferite subite durante i combattimenti. [125]
Le ricerche più recenti non appoggiano velocità superiori 40 chilometri orari. Modelli matematici sulla massa muscolare necessaria per superare i 40 chilometri orari dimostrarono che tali velocità fossero irrealizzabili in T. rex , siccome i muscoli delle gambe avrebbero dovuto costituire il 40-86% della massa corporea dell'animale. [117] [118] Un altro studio svolto utilizzando i modelli a computer, basandosi su dati presi direttamente dai fossili, stimò che la velocità massima di T. rex fosse di 8 metri al secondo, quindi leggermente più veloce d'un giocatore di calcio, ma più lento d'uno scattista. [126] [127] [128]
Nel 2010, fu proposto che la velocità di T. rex sarebbe stata aumentata grazie a muscoli caudali (della coda) robusti. [129] Fu infatti notato che le code dei teropodi avessero una distribuzione muscolare diverso da quella degli uccelli moderni ei mammiferi, ma simile a quelli di rettili moderni. [130] Fu concluso che i muscoli caudofemorali avrebbero collegato le ossa della coda con quelle del femore, così assistendo alla retrazione della gamba durante la corsa. [129] Fu anche notato che gli scheletri dei teropodi in generale avessero degli adattamenti (come processi trasversi elevate nella coda) che permettevano la crescita di muscoli caudali grandi, e che la massa muscolare della coda di T. rex fosse stata sottostimata da circa 25-45%. Fu infatti scoperto che il muscolo caudofemorale occupava il 58% della massa muscolare della coda di T. rex . Questo avrebbe migliorato l'equilibrio e l'agilità del predatore. [129] [130]
Gli studiosi che ritengono che T. rex non fosse capace di correre propongono che la sua velocità massima fosse 30 chilometri orari. Questo però è pur sempre superiore alla velocità massima delle sue prede come gli adrosauridi ei ceratopsidi . [118] Inoltre, i proponenti dell'ipotesi che T. rex fosse un predatore attivo notano che l'abilità di correre non fosse importante nell'animale, siccome le sue prede erano comunque più lente. [131] Thomas Holtz infatti notò che il piede di T. rex era relativamente più lungo di quelli degli animali che cacciava.[92] Certi esperti però hanno notato che i ceratopsidi più avanzati avevano le arti anteriori verticali, e che fossero almeno veloci come i rinoceronti . [132] Di conseguenza è plausibile che T. rex fosse solo normalmente lento, ma che in caso di necessità, ad esempio durante la caccia di animali particolarmente veloci come gli adrosauri , potesse rivelarsi un predatore notevolmente veloce e scattante.
Cervello e abilità sensorie

Uno studio condotto da scienziati dell'Ohio University rivelò che T. rex possedeva le abilità sensorie avanzate degli altri coelurosauri , come ad esempio un olfatto elevato, un coordinamento testa/occhi rapido, e persino un'abilità di percezione di suoni a bassa frequenza che gli avrebbe permesso di seguire gli spostamenti delle prede da lunghe distanze. [133] Un ulteriore studio nell'Università di Oregon concluse che l'animale avesse una vista acuta, liquidando così in maniera definitiva l'errata credenza popolare che la sua vista fosse basata solo sul movimento (ossia che vedesse solo le cose che si muovono). Attraverso un'applicazione di perimetria modificata sulle ricostruzioni facciali di vari dinosauri, incluso T. rex , lo studio rivelò che aveva un tasso binoculare di 55 gradi, sorpassando quello degli astori , e superando quello umano ben 13 volte, così per estensione superando persino quello delle aquile , che hanno una vista 3,6 volte più acuta di quella umana. Ciò avrebbe permesso al T. rex di discernere gli oggetti lontani sei chilometri, una capacità superiore ai 1,6 chilometri visibili a un umano. [34] [35] [134] [135]
Thomas Holtz Jr. ipotizzò che la vista acuta di T. rex fosse un adattamento alle prede che cacciava, tra di esse i ceratopsidi (forniti di corna formidabili), dinosauri corazzati come Ankylosaurus , e gli adrosauri , che potrebbero aver avuto comportamenti sociali complessi. Tali caratteristiche nelle prede avrebbe reso la necessità di infliggere ferite precise senza rischio di danneggiarsi, un dettaglio più cruciale che nei teropodi allosauridi , che avevano una percezione di profondità inferiore, e che cacciavano soprattutto i sauropodi , prede più grosse ma molto meno intelligenti.[92]

T. rex è notevolmente evoluto per la grandezza dei suoi bulbi olfattivi ei suoi nervi olfattivi , entrambi responsabili di un olfatto acuto. Ciò indica che T. rex potesse fiutare le carcasse da molto lontano. Il suo olfatto infatti potrebbe corrispondere con quello degli avvoltoi (se non essere superiore). [136] . In tal senso, una ricerca condotta nel 2019 dal biogolo Graham M. Hughes e dal paleontologo John A. Finarelli dell'università di Dublino la paragona a quella dei moderni gatti domestici [137] . Secondo gli studiosi il rettile aveva tra i 620 ei 645 geni dedicati alla codificazione dei recettori olfattivi: quantità leggermente inferiore a quella presente oggi nei polli e nei gatti domestici [138] . Tra i teropodi, T. rex era fornito d'una coclea relativamente lunga, indicando quindi che l'udito era un senso importante per i tirannosauridi. [133]
Uno studio condotto sul quoziente di encefalizzazione (QE) dei rettili, degli uccelli ei teropodi preistorici concluse che T. rex avesse il cervello proporzionalmente più grande di qualsiasi altro dinosauro non-aviano, tranne certi piccoli maniraptoriformi come Bambiraptor , Troodon e Ornithomimus . Lo studio però dimostrò anche che la taglia del cervello di T. rex fosse entro quella dei rettili moderni, in particolare gli alligatori. [139] La taglia del cervello però non basta a determinare l'intelligenza di un animale. A dimostrazione di ciò, gli studi condotti dagli scienziati della Duke University, capitanati da Erich Jarvis, hanno analizzato 6 aree del cervello e constatato che questo dinosauro aveva evoluto comportamenti complessi quali l'elaborazione di informazioni visive e di apprendimento e la capacità di comunicare, emettendo suoni e vocalizzazioni. [140] T. rex risulta essere quindi il teropode gigante più intelligente, capace non solo di attuare strategie di caccia e combattimento nonché operazioni di rudimentale risoluzione di problemi e possibile uso di oggetti come strumenti, ma anche di comunicare coi suoi simili e probabilmente anche di vivere e cacciare in branco (anche se questa teoria è ancora fortemente dibattuta), avendo le capacità intellettive e sensoriali più avanzate di quelle di molti teropodi simili come Giganotosaurus e Spinosaurus .
Alimentazione
Degli studi condotti nel 2012 indicano che la forza del morso di T. rex potrebbe essere stata la più forte di qualsiasi altro animale terrestre mai vissuto. I calcoli suggerirono che la bocca di un T. rex adulto potesse generare una forza di 35.000-57.000 newton nei denti posteriori. Tale pressione è tre volte la forza generata dallo squalo bianco , 15 volte quella del leone , 3½ quella del coccodrillo marino , 77 quella d'un umano, e circa sette volte quella stimata per Allosaurus . [141] [142] [143] Nel 2003 però, stime più alte furono indicate in uno studio dell'Università di Tampa, con una stima di circa 183000-235000 newton (18.3-23.5 tonnellate metriche), una forza equivalente a quella generata dagli esemplari più grandi di Megalodon . [11] [12]


Il dibattito sulla questione se T. rex fosse un predatore attivo o un puro necrofago è vecchio quanto quello sulla sua locomozione. Nel 1917, Lawrence Lambe descrisse un esemplare dell'imparentato Gorgosaurus , concludendo che esso (e T. rex per estensione) fosse un necrofago sulla base dei suoi denti, che non mostravano alcun segno d'usura. [144] Questa posizione però non è più considerata valida, visto che i teropodi rimpiazzavano rapidamente i denti danneggiati. Sin dalla sua scoperta, la maggior parte degli scienziati hanno concluso che T. rex fosse un predatore, ma che si cibava di carogne se l'opportunità si presentava. [145]

Il paleontologo Jack Horner è stato uno dei sostenitori dell'idea che T. rex fosse esclusivamente un necrofago[65] [66] [67] e, pur non avendo mai pubblicato questa ipotesi nella letteratura scientifica, la utilizza per educare il pubblico sui pericoli nel fare supposizioni senza prove. [146] In ogni caso, Horner presentò vari fattori nella letteratura popolare a sostegno dell'ipotesi che l'animale fosse necrofago:
- Siccome T. rex aveva le braccia corte in confronto a quelle degli altri predatori, Horner ha sostenuto che erano incapaci d'immobilizzare le prede. [147]
- T. rex aveva i bulbi olfattivi e nervi olfattivi relativamente grandi, indicando quindi un olfatto ben sviluppato, ideale per rintracciare le carogne, come fanno oggi gli avvoltoi. [136] Gli scienziati che si oppongono all'ipotesi della necrofagia in T. rex notano che gli avvoltoi possono permettersi di essere puramente necrofagi, siccome l'abilità di volare gli dà un vantaggio di vista sopra T. rex , e la loro capacità di planare nell'aria gli permette di consumare pochissima energia nella ricerca di cibo. [148] Ricerche condotte in Glasgow però indicano che un ecosistema produttivo come il Serengeti potrebbe fornire un numero di carcasse sufficienti per sostenere un grande teropode, benché l'animale in questione avrebbe dovuto essere ectotermico per compensare il numero di calorie bruciate nella ricerca di cibo. [149]
- I denti di T. rex potevano frantumare le ossa, permettendogli così di estrarre il midollo . Dei coproliti attribuibili a T. rex dimostrano la presenza di ossa digerite, ma gli esperti notano che i suoi denti non erano adattati a consumare le ossa sistematicamente come le iene . [150]
- Siccome alcune delle prede potenziali di T. rex erano creature agili, le presunte prove che T. rex non potesse correre potrebbero indicare che fosse un necrofago. [66] [151] D'altra parte, delle analisi recenti suggeriscono che T. rex , malgrado fosse stato più lento dei predatori moderni, poteva essere stato sufficientemente veloce per cacciare gli adrosauri ei ceratopsidi . [118] [131]
Ci sono prove che suggeriscono un comportamento da predatore da parte di T. rex . Le sue orbite sono posizionate in tal modo che gli occhi puntavano in avanti, fornendo così all'animale una visione binoculare leggermente superiore a quella d'un falco. Horner stesso notò che la stirpe dei tirannosauri era caratterizzata da un progressivo sviluppo della visione binoculare. Non sarebbe altrimenti chiaro perché la selezione naturale avrebbe favorito lo sviluppo della visione binoculare nei tirannosauri se fossero stati necrofagi dedicati. [34] [35] Negli animali moderni infatti, la visione binoculare si trova soprattutto nei predatori.

Uno scheletro di Edmontosaurus annectens trovato in Montana mostra delle ferite guarite sulla coda, che potrebbero essere dovute a un morso di T. rex . Siccome la ferita guarì, ciò indica che l'animale fu attaccato da vivo. [152] Ci sono anche prove d'uno scontro tra un T. rex e un Triceratops ; un esemplare di quest'ultimo mostra segni d'un morso su un corno e sul "collare" osseo. Il corno ferito infatti mostra segni di guarigione, quindi è probabile che il Triceratops sia sopravvissuto allo scontro. [153]
Nella sua esaminazione dell'esemplare SUE, il paleontologo Peter Larson trovò segni di rottura nella fibula e nelle vertebre caudali, cicatrici sulle ossa facciali, e un dente d'un altro T. rex conficcato in una delle vertebre del collo. Ciò potrebbe costituire una prova di aggressione tra i tirannosauri, ma non è chiaro se si trattasse d'un combattimento territoriale o un tentato cannibalismo . [154] Ulteriori studi però rivelarono che a volte le "ferite" si trattavano d'infezioni o casi di rottura dopo la morte. [66]
Certi esperti propongono che se T. rex fosse veramente un necrofago obbligato, allora un altro dinosauro avrebbe dovuto svolgere il ruolo di predatore alfa nell'ecosistema. Le prede principali includevano i marginocefali più grandi e gli ornitopodi . Siccome gli altri tirannosauri contemporanei condividevano tante caratteristiche con T. rex , gli unici candidati rimasti per il ruolo di predatori alfa erano i piccoli dromaeosauri ei troodontidi . In questo caso, i proponenti dell'ipotesi necrofaga hanno suggerito che T. rex usasse la sua taglia superiore per rubare le prede dei predatori più piccoli, [151] ma avrebbe avuto difficoltà nel trovare carcasse sufficienti, siccome era superato in numero da teropodi più piccoli. [155] La maggior parte dei paleontologi accettano che, come la maggior parte dei predatori, si cibasse sia di carogne che di prede vive.
Come gli altri teropodi, T. rex si nutriva delle carcasse in modo simile ai coccodrilli, afferrando una parte della carcassa e scuotendo la testa lateralmente per strapparla. La testa era meno manovrabile di quella degli allosauroidi, siccome le articolazioni delle vertebre cervicali erano piatte. [156]
Cannibalismo
Nel 2010, i paleontologi Currie, Horner, Erickson e Longrich produssero uno studio che mostrava prove di cannibalismo probabile in T. rex . Studiarono esemplari di T. rex con segni di denti sulle ossa attribuibili a altri tirannosauri. Tali segni furono identificati sull'omero ei metatarsali. I ricercatori proposero che queste ferite indicavano un caso di necrofagia invece d'un combattimento, siccome sarebbe stato difficile mordere i piedi durante uno scontro. Visto che in vita i piedi non portavano tanta carne, è probabile che gli esemplari fossero già stati quasi totalmente denudati di carne quando furono consumati. [157]
Comportamenti sociali

Philip J. Currie dell'Università dell'Alberta ha proposto che T. rex e gli altri tirannosauri potessero essere stati cacciatori sociali, [158] usando come prova un sito nel Dakota del Sud che conteneva tre esemplari di T. rex fossilizzati insieme. [159] Dopo aver studiato il neurocranio di T. rex , Currie propose che l'animale sarebbe stato capace di comportamenti sociali complessi, siccome il cervello era proporzionalmente più grande di quelli dei coccodrilli e tre volte più grande di quelli dei dinosauri erbivori come Triceratops . Currie ipotizzò che T. rex fosse sei volte più intelligente della maggior parte dei dinosauri e gli altri rettili. [160] [161] Currie dichiarò che cacciare in gruppo sarebbe stato vantaggioso per T. rex , siccome le sue prede, come Triceratops e Ankylosaurus , erano ben armate, e alcune erano molto veloci. Ipotizzò che gli esemplari giovanili e adulti di T. rex avrebbero cacciato insieme, con i giovani più veloci che braccavano le prede per poi permettere agli adulti di ucciderle, come fanno i predatori sociali moderni. [160]
L'ipotesi di Currie però è stata criticata d'altri scienziati. Brian Switek, scrivendo nel 2011, criticò l'ipotesi perché non fu presentata in una rivista scientifica, ma in un documentario intitolato Dino Gangs . In più, Switek notò che l'ipotesi di Currie si basava soprattutto sulla presunta sociabilità dell'imparentato Tarbosaurus bataar , le cui prove di sociabilità sono anch'esse dubbiose. Secondo Switek e altri scienziati, che parteciparono in una discussione riguardo Dino Gangs , le prove per la sociabilità nei tirannosauridi sono deboli, e si basano principalmente nell'associazione di vari scheletri scoperti insieme, per cui ci sono spiegazioni alternative (per esempio, una carestia o un'inondazione che costrinse vari individui a morire nello stesso luogo). Infatti, Switek notò che il "cimitero" degli Albertosaurus , da cui Currie sviluppò la sua ipotesi, conserva infatti prove geologiche di un'inondazione, dimostrando quindi che Currie si era sbagliato. Switek dichiarò che le ossa da sole non sono sufficienti per ricostruire i comportamenti dei dinosauri, e che Currie avrebbe fatto meglio a prendere nota delle prove geologiche dei siti contenenti tirannosauri prima di fare conclusioni affrettate sui loro comportamenti. Descrisse le affermazioni fatte nel programma e replicate nella stampa popolare come "promozioni pubblicitarie nauseanti", e notò che la compagnia creatrice del programma, Atlantic Productions, ha la reputazione di esagerare riguardo alle affermazioni sulle scoperte paleontologiche, parallelamente a quella controversa riguardo al presunto antenato umano Darwinius , che poi si rivelò essere invece un lemure . [162]
Secondo Lawrence Witmer, il comportamento sociale non può essere rivelato dai neuro crani, siccome anche i cervelli dei leopardi, che sono felini solitari, sono identici a quelli dei leoni, che sono invece felini sociali, quindi due animali possono avere la stessa intelligenza ma non necessariamente lo stesso stile di vita. Dichiarò che il miglior modo di determinarlo sarebbe stato quello di guardare la grandezza cerebrale totale, metterla a confronto con gli animali moderni e fare stime. Nella sua opinione, è probabile che, se i T.rex cacciavano in gruppo, avrebbero attaccato le prede per beneficio individuale piuttosto per il bene del gruppo. Questa strategia rappresenta uno stadio intermediario tra la caccia solitaria e la caccia cooperativa vera. [163]
Nel 2014, furono trovate le orme d'un gruppo di T. rex in Canada, fornendo così le prime prove di probabile caccia sociale del predatore. [164] [165]
Un'altra curiosità sul comportamento sociale di questi animali riguarda una scoperta di alcuni pseudo nidi fossilizzati, creati da alcune specie di teropodi, tra cui il T. rex, in un probabile rituale di accoppiamento, molto simile a quelli effettuati da alcune specie di uccelli moderni. Questi segni sono stati trovati in una vasta area del Colorado occidentale, e indicano che questi rituali di accoppiamento erano abbastanza diffusi tra i dinosauri teropodi della regione dunque è possibile che potesse esistere una convivenza collettiva tra gli esemplari. [166]
Patologie

L'analisi delle mandibole del cranio di una decina di esemplari, mostranti lesioni vacuolari, che arrivano talora a perforare completamente l'osso, ha permesso di ipotizzare che questi individui siano stati colpiti da una forma di malattia aviaria del tipo Trichomiasi , causata da infezione di parassiti; in precedenza queste lesioni erano state attribuite a segni di morsi da parte di altri predatori o, nel caso dell'esemplare noto come Sue (FMNH PR2081) a infezione da parte di Actinomyces bovis . Le lesioni osservate sono morfologicamente simili a quelle osservabili nei Falconiformi attuali.
Lo studio, [168] presentato nel 2009, rappresenta la prima evidenza sull'origine ed evoluzione di questa malattia contagiosa aviara in dinosauri terapodi non aviari. L'infezione, che dall'analisi dei reperti sembra statisticamente diffusa (circa il 15% degli esemplari), dovrebbe essere imputabile a un protozoo simile all'attuale Trichomonas gallinae , e secondo gli studiosi si sarebbe potuta sviluppare a seguito del consumo di prede infette (per quanto gli stessi studiosi osservino che infezioni di tipo Trichomiasi non siano segnalate in Ornithischia del Cretaceo superiore, suggerendo quindi anche l'ipotesi di contagio per cannibalismo [169] o trasmissione di infezione per morsi durante combattimenti per territorialità o dominanza nel branco o accoppiamento). Le patologie degli esemplari catalogati come FMNH PR2081 e MOR 980 suggeriscono che questa possa essere stata la causa diretta della morte dell'esemplare, impedendone il nutrimento e causandone quindi il decesso.
Distribuzione geografica
Fossili di T. rex sono stati ritrovati in America settentrionale ( Saskatchewan , Texas , Wyoming , Alberta , Montana , Colorado , Dakota del Nord , Dakota del Sud e Nuovo Messico ) [85] e, se il genere Tarbosaurus venisse ridotto a sinonimo di Tyrannosaurus, anche in Asia ( Mongolia ).
Principali ritrovamenti fossili
Più di 30 esemplari di Tyrannosaurus rex sono stati ritrovati ed identificati, alcuni dei quali possono considerarsi scheletri quasi completi. Inoltre, almeno in uno di questi esemplari, sono stati ritrovati tessuti molli e proteine fossili.
Tra gli esemplari più noti e meglio conservati troviamo:
- " Sue ", chiamata così in onore della paleontologa dilettante Sue Hendrickson che la scoprì il 12 agosto 1990 nel Dakota del Sud . Completo all'85% e, fino al 2001, il più grande mai ritrovato, ha richiesto circa 250.000 ore-uomo per riportarlo alla luce. [170] Da uno studio sulla fossilizzazione delle ossa è emerso che "Sue" aveva raggiunto le piene dimensioni dello scheletro a 19 anni ed era morta a 28, l'età più avanzata di qualsiasi tirannosauro noto. [171]
- Un altro Tyrannosaurus, " Stan ", in onore del paleontologo dilettante Stan Sacrison, fu trovato nella provincia di Buffalo nel Sud Dakota, nella primavera del 1987. Ci sono volute 30.000 ore-uomo di scavo e di preparazione, per ricostruire uno scheletro completo al 65%. Il 6 ottobre 2020 è stato venduto all'asta da Christie's per il valore record di 31,8 milioni di dollari, anonimo l'acquirente.
- Nell'estate del 2000, Jack Horner ha scoperto cinque scheletri di Tyrannosaurus vicino alla riserva di Fort Peck nel Montana. Uno degli esemplari, soprannominato " C. Rex " è forse il più grande mai trovato. [172]
Nella cultura di massa
Il tirannosauro è nell'immaginario popolare il carnivoro grande, feroce e inarrestabile per eccellenza ed è forse addirittura il dinosauro più famoso di tutti i tempi, tanto che sin dalla sua scoperta fino ad oggi è definito da molti il Re dei Dinosauri . È inoltre il solo dinosauro di cui il pubblico conosca il nome scientifico completo nonché il sinonimo per eccellenza di dinosauro.
È un personaggio ricorrente (il più delle volte come antagonista) della saga de Alla ricerca della valle incantata , dove viene nominato con l'appellativo "Denti aguzzi". Uno di questi ha ferito mortalmente la madre di Piedino , il cucciolo di apatosauro e protagonista assoluto della serie.
Notevole importanza della sua popolarità è dovuta alle sue apparizioni nella saga cinematografica di Jurassic Park , franchise del quale è diventato un simbolo (malgrado, a partire dal terzo film , il suo ruolo sia stato molto ridimensionato). Nella saga compaiono 4 esemplari di Tyrannosaurus rex: il primo, unico a vivere su Isla Nublar (i fan lo hanno battezzato Rexy), appare in ben 3 film ( Jurassic Park , Jurassic World e Jurassic World - Il regno distrutto ); nel film Il mondo perduto - Jurassic Park appaiono 3 esemplari, dei quali 2 adulti (battezzati Buck e Doe) e un cucciolo (Junior); infine in Jurassic Park III appare un altro esemplare di Tyrannosarus rex adulto (rinominato Bull), il quale però, dopo un combattimento, viene ucciso da uno Spinosaurus .
Appare nel primo episodio della miniserie tv inglese Prehistoric Park , in cui lo zoologo inglese Nigel Marven riesce a riportare al presente due cuccioli (chiamati Terrence e Mathilda).
Il tirannosauro appare nel film d'animazione Disney del 1940 Fantasia , nel 4º episodio La sagra della primavera . Qui lo si vede intento a uccidere uno Stegosauro (in realtà i due dinosauri appartenevano a due periodi diversi: lo stegosauro visse nel Giurassico mentre il tirannosauro comparve nel Cretaceo ). Inoltre nel film è ritratto con le zampe anteriori dotate di tre dita.
Inoltre è la forma animale scelta dal nuovo Megatron , discendente del Megatron originale e capo dei Predacon nella serie Beast Wars .
Appare nel primo episodio della serie animata C'era una volta... l'uomo .
Uno scheletro del dinosauro appare nel film Una notte al museo e nei sequel ( Una notte al museo 2 - La fuga e Una notte al museo 3 - : Il segreto del faraone )
Note
- ^ ( EN ) JF Hicks, KR Johnson, JD Obradovich, L. Tauxe e D. Clark, Magnetostratigraphy and geochronology of the Hell Creek and basal Fort Union Formations of southwestern North Dakota and a recalibration of the Cretaceous–Tertiary Boundary ( PDF ), in Geological Society of America Special Papers , vol. 361, 2002, pp. 35-55, DOI : 10.1130/0-8137-2361-2.35 .
- ^ a b c d e JR Hutchinson, KT Bates, J. Molnar, V. Allen e PJ Makovicky, A Computational Analysis of Limb and Body Dimensions in Tyrannosaurus rex with Implications for Locomotion, Ontogeny, and Growth , in PLoS ONE , vol. 6, n. 10, 2011, p. e26037, DOI : 10.1371/journal.pone.0026037 .
- ^ a b c Sue's vital statistics , su Sue at the Field Museum , Field Museum of Natural History . URL consultato il 15 settembre 2007 (archiviato dall' url originale il 29 settembre 2007) .
- ^ a b c Gregory M. Erickson, Peter J. Makovicky, Philip J. Currie , Mark A. Norell, Scott A. Yerby e Christopher A. Brochu, Gigantism and comparative life-history parameters of tyrannosaurid dinosaurs , in Nature , vol. 430, n. 7001, 2004, pp. 772-775, DOI : 10.1038/nature02699 , PMID 15306807 .
- ^ a b c d Scotty, il T.rex più grande mai scoperto , su nationalgeographic.it . URL consultato il 12 aprile 2019 (archiviato dall' url originale il 1º aprile 2019) .
- ^ a b c d e Scotty, un T.rex da record: è il più grande mai scoperto , su repubblica.it .
- ^ Scotty, un T.rex da record: è il più grande mai scoperto. , su repubblica.it .
- ^ Horner John R - Padian Kvin 2004, Age an growth dynamics of Tyrannosaurus rex , in Royal Society Publishing , vol. 271, 1551/22 settembre 2004 Pagg. 1875-1880..
- ^ ( EN ) Brian Switeck, When Tyrannosaurus Chomped Sauropods , su blogs.smithsonianmag.com , Smithsonian Media, 13 aprile 2012. URL consultato il 24 agosto 2013 .
- ^ ( EN ) John Hutchinson, Tyrannosaurus rex: predator or media hype? , su whatsinjohnsfreezer.com , What's in John's Freezer?, 15 luglio 2013. URL consultato il 26 agosto 2013 .
- ^ a b c MB Meers, Maximum bite force and prey size of Tyrannosaurus rex and their relationships to the inference of feeding behavior , in Historical Biology: A Journal of Paleobiology , vol. 16, n. 1, agosto 2003, pp. 1-12, DOI : 10.1080/0891296021000050755 .
- ^ a b Copia archiviata , su utweb.ut.edu . URL consultato il 14 marzo 2015 (archiviato dall' url originale il 1º febbraio 2014) . , Meers, Mason B. (2003) T. Rex Bite Force and Prey Size. Retrieved on July 15, 2013 from Copia archiviata , su utweb.ut.edu . URL consultato il 14 marzo 2015 (archiviato dall' url originale il 1º febbraio 2014) .
- ^ Andrea Centini, Il morso del T-Rex era devastante : poteva davvero distruggere un auto,come in jurassic park , su scienze.fanpage.it .
- ^ Andrea Centini, Il morso del T-Rex era devastante : poteva davvero distruggere un auto, come in jurassic park , su scienze.fanpage.it .
- ^ Andrea Centini, Il morso del T-Rex era devastante : poteva davvero distruggere un auto, come in jurassic park , su scienze.fanpage.it .
- ^ a b Andrea Centini, Il morso del T-rex era devastante: poteva davvero distruggere un'auto, come in Jurassic Park , su scienze.fanpage.it , fanpage.it , 1º ottobre 2019. URL consultato il 13 ottobre 2019 .
- ^ Andrea Cau, Theropoda volume I: Tyrannosauroidea , Amazon Media EU S.à rl, 2012.
- ^ Il Pianeta dei dinosauri , Piero e Alberto Angela, Arnoldo Monadori Editore , Ottobre 1993, pag .55 .
- ^ JF Anderson, AJ Hall-Martin e Dale Russell ,Long bone circumference and weight in mammals, birds and dinosaurs , in Journal of Zoology , vol. 207, n. 1, 1985, pp. 53-61, DOI : 10.1111/j.1469-7998.1985.tb04915.x .
- ^ a b Robert T. Bakker , The Dinosaur Heresies , New York, Kensington Publishing, 1986, ISBN 0-688-04287-2 , OCLC 13699558 .
- ^ DM Henderson, Estimating the masses and centers of mass of extinct animals by 3-D mathematical slicing , in Paleobiology , vol. 25, n. 1, 1º gennaio 1999, pp. 88-106.
- ^ a b JO Farlow, MB Smith e JM Robinson, Body mass, bone "strength indicator", and cursorial potential of Tyrannosaurus rex , in Journal of Vertebrate Paleontology , vol. 15, n. 4, 1995, pp. 713-725, DOI : 10.1080/02724634.1995.10011257 (archiviato dall' url originale il 23 ottobre 2008) .
- ^ Frank. Seebacher, [0051:ANMTCA 2.0.CO;2 A new method to calculate allometric length–mass relationships of dinosaurs ], in Journal of Vertebrate Paleontology , vol. 21, n. 1, 2001, pp. 51-60, DOI : 10.1671/0272-4634(2001)021[0051:ANMTCA]2.0.CO;2 .
- ^ Per Christiansen e Richard A. Fariña, Mass prediction in theropod dinosaurs , in Historical Biology , vol. 16, 2–4, 2004, pp. 85-92, DOI : 10.1080/08912960412331284313 .
- ^ TJ Boardman, GC Packard e GF Birchard, Allometric equations for predicting body mass of dinosaurs , in Journal of Zoology , vol. 279, n. 1, 2009, pp. 102-110, DOI : 10.1111/j.1469-7998.2009.00594.x .
- ^ Scott Hartman, Mass estimates: North vs South redux , su skeletaldrawing.com , Scott Hartman's Skeletal Drawing.com, 7 luglio 2013. URL consultato il 24 agosto 2013 .
- ^ http://dinoweb.ucoz.ru/_fr/4/My_theropod_is_.pdf
- ^ F. Therrien e Henderson, DM, [108:MTIBTY 2.0.CO;2 My theropod is bigger than yours...or not: estimating body size from skull length in theropods ], in Journal of Vertebrate Paleontology , vol. 27, n. 1, 2007, pp. 108-115, DOI : 10.1671/0272-4634(2007)27[108:MTIBTY]2.0.CO;2 , ISSN 0272-4634 .
- ^ Tyrannosaurus rex - Wikiversità , su it.wikiversity.org . URL consultato l'11 febbraio 2019 .
- ^ Horner John R - Padian Kvin 2004, Age an growth dynamics of Tyrannosaurus rex , in Royal Society Publishing , Volume 271,, Numero 1551/22 settembre 2004 Pagg. 1875-1880.
- ^ a b c d CR Brochu, Osteology of Tyrannosaurus rex : insights from a nearly complete skeleton and high-resolution computed tomographic analysis of the skull , in Society of Vertebrate Paleontology Memoirs , vol. 7, 2003, pp. 1-138, DOI : 10.2307/3889334 .
- ^ Christine Lipkin e Kenneth Carpenter, Looking again at the forelimb of Tyrannosaurus rex , in Carpenter, Kenneth; and Larson, Peter E. (editors) (a cura di), Tyrannosaurus rex, the Tyrant King (Life of the Past) , Bloomington, Indiana University Press, 2008, pp. 167 –190, ISBN 0-253-35087-5 .
- ^ Museum unveils world's largest T-rex skull , su montana.edu . URL consultato il 13 settembre 2008 (archiviato dall' url originale il 14 aprile 2006) .
- ^ a b c Kent A. Stevens, Binocular vision in theropod dinosaurs ( PDF ), in Journal of Vertebrate Paleontology , vol. 26, n. 2, giugno 2006, pp. 321-330, DOI : 10.1671/0272-4634(2006)26[321:BVITD]2.0.CO;2 .
- ^ a b c Eric Jaffe, Sight for 'Saur Eyes: T. rex vision was among nature's best , in Science News , vol. 170, n. 1, 1º luglio 2006, p. 3, DOI : 10.2307/4017288 . URL consultato il 6 ottobre 2008 (archiviato dall' url originale il 19 ottobre 2008) .
- ^ Eric Snively, Donald M. Henderson, and Doug S. Phillips, Fused and vaulted nasals of tyrannosaurid dinosaurs: Implications for cranial strength and feeding mechanics ( PDF ), in Acta Palaeontologica Polonica , vol. 51, n. 3, 2006, pp. 435-454. URL consultato l'8 ottobre 2008 .
- ^ GM Erickson, Van Kirk, SD; Su, J.; Levenston, ME; Caler, WE; and Carter, DR, Bite-force estimation for Tyrannosaurus rex from tooth-marked bones , in Nature , vol. 382, 1996, pp. 706-708, DOI : 10.1038/382706a0 .
- ^ Smith, JB, Heterodonty in Tyrannosaurus rex : implications for the taxonomic and systematic utility of theropod dentitions ( PDF ), in Journal of Vertebrate Paleontology , vol. 25, n. 4, dicembre 2005, pp. 865-887, DOI : 10.1671/0272-4634(2005)025[0865:HITRIF]2.0.CO;2 .
- ^ Douglas K, Young S, The dinosaur detectives , in New Scientist , 1998. URL consultato il 16 ottobre 2008 .
«One palaeontologist memorably described the huge, curved teeth of T. rex as 'lethal bananas'» . - ^ ( EN ) Nicholas St Fleur, Between a T. Rex's Powerful Jaws, Bones of Its Prey Exploded , in The New York Times , 18 maggio 2017. URL consultato il 6 agosto 2019 .
- ^ ( EN ) Riley Black, The Tyrannosaurus Rex's Dangerous and Deadly Bite , su Smithsonian . URL consultato il 6 agosto 2019 .
- ^ ( EN ) Nicholas St Fleur, Between a T. Rex's Powerful Jaws, Bones of Its Prey Exploded , in The New York Times , 18 maggio 2017. URL consultato il 6 agosto 2019 .
- ^ Redazione ANSA, Misurata l'incredibile forza del morso del T-rex , su ANSA , 18 maggio 2017.
- ^ a b Keim, B. (2012). " Giant Feathered Tyrannosaur Found in China. " Wired , April 4, 2012. Accessed online August 8, 2013, https://www.wired.com/wiredscience/2012/04/yutyrannus-huali-feathers/
- ^ Hone, D. (2012) "Did Tyrannosaurus rex have feathers?" The Guardian , October 17, 2012. Accessed online August 8, 2013, https://www.theguardian.com/science/lost-worlds/2012/oct/17/dinosaurs-fossils
- ^ a b Xing Xu, Mark A. Norell, Xuewen Kuang, Xiaolin Wang, Qi Zhao e Chengkai Jia, Basal tyrannosauroids from China and evidence for protofeathers in tyrannosauroids , in Nature , vol. 431, n. 7009, 7 ottobre 2004, pp. 680-684, DOI : 10.1038/nature02855 , PMID 15470426 .
- ^ a b X. Xu, K. Wang, K. Zhang, Q. Ma, L. Xing, C. Sullivan, D. Hu, S. Cheng e S. et al. Wang, A gigantic feathered dinosaur from the Lower Cretaceous of China ( PDF ), in Nature , vol. 484, 2012, pp. 92-95, DOI : 10.1038/nature10906 , PMID 22481363 (archiviato dall' url originale il 17 aprile 2012) .
- ^ a b Tyrannosauroid integument reveals conflicting patterns of gigantism and feather evolution , su royalsocietypublishing.org .
- ^ a b National Geographic Society, Che effetto farebbe accarezzare un T-Rex? , su National Geographic . URL consultato il 10 febbraio 2019 (archiviato dall' url originale il 12 febbraio 2019) .
- ^ a b Il tirannosauro aveva le squame e non le penne: l'errore degli scienziati cambia la storia , su Scienze fanpage . URL consultato il 10 febbraio 2019 .
- ^ a b ( EN ) Brian Switek, Palaeontology: The truth about T. rex , in Nature News , vol. 502, n. 7472, 24 ottobre 2013, p. 424, DOI : 10.1038/502424a . URL consultato l'11 febbraio 2019 .
- ^ https://news.nationalgeographic.com/2017/03/tyrannosaur-face-touch-dinosaurs-evolution-science/
- ^ https://www.thesun.co.uk/news/3214931/tyrannosaurus-rex-fossil-reveals-that-fearsome-beast-looked-like-a-crocodile-and-had-incredible-sixth-sense/
- ^ http://www.upi.com/Science_News/2017/03/30/Scientists-discover-new-dinosaur-evolved-by-anagenesis/7461490880589/
- ^ Kenneth Carpenter , Variation in Tyrannosaurus rex , in Kenneth Carpenter and Philip J. Currie (a cura di), Dinosaur Systematics: Approaches and Perspectives , Cambridge, Cambridge University Press, 1992, pp. 141–145, ISBN 0-521-43810-1 .
- ^ Larson, PL 1994. Tyrannosaurus sex. In: Rosenberg, GD & Wolberg, DL Dino Fest . The Paleontological Society Special Publications . 7: 139–155.
- ^ Schweitzer MH, Elsey RM, Dacke CG, Horner JR, Lamm ET, Do egg-laying crocodilian (Alligator mississippiensis) archosaurs form medullary bone? , in Bone , vol. 40, n. 4, aprile 2007, pp. 1152-8, DOI : 10.1016/j.bone.2006.10.029 , PMID 17223615 . URL consultato l'8 ottobre 2008 .
- ^ a b c d Gregory S. Paul , Predatory dinosaurs of the world: a complete illustrated guide , New York, Simon and Schuster, 1988, ISBN 0-671-61946-2 , OCLC 18350868 .
- ^ J Leidy, Memoir on the extinct reptiles of the Cretaceous formations of the United States , in Smithsonian Contributions to Knowledge , vol. 14, 1865, pp. 1-135.
- ^ Tyrannosaurus , su amnh.org , American Museum of Natural History . URL consultato il 16 ottobre 2008 (archiviato dall' url originale l'8 dicembre 2008) .
- ^ a b BH Newman,Stance and gait in the flesh-eating Tyrannosaurus , in Biological Journal of the Linnean Society , vol. 2, n. 2, 1970, pp. 119-123, DOI : 10.1111/j.1095-8312.1970.tb01707.x .
- ^ HF Osborn , Tyrannosaurus and other Cretaceous carnivorous dinosaurs , in Bulletin of the AMNH , vol. 21, n. 14, New York City, American Museum of Natural History, 1905, pp. 259-265. URL consultato il 6 ottobre 2008 .
- ^ HF Osborn , Skeletal adaptations of Ornitholestes, Struthiomimus, Tyrannosaurus , in Bulletin of the American Museum of Natural History , vol. 35, n. 43, New York City, American Museum of Natural History, 1917, pp. 733-771. URL consultato l'8 ottobre 2008 .
- ^ LM Lambe , On a new genus and species of carnivorous dinosaur from the Belly River Formation of Alberta, with a description of the skull of Stephanosaurus marginatus from the same horizon , in Ottawa Naturalist , vol. 27, 1914, pp. 129-135.
- ^ a b c John R. Horner , Don Lessem , The complete T. rex , New York City, Simon & Schuster, 1993, ISBN 0-671-74185-3 .
- ^ a b c d Horner, JR, Steak knives, beady eyes, and tiny little arms (a portrait of Tyrannosaurus as a scavenger) , in The Paleontological Society Special Publication , vol. 7, 1994, pp. 157-164.
- ^ a b Amos, J., T. rex goes on trial , su news.bbc.co.uk , BBC, 31 luglio 2003.
- ^ a b Kenneth Carpenter , Matt Smith,Forelimb Osteology and Biomechanics of Tyrannosaurus rex , in Darren Tanke and Kenneth Carpenter (a cura di), Mesozoic vertebrate life , Bloomington, Indiana University Press, 2001, pp. 90–116, ISBN 0-253-33907-3 .
- ^ Henry Fairfield Osborn , Barnum Brown, Tyrannosaurus, Upper Cretaceous carnivorous dinosaur , in Bulletin of the AMNH , vol. 22, n. 16, New York City, American Museum of Natural History, 1906, pp. 281-296. URL consultato il 6 ottobre 2008 .
- ^ a b Philip J. Currie, Jørn H. Hurum e Karol Sabath, Skull structure and evolution in tyrannosaurid dinosaurs ( PDF ), in Acta Palaeontologica Polonica , vol. 48, n. 2, 2003, pp. 227-234. URL consultato l'8 ottobre 2008 .
- ^ a b c Thomas R., Jr. Holtz , Tyrannosauroidea , in David B. Weishampel , Peter Dodson and Halszka Osmólska (a cura di), The dinosauria , Berkeley, University of California Press, 2004, pp. 111 –136, ISBN 0-520-24209-2 .
- ^ a b Thomas R. Holtz , The Phylogenetic Position of the Tyrannosauridae: Implications for Theropod Systematics , in Journal of Palaeontology , vol. 68, n. 5, 1994, pp. 1100-1117, JSTOR 1306180 .
- ^ ( RU ) EA Maleev, (title in Russian) [ Gigantic carnivorous dinosaurs of Mongolia ] ( PDF ), in Doklady Akademii Nauk SSSR , translated by FJ Alcock, vol. 104, n. 4, 1955, pp. 634-637.
- ^ AK Rozhdestvensky, Growth changes in Asian dinosaurs and some problems of their taxonomy , in Paleontological Journal , vol. 3, 1965, pp. 95-109.
- ^ Kenneth Carpenter , Tyrannosaurids (Dinosauria) of Asia and North America , in Niall J. Mateer and Pei-ji Chen (a cura di), Aspects of nonmarine Cretaceous geology , Beijing, China Ocean Press, 1992, ISBN 978-7-5027-1463-5 , OCLC 28260578 .
- ^ Thomas D. Carr, Thomas E. Williamson e David R. Schwimmer, [0119:ANGASO 2.0.CO;2 A New Genus and Species of Tyrannosauroid from the Late Cretaceous (Middle Campanian) Demopolis Formation of Alabama ], in Journal of Vertebrate Paleontology , vol. 25, n. 1, marzo 2005, pp. 119-143, DOI : 10.1671/0272-4634(2005)025[0119:ANGASO]2.0.CO;2 .
- ^ Jørn H. Hurum e Karol Sabath, Giant theropod dinosaurs from Asia and North America: Skulls of Tarbosaurus bataar and Tyrannosaurus rex compared ( PDF ), in Acta Palaeontologica Polonica , vol. 48, n. 2, 2003, pp. 161-190. URL consultato l'8 ottobre 2008 .
- ^ George Olshevsky , The origin and evolution of the tyrannosaurids , in Kyoryugaku Saizensen [Dino Frontline] , 9–10, 1995, pp. 92-119.
- ^ TD Carr e TE Williamson, Diversity of late Maastrichtian Tyrannosauridae (Dinosauria: Theropoda) from western North America , in Zoological Journal of the Linnean Society , vol. 142, n. 4, 2004, pp. 479-523, DOI : 10.1111/j.1096-3642.2004.00130.x .
- ^ CW Gilmore, A new carnivorous dinosaur from the Lance Formation of Montana , in Smithsonian Miscellaneous Collections , vol. 106, 1946, pp. 1-19.
- ^ RT Bakker, M. Williams e PJ Currie, Nanotyrannus , a new genus of pygmy tyrannosaur, from the latest Cretaceous of Montana , in Hunteria , vol. 1, n. 5, 1988, pp. 1-30.
- ^ TD Carr, Craniofacial ontogeny in Tyrannosauridae (Dinosauria, Theropoda) , in Journal of Vertebrate Paleontology , vol. 19, n. 3, 1999, pp. 497-520, DOI : 10.1080/02724634.1999.10011161 .
- ^ Philip J. Currie , Cranial anatomy of tyrannosaurid dinosaurs from the Late Cretaceous of Alberta, Canada ( PDF ), in Acta Palaeontologica Polonica , vol. 42, n. 2, 2003, pp. 191-226. URL consultato il 9 ottobre 2008 .
- ^ DOI : 10.1371/journal.pone.0079420
- ^ a b †Tyrannosaurus rex Osborn 1905 (tyrant lizard king) , su fossilworks.org . URL consultato il 12 marzo 2015 .
- ^ a b http://news.discovery.com/animals/dinosaurs/t-rex-was-likely-an-invasive-species-160302.htm
- ^ Erickson GM, Makovicky PJ, Currie PJ, Norell MA, Yerby SA, Brochu CA, Gigantism and comparative life-history parameters of tyrannosaurid dinosaurs , in Nature , vol. 430, n. 7001, 2004, pp. 772-775, DOI : 10.1038/nature02699 , PMID 15306807 .
- ^ a b JR Horner e K. Padian, Age and growth dynamics of Tyrannosaurus rex , in Proceedings. Biological sciences / the Royal Society , vol. 271, n. 1551, settembre 2004, pp. 1875-80, DOI : 10.1098/rspb.2004.2829 , PMC 1691809 , PMID 15347508 . URL consultato il 5 ottobre 2008 .
- ^ Schweitzer MH, Wittmeyer JL, Horner JR, Gender-specific reproductive tissue in ratites and Tyrannosaurus rex , in Science , vol. 308, n. 5727, giugno 2005, pp. 1456-60, DOI : 10.1126/science.1112158 , PMID 15933198 . URL consultato il 5 ottobre 2008 .
- ^ Andrew H. Lee e Sarah Werning,Sexual maturity in growing dinosaurs does not fit reptilian growth models , in Proceedings of the National Academy of Sciences , vol. 105, n. 2, 2008, pp. 582-587, DOI : 10.1073/pnas.0708903105 , PMC 2206579 , PMID 18195356 .
- ^ a b Erickson GM, Currie PJ, Inouye BD, Winn AA, Tyrannosaur life tables: an example of nonavian dinosaur population biology , in Science , vol. 313, n. 5784, luglio 2006, pp. 213-7, DOI : 10.1126/science.1125721 , PMID 16840697 .
- ^ a b c d Holtz, Thomas R Jr. (19 marzo, 2013) The Life and Times of Tyrannosaurus rex, with Dr. Thomas Holtz (conferenza), Burke Museum of Natural History and Culture , Kane Hall Room 130 University of Washington Seattle, WA 98195
- ^ Gregory S. Paul, Chapter 18: The Extreme Life Style and Habits of the Gigantic Tyrannosaurid Superpredators of the Cretaceous North America and Asia , in Peter L. Larson e Kenneth Carpenter (a cura di), Tyrannosaurus, The Tyrant King , Indiana University Press, 2008, pp. 307-345, ISBN 978-0-253-35087-9 . URL consultato il 14 settembre 2013 .
- ^ ( EN ) Charles R. Marshall, Daniel V. Latorre e Connor J. Wilson, Absolute abundance and preservation rate of Tyrannosaurus rex , in Science , vol. 372, n. 6539, 16 aprile 2021, pp. 284-287, DOI : 10.1126/science.abc8300 . URL consultato il 17 aprile 2021 .
- ^ Paolo Virtuani, T. Rex: erano 2,5 miliardi, a ognuno servivano 110 kmq per vivere , su Corriere della Sera , 17 aprile 2021. URL consultato il 17 aprile 2021 .
- ^ a b Helen Fields, Dinosaur Shocker , in Smithsonian Magazine , maggio 2006. URL consultato il 2 ottobre 2008 .
- ^ Mary H. Schweitzer , Jennifer L. Wittmeyer, John R. Horner e Jan K. Toporski, Soft-tissue vessels and cellular preservation in Tyrannosaurus rex , in Science , vol. 307, n. 5717, marzo 2005, pp. 1952-5, Bibcode : 2005Sci...307.1952S , DOI : 10.1126/science.1108397 , PMID 15790853 . URL consultato il 2 ottobre 2008 .
- ^ Paul Rincon, Protein links T. rex to chickens , in BBC News , 12 aprile 2007. URL consultato il 2 ottobre 2008 .
- ^ Dan Vergano, Yesterday's T. Rex is today's chicken , in USA Today , 13 aprile 2007. URL consultato l'8 ottobre 2008 .
- ^ Randolph E. Schmid e Associated Press , Scientists study evidence modern birds came from dinosaurs , in Newsvine , 24 aprile 2008. URL consultato l'8 ottobre 2008 .
- ^ Thomas G. Kaye, Gary Gaugler e Zbigniew Sawlowicz,Dinosaurian Soft Tissues Interpreted as Bacterial Biofilms , in Anna Stepanova (a cura di), PLoS ONE , vol. 3, n. 7, luglio 2008, pp. e2808, DOI : 10.1371/journal.pone.0002808 , PMC 2483347 , PMID 18665236 .
- ^ New Research Challenges Notion That Dinosaur Soft Tissues Still Survive , su newswise.com , Newswise, 24 luglio 2008. URL consultato l'8 ottobre 2008 .
- ^ Researchers Debate: Is It Preserved Dinosaur Tissue, or Bacterial Slime? , su blogs.discovermagazine.com , Discover, 30 luglio 2008. URL consultato il 4 settembre 2008 .
- ^ a b http://www.corriere.it/cronache/16_marzo_16/scoperta-t-rex-incinta-forse-presente-dna-f5156cac-ebb3-11e5-bc24-916916ab221e.shtml?refresh_ce-cp
- ^ Robert T. Bakker , The superiority of dinosaurs ( PDF ), in Discovery , vol. 3, n. 2, 1968, pp. 11-12. URL consultato il 7 ottobre 2008 (archiviato dall' url originale il 9 settembre 2006) .
- ^ Robert T. Bakker , Anatomical and ecological evidence of endothermy in dinosaurs ( PDF ), in Nature , vol. 238, n. 5359, 1972, pp. 81-85, Bibcode : 1972Natur.238...81B , DOI : 10.1038/238081a0 . URL consultato il 7 ottobre 2008 (archiviato dall' url originale il 9 settembre 2006) .
- ^ Reese E. Barrick e William J. Showers, Thermophysiology of Tyrannosaurus rex: Evidence from Oxygen Isotopes , in Science , vol. 265, n. 5169, New York City, luglio 1994, pp. 222-224, DOI : 10.1126/science.265.5169.222 , PMID 17750663 . URL consultato il 7 ottobre 2008 .
- ^ Clive Trueman, Carolyn Chenery, David A. Eberth e Baruch Spiro, Diagenetic effects on the oxygen isotope composition of bones of dinosaurs and other vertebrates recovered from terrestrial and marine sediments , in Journal of the Geological Society , vol. 160, n. 6, 2003, p. 895, DOI : 10.1144/0016-764903-019 .
- ^ Reese E. Barrick e William J. Showers, Thermophysiology and biology of Giganotosaurus : comparison with Tyrannosaurus , in Palaeontologia Electronica , vol. 2, n. 2, ottobre 1999. URL consultato il 7 ottobre 2008 .
- ^ Reese E. Barrick, Michael K. Stoskopf e William J. Showers, Oxygen isotopes in dinosaur bones , in James O. Farlow and MK Brett-Surman (a cura di), The Complete Dinosaur , Bloomington, Indiana University Press, 1999, pp. 474-490, ISBN 0-253-21313-4 .
- ^ Frank V. Paladino, James R. Spotila e Peter Dodson, A blueprint for giants: modeling the physiology of large dinosaurs , in James O. Farlow and MK Brett-Surman (a cura di), The Complete Dinosaur , Bloomington, Indiana University Press, 1999, pp. 491-504, ISBN 0-253-21313-4 .
- ^ Anusuya Chinsamy e Willem J. Hillenius, Physiology of nonavian dinosaurs , in David B. Weishampel, Peter Dodson and Halszka Osmólska (a cura di), The dinosauria , Berkeley, University of California Press, 2004, pp. 643 –659, ISBN 0-520-24209-2 .
- ^ ( EN ) Casey M. Holliday, William Ruger Porter, Kent A. Vliet, Lawrence M. Witmer, The Frontoparietal Fossa and Dorsotemporal Fenestra of Archosaurs and Their Significance for Interpretations of Vascular and Muscular Anatomy in Dinosaurs , su anatomypubs.onlinelibrary.wiley.com , American Association for Anatomy, 1º luglio 2019. URL consultato il 7 settembre 2019 .
- ^ a b Copia archiviata ( PDF ), su ohio.edu . URL consultato il 17 ottobre 2015 (archiviato dall' url originale il 24 settembre 2015) . , Snively, Eric, Russell, Anthony P. (2003) "Kinematic Model of Tyrannosaurid (Dinosauria: Theropoda) Arctometatarsus Function" Journal of Morphology 255(2)215-227. DOI : 10.1002/jmor.10059
- ^ Hutchinson JR, Ng-Thow-Hing V, Anderson FC, A 3D interactive method for estimating body segmental parameters in animals: application to the turning and running performance of Tyrannosaurus rex , in Journal of Theoretical Biology , vol. 246, n. 4, giugno 2007, pp. 660-80, DOI : 10.1016/j.jtbi.2007.01.023 , PMID 17363001 .
- ^ David R. Carrier, Rebecca M. Walter e David V. Lee, Influence of rotational inertia on turning performance of theropod dinosaurs: clues from humans with increased rotational inertia , in Journal of Experimental Biology , vol. 204, n. 22, Company of Biologists , 15 novembre 2001, pp. 3917-3926, PMID 11807109 .
- ^ a b Hutchinson, JR, Biomechanical Modeling and Sensitivity Analysis of Bipedal Running Ability. II. Extinct Taxa ( PDF ), in Journal of Morphology , vol. 262, n. 1, 2004, pp. 441-461, DOI : 10.1002/jmor.10240 , PMID 15352202 (archiviato dall' url originale il 31 ottobre 2008) .
- ^ a b c d Hutchinson JR, Garcia M, Tyrannosaurus was not a fast runner , in Nature , vol. 415, n. 6875, febbraio 2002, pp. 1018-21, DOI : 10.1038/4151018a , PMID 11875567 .
- ^ Thomas R. Holtz , Phylogenetic taxonomy of the Coelurosauria (Dinosauria; Theropoda) , in Journal of Paleontology , vol. 70, n. 3, 1º maggio 1996, pp. 536-538. URL consultato il 3 ottobre 2008 .
- ^ Christiansen, P., Strength indicator values of theropod long bones, with comments on limb proportions and cursorial potential ( PDF ), in Gaia , vol. 15, 1998, pp. 241-255, ISSN 0871-5424 .
- ^ Giraffe , su wildlifesafari.info . URL consultato il 29 aprile 2006 (archiviato dall' url originale il 12 giugno 2010) .
- ^ Chronological History of Woodland Park Zoo – Chapter 4 , su zoo.org . URL consultato il 24 ottobre 2014 .
- ^ Alexander, RM,Dinosaur biomechanics , in Proc Biol Sci. , vol. 273, n. 1596, The Royal Society, 7 agosto 2006, pp. 1849-1855, DOI : 10.1098/rspb.2006.3532 , PMC 1634776 , PMID 16822743 .
- ^ Rebecca R. Hanna, [0076:MIAIIA 2.0.CO;2 Multiple injury and infection in a sub-adult theropod dinosaur ( Allosaurus fragilis ) with comparisons to allosaur pathology in the Cleveland-Lloyd dinosaur quarry collection ], in Journal of Vertebrate Paleontology , vol. 22, n. 1, 2002, pp. 76-90, DOI : 10.1671/0272-4634(2002)022[0076:MIAIIA]2.0.CO;2 , ISSN 0272-4634 . catalogs the injuries of the Allosaurus known as "Big Al" – at least one was attributed to a fall.
- ^ Gregory S. Paul, Limb design, function and running performance in ostrich-mimics and tyrannosaurs ( PDF ), in Gaia , vol. 15, 2000, pp. 257-270. URL consultato il 2013 .
- ^ Sellers, WI, and Manning, PL, Estimating dinosaur maximum running speeds using evolutionary robotics , in Proc. R. Soc. B , vol. 274, n. 1626, The Royal Society, luglio 2007, pp. 2711-6, DOI : 10.1098/rspb.2007.0846 , PMC 2279215 , PMID 17711833 .
- ^ L Seward, T. rex 'would outrun footballer' , su news.bbc.co.uk , BBCNews, 21 agosto 2007. URL consultato il 16 ottobre 2008 .
- ^ G. Callison e HM Quimby, Tiny dinosaurs: Are they fully grown? , in Journal of Vertebrate Paleontology , vol. 3, n. 4, 1984, pp. 200-209, DOI : 10.1080/02724634.1984.10011975 .
- ^ a b c Scott W. Persons e Philip J. Currie, The Tail of Tyrannosaurus: Reassessing the Size and Locomotive Importance of the M. caudofemoralis in Non-Avian Theropods , in The Anatomical Record: Advances in Integrative Anatomy and Evolutionary Biology , vol. 294, n. 1, gennaio 2011 [Article first published online November 12, 2010] , pp. 119–131,, DOI : 10.1002/ar.21290 . URL consultato il 2013 .
- ^ a b Persons, Scott W. (2010) Guest Post: Bulking-Up the Back End – Why Tyrannosaurus Tail Mass Matters . (2010) Retrieved from https://archosaurmusings.wordpress.com/2010/12/06/guest-post-bulking-up-the-back-end-why-tyrannosaurus-tail-mass-matters/
- ^ a b Manning P, T. rex speed trap , in Carpenter, Kenneth; Larson, Peter E. (a cura di), Tyrannosaurus rex, the Tyrant King (Life of the Past) , Bloomington, Indiana University Press, 2008, pp. 205 –228, ISBN 0-253-35087-5 .
- ^ Paul, GS, and Christiansen, P., Forelimb posture in neoceratopsian dinosaurs: implications for gait and locomotion , in Paleobiology , vol. 26, n. 3, settembre 2000, p. 450, DOI : 10.1666/0094-8373(2000)026<0450:FPINDI>2.0.CO;2 , ISSN 0094-8373 .
- ^ a b [1] , Witmer, Lawrence M., Ridgely, Ryan C. (Aug 26, 2009) New insights into the brain, braincase, and ear region of tyrannosaurs (Dinosauria, Theropoda), with implications for sensory organization and behavior. The Anatomical Record 292(9) 1266-1296. DOI : 10.1002/ar.20983
- ^ [2] [ collegamento interrotto ] , Emily, John(July 3, 2006). Supersight for a Dino King . Retrieved July 7, 2006 from http://www.sciencenewsforkids.org/2006/07/supersight-for-a-dino-king-3/ [ collegamento interrotto ]
- ^ [3] , Stevens, Kent A. (April 1, 2011) The Binocular Vision of Theropod Dinosaurs . Retrieved July 29, 2013 from http://ix.cs.uoregon.edu/~kent/paleontology/binocularVision/
- ^ a b T. Rex brain study reveals a refined 'nose' , su canada.com , Calgary Herald, 28 ottobre 2008. URL consultato il 29 ottobre 2008 .
- ^ Hughes Graham M. e Finarelli John A., Olfactory receptor repertoire size in dinosaurs , in Proceedings of the Royal Society B: Biological Sciences , vol. 286, n. 1904, 12 giugno 2019, p. 20190909, DOI : 10.1098/rspb.2019.0909 . URL consultato il 24 giugno 2019 .
- ^ National Geographic Society, Il Tyrannosaurus rex aveva un fiuto incredibile , su National Geographic . URL consultato il 24 giugno 2019 (archiviato dall' url originale il 24 giugno 2019) .
- ^ Grant S. Hurlburt, Ryan C. Ridgely e Lawrence M. Witmer, Chapter 6: Relative size of brain and cerebrum in Tyrannosaurid dinosaurs: an analysis using brain-endocast quantitative relationships in extant alligators , in Michael J. Parrish, Ralph E. Molnar, Philip J. Currie e Eva B. Koppelhus (a cura di), Tyrannosaurid Paleobiology (Life of the Past) , Indiana University Press, 5 luglio 2013 [This volume originated in a conference held on September 16–18, 2005, titled 'The Origin, Systematics, and Paleobiology of Tyrannosauridae,' and sponsored by the Burpee Museum of Natural History and Northern Illinois University] , pp. 134-154, ISBN 978-0-253-00947-0 . URL consultato il 20 ottobre 2013 .
- ^ http://www.blitzquotidiano.it/scienza-e-tecnologia/t-rex-sapeva-parlare-mappa-cervello-dinosauri-svela-i-comportamenti-complessi-1717502/
- ^ Brian Switek, The Tyrannosaurus Rex's Dangerous and Deadly Bite , su Smithsonian.com , ottobre 2012.
- ^ [4] Archiviato il 19 agosto 2018 in Internet Archive ., Bates, KT & Falkingham PL (2012). Estimating maximum bite performance in Tyrannosaurus rex using multi-body dynamics . Biological Letters. DOI : 10.1098/rsbl.2012.0056
- ^ Crispian Scully, (2002) Oxford Handbook of Applied Dental Sciences , Oxford University Press – ISBN 978-0-19-851096-3 P156
- ^ Lambe, LB, The Cretaceous theropodous dinosaur Gorgosaurus , in Memoirs of the Geological Survey of Canada , vol. 100, 1917, pp. 1-84, DOI : 10.4095/101672 .
- ^ Farlow, JO and Holtz, TR, The fossil record of predation in dinosaurs , in Kowalewski, M. and Kelley, PH (a cura di), The Fossil Record of Predation ( PDF ), The Paleontological Society Papers, vol. 8, 2002, pp. 251-266. URL consultato il 14 giugno 2010 (archiviato dall'originale il 31 ottobre 2008) .
- ^ Novella, S. "Interview with Jack Horner." The Skeptics Guide to the Universe. October 14, 2011. Accessed October 24, 2011, https://media.libsyn.com/media/skepticsguide/skepticast2009-10-14.mp3
- ^ The king of killers or fearsome freeloader? It is one of the big questions currently in palaeontology.
- ^ GS Paul, Predatory Dinosaurs of the World , Simon and Schuster, 1988, ISBN 0-671-61946-2 , OCLC 18350868 .
- ^ GD Ruxton e DC Houston, Could Tyrannosaurus rex have been a scavenger rather than a predator? An energetics approach , in Proceedings. Biological sciences / the Royal Society , vol. 270, n. 1516, aprile 2003, pp. 731-3, DOI : 10.1098/rspb.2002.2279 , PMC 1691292 , PMID 12713747 . URL consultato il 5 ottobre 2008 .
- ^ Chin, K., Erickson, GM et al., A king-sized theropod coprolite , in Nature , vol. 393, n. 6686, 18 giugno 1998, p. 680, DOI : 10.1038/31461 . Summary at R. Monastersky, Getting the scoop from the poop of T. rex , in Science News , vol. 153, n. 25, 20 giugno 1998, p. 391, DOI : 10.2307/4010364 , JSTOR 4010364 . URL consultato il 5 maggio 2019 (archiviato dall' url originale il 1º novembre 2012) .
- ^ a b Martin Walters, Bloomsbury Illustrated Dictionary of Prehistoric Life (Bloomsbury Illustrated Dictionaries) , Godfrey Cave Associates Ltd, 1995, ISBN 1-85471-648-4 .
- ^ K. Carpenter , Evidence of predatory behavior by theropod dinosaurs , in Gaia , vol. 15, 1998, pp. 135-144. URL consultato il 5 dicembre 2007 (archiviato dall' url originale il 17 novembre 2007) .
- ^ John Happ e Kenneth Carpenter, An analysis of predator–prey behavior in a head-to-head encounter between Tyrannosaurus rex and Triceratops , in Carpenter, Kenneth; and Larson, Peter E. (editors) (a cura di), Tyrannosaurus rex, the Tyrant King (Life of the Past) , Bloomington, Indiana University Press, 2008, pp. 355 –368, ISBN 0-253-35087-5 .
- ^ Darren H. Tanke e Philip J. Currie, Head-biting behavior in theropod dinosaurs: paleopathological evidence ( PDF ), in Gaia , n. 15, 1998, pp. 167-184, ISSN 0871-5424 .
- ^ DOI : 10.1098/rspb.2010.2497
- ^ Eric. Snively, John R. Cotton, Ryan Ridgely e Lawrence M. Witmer, Multibody dynamics model of head and neck function in Allosaurus (Dinosauria, Theropoda) , in Palaeontologica Electronica , vol. 16, n. 2, 2013.
- ^ Longrich N R., Horner JR, Erickson GM & Currie PJ (2010), "Cannibalism in Tyrannosaurus rex" , Public Library of Science .
- ^ Copia archiviata , su discoveryuk.com . URL consultato il 28 luglio 2013 (archiviato dall' url originale il 19 gennaio 2012) . , Discover Ed (June 22, 2011) Dino Gangs . Retrieved on July 28, 2013 from Copia archiviata , su discoveryuk.com . URL consultato il 28 luglio 2013 (archiviato dall' url originale il 19 gennaio 2012) .
- ^ Nick Collins, Tyrannosaurus Rex 'hunted in packs' , in The Telegraph , 22 giugno 2011. URL consultato il 23 marzo 2014 .
- ^ a b Copia archiviata , su discoveryuk.com . URL consultato il 28 luglio 2013 (archiviato dall' url originale il 19 gennaio 2012) . , Discover Ed(June 22, 2011) Dino Gangs. Retrieved on July 28, 2013 from Copia archiviata , su discoveryuk.com . URL consultato il 28 luglio 2013 (archiviato dall' url originale il 19 gennaio 2012) .
- ^ [5] , Wallis, Paul(June 11, 2012) Op-Ed: T. Rex pack hunters? Scary, but likely to be true. Retrieved on July 28, 2013 from http://digitaljournal.com/article/326451
- ^ Switek, B. (2011). "A bunch of bones doesn't make a gang of bloodthirsty tyrannosaurs: Evidence presented in Dino Gangs that tyrannosaurs hunted cooperatively is a triumph of PR over hard scientific evidence. " The Guardian , July 25, 2011. Accessed online July 29, 2013, http://www.guardian.co.uk/science/blog/2011/jul/25/bunch-bones-gang-bloodthirsty-tyrannosaurs
- ^ Lawrence Witmer, Dino Gangs: solitary, communal, or cooperative hunting in tyrannosaurs , su witmerlab.wordpress.com , Pick & Scalpel WitmerLab at Ohio University, 13 luglio 2011. URL consultato il 12 ottobre 2013 .
- ^ Ian Sample, Researchers find first sign that tyrannosaurs hunted in packs , su The Guardian , 23 luglio 2014. URL consultato il 28 luglio 2014 .
- ^ DOI : 10.1371/journal.pone.0103613
- ^ Dinosauro.eu, Rituali di corteggiamento tra dinosauri , su dinosauro.eu .
- ^ Wolff EDS, Salisbury SW, Horner JR, Varricchio DJ, Common Avian Infection Plagued the Tyrant Dinosaurs . PLoS ONE 4(9) (2009): e7288 online
- ^ EDS Wolff, SW Salisbury, JR Horner, DJ Varricchio, Common Avian Infection Plagued the Tyrant Dinosaurs . PLoS ONE, 4(9),(2009)
- ^ Più precisamente nutrimento simpatrico o conspecifico di Tyrannosauridae
- ^ Preparation and mounting , su fieldmuseum.org . URL consultato il 10 giugno 2010 (archiviato dall' url originale il 17 marzo 2009) .
- ^ ( EN ) Erickson, G., Makovicky, PJ, Currie, PJ, Norell, M., Yerby, S., Brochu, CA, Gigantism and life history parameters of tyrannosaurid dinosaurs , in Nature , vol. 430, maggio 2004. URL consultato il 10 giugno 2010 .
«no» . - ^ Dig pulls up five T. rex specimens , su news.bbc.co.uk . URL consultato il 10 giugno 2010 .
Bibliografia
- Brochu, CR 2003. Osteology of Tyrannosaurus rex: insights from a nearly complete skeleton and high-resolution computed tomographic analysis of the skull. Memoirs of the Society of Vertebrate Paleontology. 7: 1-138.
- AA.vv. Dinosauri e altri animali preistorici, Giorgio Mondadori ed., pp. 256 257
- Henderson, DM 1999. Estimating the masses and centers of mass of extinct animals by 3-D mathematical slicing. Paleobiology 25: 88–106.
- Farlow, JO, Smith, MB, & Robinson, JM 1995. Body mass, bone "strength indicator", and cursorial potential of Tyrannosaurus rex. Journal of Vertebrate Paleontology 15: 713-725
- Carr, TD & Williamson, TE 2004. Diversity of late Maastrichtian Tyrannosauridae (Dinosauria: Theropoda) from western North America. Zoological Journal of the Linnean Society 142: 479–523
- Osborn, HF 1905.Tyrannosaurus and other Cretaceous carnivorous dinosaurs. Bulletin of the American Museum of Natural History 21;259–265
- Horner John R - Padian Kvin, 2004. Age an growth dynamics of Tyrannosaurus rex, Royal Society Publishing, Volume 271, Numero 1551/22 settembre 2004 Pagg. 1875-1880
- Alexander, RM 1997 Size and scaling. In "The encyclopedia of dinosaurs" (ed. P. Currie & K. Padian), pp. 665–667. San Diego, CA: Academic.
- Schweitzer Mary Higby, Suo Zhiyong, Avci Recep, Asara John M., Allen Mark A., Arce Fernando Teran, Horner John R. 2007. Analyses of Soft Tissue from Tyrannosaurus rex Suggest the Presence of Protein. In "Science", Vol. 316. no. 5822, pp. 277 – 280, 13 aprile 2007, DOI: 10.1126/science.1138709
- Carpenter Kenneth, 1990. Variation in Tyrannosaurus Rex, In: Carpenter, K. & Currie, PJ (Eds.). Dinosaur Systematics: Approaches and Perspectives. New York: Cambridge University
- Carpenter Kennet, Smith Matt, 2001. Forelimb osteology and biomechanics of Tyrannosaurus Rex. In: Tanke D., Carpenter K. "Mesozoic vertebrate life", ed. Indiana university press, Bloomington, Indiana, 2001.
- Erickson, GM, Makovicky, PJ, Currie, PJ, Norell, MA, Yerby, SA, & Brochu, CA 2004. Gigantism and comparative life-history parameters of tyrannosaurid dinosaurs. Nature 430: 772–775.
- Appenzler, T., T. rex Was Fierce, Yes, But Feathered, Too. Science vol. 258/n.5436, pp. 2052–2053, 24 settembre 1999.
- Xu X., Norell, MA, Kuang X., Wang X., Zhao Q., & Jia C. 2004. Basal tyrannosauroids from China and evidence for protofeathers in tyrannosauroids. Nature 431, pp 680–684.
- Sumida SS, Brochu CA Phylogenetic Context for the Origin of Feathers. American Zoology, vol.40, Numero 4, pp. 475–485, Agosto 2000, DOI 101093
- Ruxton GD, Huston DC, 2003. Could Tyrannosaurus rex have been a scavenger rather than a predator? An energetics approach. The royal society, Biological Sciences, vol. 270, pp. 731–733, 7 aprile 2003
- Horner JR, 1994. Steak knives, beady eyes, and tiny little arms (a portrait of T. rex as a scavenger), Special Publication – The Paleontological Society, v.7, pp. 157–164.
- Horner JR, Lessem D.; The Complete T-rex; Simon & Schuster, 1993; ISBN 0-671-74185-3 , 9781741853.
- Lingham-Soliar T., 2002. Guess who's coming to dinner: A portrait of Tyrannosaurus as a predator. Geology today, v.14, pp. 16–20, 4 gennaio 2002
Altri progetti
-
 Wikiquote contiene citazioni sul tirannosauro
Wikiquote contiene citazioni sul tirannosauro -
 Wikibooks contiene un libro per bambini con un capitolo su Tyrannosaurus
Wikibooks contiene un libro per bambini con un capitolo su Tyrannosaurus -
 Wikizionario contiene il lemma di dizionario « tirannosauro »
Wikizionario contiene il lemma di dizionario « tirannosauro » -
 Wikiversità contiene risorse su Tyrannosaurus rex
Wikiversità contiene risorse su Tyrannosaurus rex -
 Wikimedia Commons contiene immagini o altri file su Tyrannosaurus
Wikimedia Commons contiene immagini o altri file su Tyrannosaurus -
 Wikispecies contiene informazioni su Tyrannosaurus
Wikispecies contiene informazioni su Tyrannosaurus
Collegamenti esterni
- ( EN ) Tyrannosaurus rex , su Enciclopedia Britannica , Encyclopædia Britannica, Inc.
- ( EN ) Tyrannosaurus rex , su Fossilworks.org .