Ankylosaurus magniventris
| Ankilosaur | |
|---|---|
 | |
| Gama geologică | |
| Starea de conservare | |
Fosil | |
| Clasificare științifică | |
| Domeniu | Eukaryota |
| Regatul | Animalia |
| Phylum | Chordata |
| Clasă | Sauropsida |
| Superordine | Dinozauria |
| Ordin | † Ornitischia |
| Subordine | † Thyreophora |
| Familie | † Ankylosauridae |
| Subfamilie | † Ankylosaurinae |
| Trib | † Ankilozaurini |
| Tip | † Ankilosaur Brown , 1908 |
| Nomenclatura binominala | |
| † Ankylosaurus magniventris Brown, 1908 | |
Ankylosaurus ( Brown , 1908 - al cărui nume din limba greacă înseamnă literalmente „șopârlă topită” [1] ) este un gen dispărut de dinozaur ornithischian tirofor trăit în Cretacicul superior , cu aproximativ 70-65 milioane de ani în urmă ( Maastrichtian ), în ceea ce este astăzi America de Nord , ceea ce îl face unul dintre ultimii dinozauri care au apărut înainte de marea dispariție a dinozaurilor . Ankylosaurus a fost descris pentru prima dată în 1908 de paleontologul Barnum Brown , iar specia tip , A. magniventris , este singura specie atribuită genului Ankylosaurus . De la descoperirea sa, au fost descoperite doar o mână de exemplare fragmentare, dar nu s-a găsit niciodată un schelet complet. Deși majoritatea membrilor Ankylosauria sunt reprezentați de materiale fosile mult mai complete, Ankylosaurus este adesea considerat ca arhetipul grupului său.
Cu o lungime maximă de 6,25 m (20,5 ft), o înălțime de 1,7 m (5,6 ft) și o greutate de 6 tone (13,000 lb), în ciuda materialului fosil redus, Ankylosaurus este încă considerat astăzi cel mai mare gen de Ankylosauria . Corpul său lat și robust era susținut de patru picioare scurte, dar puternice, și era acoperit cu o armură groasă de osteoderme , dispuse în inele care înconjurau și regiunea gâtului. Coada sa lungă, blindată, s-a încheiat cu o coadă mare, formată din oase topite. Capul său lat și scăzut avea două coarne mici care se ramificau în spate și o altă pereche de aceeași formă, dar care în schimb arăta în jos. Partea din față a fălcilor era acoperită cu un cioc excitat, echipat cu un șir de dinți mici în formă de frunze. Oasele craniului au fost topite, ceea ce a dat o rezistență mai mare oaselor, această caracteristică dă și numele genului.
Ankylosaurus este un membru al familiei Ankylosauridae , iar rudele sale cele mai apropiate par a fi Anodontosaurus și Euoplocephalus . Datorită armurii sale masive, Ankylosaurus nu trebuia să fie un animal foarte rapid, deci se crede că este un animal lent, capabil să facă mișcări rapide și rapide doar atunci când este necesar. Botul larg și aplatizat indică faptul că era un erbivor, nu foarte selectiv capabil să mestece și să rupă orice tip de legume. Sinusurile și cavitățile nazale au fost dezvoltate în mod excepțional, ceea ce indică un simț al mirosului foarte dezvoltat, precum și faptul că i-a făcut pe paleontologi să presupună că animalul ar putea emite diferite tipuri de vocalizări. Marele baston caudal plasat la capătul cozii lui Ankylosarus a fost probabil folosit de animal în viață ca armă de apărare și ofensare împotriva prădătorilor sau în lupte intraspecifice . Fosilele acestui animal au fost găsite în formațiunile Hell Creek, Lance și Scollard, dar în toate formațiunile se pare că a fost un animal destul de rar în habitatul său. Deși a trăit lângă un alt anchilozaur Edmontonia , se pare că animalele nu au intrat niciodată în competiție deoarece ocupau nișe ecologice diferite, de fapt, ankilosaurul pare să fi locuit în cele mai montane zone. Acest animal a trăit în aceleași locuri și în același timp în care au trăit alți uriași precum Tiranosaurul , Triceratopsul și Edmontosaurul .
Descriere

Ankylosaurus este cel mai mare dinozaur ankilozaurid cunoscut, [2] cu o lungime maximă de 6,25 m (20,5 ft), o lățime de 1,5 m (4,9 ft) și o înălțime de 1,7 m (5,6 picioare) la nivelul șoldului. Aceste estimări au fost publicate de paleontologul american Kenneth Carpenter , pe baza celui mai mare craniu cunoscut (NMC 8880), măsurând aproximativ 64,5 cm (25,4 in) în lungime și 74,5 cm (29,3 in) în lățime. Cel mai mic craniu cunoscut (AMNH 5214), pe de altă parte, măsoară doar 55,5 cm (21,9 in) în lungime și 64,5 cm (25,4 in) în lățime, pentru acest exemplar o lungime de aproximativ 5,4m (18ft) în lungime și aproximativ 1,4 m (4,6 ft) în înălțime. [3] Alți autori au propus o lungime a corpului de aproximativ 7 metri (23 picioare) sau 8-9 m (26-30 picioare) sau mai mult de 9 metri (30 picioare). Greutatea animalului este estimată în schimb la aproximativ 6 tone (13.000 lb). [4]

Datorită rămășițelor fragmentare rare, structura multor scheleturi Ankylosaurus , care include pelvisul , partea centrală a cozii și picioarele, este încă necunoscută. Animalul era cu siguranță cvadrupedal , iar membrele posterioare erau mai lungi decât cele din față. Scapula și coracoidul (un os dreptunghiular conectat la capătul inferior al scapulei) ale specimenului AMNH 5895 au fost fuzionate și au posedat entese (țesut conjunctiv) pentru diferite inserții musculare. Omoplatul avea o lungime de 61,5 cm (24,2 in). Humerusul (osul brațului superior) a fost foarte scurt și lat, măsurând aproximativ 54 cm (21 inci) în lungime în exemplarul AMNH 5214. Femurul (osul coapsei) a fost foarte puternic, măsurând aproximativ 67 cm (26 inci) în lungime, în eșantionul AMNH 5214. Cu toate acestea, picioarele animalului sunt complet necunoscute, dar din comparație cu animalele înrudite, picioarele din spate probabil aveau doar trei degete. [5]
Vertebrele cervicale din gât posedau coloane neuronale mari care au crescut în înălțime pe măsură ce se apropiau de corp. Partea anterioară a coloanei vertebrale neuronale avea enteze bine dezvoltate, foarte frecvente la dinozaurii adulți, indicând prezența ligamentelor mari care au ajutat la susținerea greutății capului masiv. Vertebrele dorsale ale spatelui aveau centre (sau corpuri) destul de scurte în raport cu lățimea lor, iar coloanele vertebrale ale acestora erau scurte și înguste. Vertebrele dorsale erau strâns distanțate, ceea ce restricționa mișcarea spatelui în jos. Spinele neuronale aveau tendoane osificate (transformate în os), care se suprapuneau și pe unele vertebre. Coastele ultimelor patru vertebre ale spatelui și ale cutiei toracice erau foarte largi. Coaste prezintă diferite cicatrici care arată atașamentul mușchilor. Vertebrele caudale ale cozii aveau un centru ușor amficoel , adică erau concav pe ambele părți. Procesul articular blocat al vertebrelor caudale a avut o formă vagă de V atunci când este privit de sus. [6]
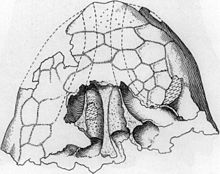
Craniu

Cele trei cranii cunoscute ale lui Ankylosaurus diferă între ele prin diferite caracteristici; se crede în mod obișnuit că aceste caracteristici diferite sunt rezultatul tafonomiei (modificări care apar în timpul decăderii și fosilizării rămășițelor) și a variațiilor individuale . Craniul era jos și de formă triunghiulară, mai lat decât era lung; spatele era, pe de altă parte, lat și jos. Craniul avea un cioc mare pe premaxilla . Orificiile erau aproape rotunde sau ușor ovale și nu erau întoarse direct în lateral, deoarece craniul se înclină spre față. Craniul era scurt și puternic, ca la alte anchilosaurine. Crestele de deasupra orificiilor ochiului s-au contopit în coarnele scuamoase superioare (forma lor a fost descrisă ca „ piramidală ”), care arătau spre părțile laterale din spatele craniului: creasta și cornul erau probabil elemente separate inițial, așa cum sa menționat în genurile înrudite, cum ar fi Pinacosaurus și Euoplocephalus , în timp ce sub coarnele superioare erau prezenți coarnele jugulare, care arătau înapoi. Se crede că coarnele erau inițial osteoderme (plăci ale armurii) care se topeau cu restul craniului. Modelul asemănător scării prezent pe suprafața dorsală a craniului (numit "caputegulae" la anchilozauri) este rezultatul remodelării craniului în sine, care a obliterat suturile dintre elementele craniului, obișnuite la anchilozaurii adulți. Fiecare craniu pare să aibă propriul model caputegulae pe craniu, deși unele detalii sunt împărtășite. Ankylosaurus poseda, de asemenea, o scară rostrală în formă de diamant (sau hexagonală) în partea din față a botului dintre nări, două osteoderme solzoase deasupra orbitei și o creastă de solzi în partea din spate a craniului. [2] [6] [7]

Regiunea botului Ankylosaurus este unică în rândul ankilozaurilor și a suferit o transformare „extremă” în comparație cu rudele sale. Botul era arcuit și trunchiat în față, iar nările erau eliptice și direcționate în jos și în exterior, spre deosebire de toate celelalte anchilosauride cunoscute care le posedă în față sau în față oblic. Mai mult, nările nu au fost vizibile din față, deoarece sinusurile au fost extinse pe părțile laterale ale osului premaxilar într-o măsură mai mare decât se observă la alți anchilozauri. Osteodermele mari ale caputegoleului - osteodermele laterale ale botului - au acoperit complet deschiderea lărgită a nărilor, conferindu-i un aspect bulbos. Nările au avut, de asemenea, un sept intranar, care a separat pasajul nazal de sinus. Fiecare parte a botului avea cinci sinusuri, dintre care patru s-au extins în maxilar . Cavitățile nazale (sau camerele) lui Ankylosaurus au fost alungite și separate de un sept la linia mediană, care a împărțit interiorul botului în două jumătăți oglindite. Septul avea două deschideri, inclusiv cananele (nările interne). [2] [6]
Fălcile se extind în lateral, dând impresia unei umflături, care s-ar putea datora sinusurilor din interior. Fălcile aveau o creastă care putea reprezenta punctul de atașare pentru obrajii cărnoși; prezența sau absența obrajilor la ornitischieni este oarecum controversată, dar unii anchilozauri nodosauri au plăci de armură care acoperă regiunea obrazului, care ar fi putut fi încorporată în partea cărnoasă. Specimenul AMNH 5214 are 34-35 de alveole dentare în maxilar și are mai mulți dinți decât orice alt anchilosaurid cunoscut. Șirurile de dinți din fălcile acestui exemplar au o lungime de aproximativ 20 de centimetri (8 inci). Fiecare alveolă avea un foramen (deschidere) lângă partea sa unde se vedea un dinte de înlocuire. [6]
În comparație cu alți anchilozauri, maxilarul lui Ankylosaurus era scăzut proporțional cu lungimea sa și, când era privit din lateral, șirul de dinți era aproape drept, mai degrabă decât arcuit. Singura mandibulă complet conservată este prezentă doar la cel mai mic exemplar (AMNH 5214) și are o lungime de aproximativ 41 de centimetri (16 inci). Mandibula incompletă a celui mai mare exemplar (CMN 8880) are aceeași lungime. Specimenul AMNH 5214 are 35 de alveole în stânga și 36 în dentiția dreaptă, pentru un total de 71, cel mai mare număr cunoscut pentru orice alt anchilosaurid. Osul predentar al vârfului maxilarului nu a fost încă găsit. [6] Rândul dinților era relativ scurt. [2] La fel ca alți anchilozauri, Ankylosaurus avea dinți mici, în formă de frunze, care au fost comprimați lateral. [8] Dinții erau în mare parte mai înalți decât largi și erau foarte mici; dimensiunea lor proporțional cu craniul sugerează că maxilarele ar putea găzdui mai mulți dinți decât alte anchilosaurine. Dinții craniului Ankylosaurus mai mare sunt mai mici decât cei ai craniului cel mai mic în sens absolut. Ultimii dinți ai rândurilor se îndoiau înapoi, iar coroanele dinților erau de obicei mai plate pe o parte decât pe cealaltă. [6] Dinții din Ankylosaurus sunt diagnosticați și se pot distinge de dinții altor anchilosauri prin laturile lor netede. Denticulele erau mari, iar numărul lor variază de la șase la opt pe partea din față a dintelui și de la cinci la șapte pe spate. [6] [9]
Osteodermele

O caracteristică proeminentă a Ankylosaurus a fost armura sa, constând din butoane și plăci osoase cunoscute sub numele de osteoderme sau scuturi, încorporate în pielea sa. Aceste structuri nu au fost găsite niciodată în poziția lor inițială, astfel încât locația lor exactă pe corp este necunoscută, deși se pot face presupuneri pe baza animalelor înrudite, pentru care au fost propuse diferite ansambluri. Osteodermele au variat de la 1 centimetru (0,4 in) în diametru la 35,5 centimetri (14,0 in) în lungime și au variat ca formă. Osteodermele din Ankylosaurus erau în general cu pereți subțiri și scobite în partea inferioară. În comparație cu Euoplocephalus , osteodermele Ankylosaurus au fost mai confortabile. Osteodermele și osiculele mici ocupau probabil spațiul dintre cele mai mari. Osteodermele care acopereau corpul erau foarte plate, deși cu o chilă de margine mică. În schimb, nodosauridul Edmontonia poseda chile înalte care se extindeau de la margine la margine pe linia mediană a osteodermelor. Ankylosaurus poseda niște osteoderme mai mici, cu o chilă, de-a lungul liniei medii a corpului. [2] [6]

La fel ca alte anchilozauride, Ankylosaurus poseda semicercuri cervicale (plăci de armură pe partea dorsală a gâtului), dar acestea sunt cunoscute doar din fragmente, ceea ce face ca dispunerea lor exactă să fie incertă. Carpenter a sugerat că, atunci când sunt privite de sus, plăcile ar fi împerecheate, creând o formă de V inversată pe gât, cu golul liniei medii probabil umplut cu osici mici (scuturi rotunde osoase) pentru a permite mișcarea. El credea că lățimea acestei centuri de armură era prea largă pentru a se potrivi singur cu gâtul și că acoperea baza gâtului și continua peste zona umerilor. Arbor și Philip J. Currie nu sunt de acord cu interpretarea lui Carpenter's din 2015 și au subliniat faptul că fragmentele de semicerc cervical AMNH 5895 nu se încadrează în modul propus de Carpenter's (deși acest lucru se poate datora naturii lor fragmentare). În schimb, au sugerat că fragmentele reprezintă rămășițele a două jumătăți de inele cervicale, care formează două plăci semicirculare înarmate în jurul vârfului gâtului, ca în Anodontosaurus și Euoplocephalus . [6] [7] Arbor și Mallon au elaborat această idee, descriind forma acestor jumătăți de inele ca „în formă continuă de U” peste partea superioară a gâtului și au sugerat că Ankylosaurus avea șase osteodermi cu cheile cu baze ovale pe fiecare jumătate de inel. . Alte anchilosauride au adesea multe osteoderme mai mici care le înconjoară pe cele mai mari. [2]
Primele osteoderme din spatele celui de-al doilea semicerc cervical ar fi avut o formă similară cu cele din semicercul , iar osteodermele de pe spate probabil au scăzut în diametru pe măsură ce s-au îndepărtat de gât. Cele mai mari osteoderme erau probabil aranjate în rânduri transversale și longitudinale pe o mare parte a corpului, cu patru sau cinci rânduri transversale separate prin pliuri în piele. Osteodermele de pe șolduri ar fi avut probabil un contur mai pătrat decât cele de pe spate. Este posibil să fi existat patru șiruri longitudinale de osteoderme pe șolduri. Spre deosebire de unii anchilozauri bazali și mulți nodosauri, anchilozaurii nu par să fi avut scuturi pelviene co-osificate deasupra flancurilor. Este posibil ca unele osteoderme fără cheie să fi fost poziționate peste regiunea șoldului Ankylosaurus , ca în Euoplocephalus . Ankylosaurus poate avea alternativ trei sau patru rânduri transversale de osteoderme circulare pe regiunea pelviană, care erau mai mici decât cele din restul corpului, ca în Scolosaurus . Osteodermele triunghiulare mai mici ar putea fi prezente pe părțile laterale ale bazinului. Plăcile turtite și ascuțite seamănă cu cele de pe părțile laterale ale cozii lui Saichania . Osteodermele ovale cu chile pot fi plasate pe partea superioară a cozii sau pe partea membrelor. Osteodermele triunghiulare comprimate găsite cu exemplarele Ankylosaurus pot fi plasate pe părțile laterale ale bazinului sau cozii. Osteodermele ovale, chile și în formă de lacrimă sunt cunoscute la Ankylosaurus și ar fi putut fi plasate pe membrele anterioare, cum ar fi cele cunoscute de la Pinacosaurus , dar nu se știe dacă membrele posterioare aveau osteoderme. [2] [6]

Clubul caudal al lui Ankylosaurus era compus din două osteoderme mai mari, cu un rând de osteoderme mai mici în linia mediană și două osteoderme mici la vârf; aceste osteoderme au ascuns ultima vertebră caudală. Din moment ce buzduganul caudal este cunoscut dintr-un singur exemplar, AMNH 5214, nu se cunoaște intervalul de variație între indivizi. Buzduganul caudal al AMNH 5214 are o lungime de 60cm, o lățime de 49cm și o înălțime de 19cm. Cel mai mare club de specimene ar fi putut ajunge la 57 de centimetri (22 inci) în lățime. Clubul caudal al lui Ankylosaurus era semicircular când era privit de sus, similar cu cel al lui Euoplocephalus și Scolosaurus , dar diferit de clubul ascuțit al Anodontosaurus sau clubul îngust și alungit al Dyoplosaurus . Ultimele șapte vertebre ale cozii au format un „mâner” rigid și solid la club. Aceste vertebre erau în contact, fără cartilaj între ele și uneori erau co-osificate împreună, ceea ce le făcea rigide și imobile. Tendoanele osificate atașate vertebrelor din fața clubului caudal, iar aceste caracteristici împreună au ajutat coada să rămână rigidă și imobilă, făcând-o și mai rezistentă. Zigapofizele interconectate (procesele articulare) și coloanele vertebrale ale vertebrelor mânerului au fost în formă de U atunci când sunt privite de sus, în timp ce cele ale majorității celorlalte anchilosauride au formă de V. Acest lucru se poate datora faptului că mânerul Ankylosaurus a fost mai larg. . O lățime mai mare ar putea indica faptul că coada Ankylosaurus era mai mică decât lungimea corpului sau că avea aceleași proporții ca și celelalte anchilosauride, dar cu un club caudal mai mic. [2] [6] [10]
Clasificare

Brown a considerat Ankylosaurus atât de distinctiv încât a devenit arhetipul unei noi familii , Ankylosauridae (ai cărei membri se numesc ankilosauride), caracterizată prin cranii uriașe triunghiulare, gâturi scurte, spate rigide, corpuri mari și presărate cu osteodermi. De asemenea, a atribuit acestei familii genurile Palaeoscincus (cunoscut doar din dinți) și Euoplocephalus (cunoscut doar dintr-un craniu parțial și unele osteoderme). Datorită naturii fragmentare a rămășițelor, Brown nu a reușit să distingă pe deplin Euoplocephalus și Ankylosaurus . Având în comparație doar câțiva membri incompleti ai familiei, Brown credea că grupul face parte din subordinea Stegosauria [11] . În 1923, Osborn a inventat denumirea de Ankilozaurie (ai cărei membri se numesc anchilozauri), plasând anchilosauridele în propriul lor subordine [12] .
Ankilozauria și Stegosauria sunt acum grupate în cadrul cladei Thyreophora . Acest grup a apărut pentru prima dată în sinemurian și a supraviețuit 135 de milioane de ani până când a dispărut în Maastrichtian . Aceste animale au avut foarte mult succes într-un timp scurt, răspândind și colonizând o gamă largă de medii. [6] [13] Pe măsură ce au fost descoperite specimene mai complete și au fost descrise noi genuri, teoriile despre relația dintre anchilozauri au devenit din ce în ce mai complexe și ipotezele s-au schimbat adesea între studii. În plus față de anchilozauride, Ankilozauria a fost, de asemenea, subdivizată în familiile Nodosauridae și, uneori, Polacanthidae (familii lipsite de cluburi caudale). [14] Ankylosaurus este considerat a face parte din subfamilia Ankylosaurinae (ai cărei membri sunt numiți anchylosaurins) din Ankylosauridae. [14] Ankylosaurus pare a fi strâns legat de Anodontosaurus și Euoplocephalus . [15]

Următoarea cladogramă se bazează pe o analiză filogenetică a Ankylosaurinae, efectuată de Arbor și Currie (2015): [7]
| Ankylosaurinae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Deoarece Ankylosaurus și alte America de Nord cretacicului superior ankylosaurids au fost grupate cu genuri asiatice (într - un trib ce autorii numesc ca Ankylosaurini ), Arbor și Currie a emis ipoteza că mai devreme ankylosaurids din America de Nord au devenit dispărut de târziu Albian sau Cenomanian perioada de Cretacic. Mediu . Ankilosauridele au recolonizat apoi America de Nord din Asia în epoca campaniană sau turoniană din Cretacicul târziu și s-au diversificat din nou, ducând la evoluția unor noi genuri precum Ankylosaurus , Anodontosaurus și Euoplocephalus . Acest lucru ar explica o diferență de 30 de milioane de ani în înregistrările fosile ale anchilozauridelor nord-americane dintre aceste ere. [7]
Paleobiologie
Dietă

La fel ca toți ceilalți ornitischieni, Ankylosaurus era erbivor . Botul larg și plat era perfect potrivit pentru pășunatul selectiv al plantelor joase. Dinții lui Ankylosaurus erau ascuțiți pe latura coroanelor, mai degrabă decât vârfurile coroanelor, ca la nodosauride . [6] În 1982, Carpenter a descoperit doi dinți mici, respectiv de la Lance Formation și Hell Creek Formation, atribuindu-i unui tânăr Ankylosaurus . Cei doi dinți mici au măsurat 3,2 și 3,3 mm lungime, respectiv. Dintele mai mic era foarte uzat, așa că Carpenter a sugerat că anchilosauridele în general sau cel puțin puii nu și-au înghițit mâncarea, ci au folosit un fel de mestecat. [9] Deși anchilozaurii nu s-au hrănit cu materiale vegetale fibroase și plante lemnoase, au avut încă o dietă foarte variată, inclusiv frunze tari și ferigi. [16] Presupunând că este endoterm , Ankylosaurus ar trebui să mănânce aproximativ 60 kg de ferigi pe zi, similar cu cantitatea de vegetație uscată pe care elefantul ar trebui să o consume zilnic. Cu toate acestea, animalul și-ar putea îndeplini cerințele nutriționale mai eficient dacă s-ar hrăni și cu fructe, pe care dinții mici de formă de cuspidă și forma ciocului par bine adaptate deschiderii și dezbrăcării, comparativ cu Euoplocephalus , de exemplu. Chiar și ingestia de nevertebrate mici, pe care dinții săi mici îi pare potrivite pentru manipulare, ar putea oferi nutriție suplimentară. [2]
Dinții de Ankylosaurus erau așezați mai degrabă pe fețele coroanelor lor decât pe vârfurile coroanelor, la fel ca la ankilozaurii nodosauri. [6] În 1982, Carpenter a atribuit doi dinți foarte mici din formațiunile Lance și Hell Creek unui tânăr Ankylosaurus , cu o lungime de 3,2 până la 3,3 milimetri (0,13 până la 0,13 inci), respectiv. Dintele mai mic este foarte uzat, ceea ce l-a determinat pe Carpenter să speculeze că anchilosauridele în general sau cel puțin cele mai tinere nu și-au înghițit mâncarea întreagă, ci au implicat un fel de mestecare. [17] Întrucât un adult Ankylosaurus nu și-ar putea mesteca bine hrana, ar fi petrecut mai puțin timp pe pășunatul zilnic al unui elefant. [2] Pe baza lățimii cuștii toracice, un Ankylosaurus ar putea digera mesele printr-un sistem de fermentare internă, ca niște șopârle moderne erbivore, care au mai multe camere în colonul lor mărit. [6]

În 1969, paleontologul austriac Georg Haas a concluzionat că, în ciuda dimensiunilor mari ale craniilor anchilozaurilor, musculatura asociată cu maxilarele a fost relativ slabă. De asemenea, el a speculat că mișcarea maxilarului era limitată la urcare și coborâre. Extrapolând din aceasta, Haas a sugerat că anchilozaurii se hrăneau cu vegetație moale, non-abrazivă. [18] Cu toate acestea, cercetările ulterioare asupra craniului lui Euoplocephalus au arătat că aceste animale au fost capabile să miște maxilarele chiar și lateral permițând mestecarea, indicând, de asemenea, că craniul a fost capabil să reziste la forțe considerabile. [19] În 2016, un nou studiu privind ocluzia dentară (contactul dintre dinți) și capacitatea de a muta maxilarul înapoi (PALINAL), a fost absent la anchilosauride, cu excepția Ankylosaurus și Euoplocephalus , având dezvoltată această abilitate incredibilă, indiferent de nodosauride. . [20]
Descoperirea unui specimen Pinacosaurus care a păstrat o paraglossalia mare (oase triunghiulare sau cartilaj localizate în limbă) care prezintă semne de stres muscular, iar acest lucru este considerat a fi o caracteristică comună la ankilozauri. Prin urmare, unii cercetători au sugerat că anchilozaurii s-au bazat pe limbi musculare și hyobranchia (oasele limbii) atunci când se hrăneau, deoarece dinții lor erau destul de mici și au fost înlocuiți într-un ritm relativ lent. Unele salamandre moderne au o limbă dezosată în mod similar și își folosesc limba prensilă pentru a ajunge la hrană. Cu o limbă atât de lungă, anchilozaurii ar putea chiar să integreze insecte mici în dieta lor pe care le-au colectat cu limba lungă și lipicioasă. [16] Poziția retrasă a nărilor lui Ankylosaurus a fost comparată cu cea a amfisbenului în vizuină și, deși probabil nu era un animal în vizuină, botul lui Ankylosaurus poate indica o adaptare pentru a mișca pământul. Acești factori, precum și rata scăzută de formare a dinților la ankilozauri în comparație cu alți ornitischieni, indică faptul că Ankylosaurus ar fi putut fi omnivor , hrănindu-se atât cu material vegetal, cât și cu animale mici. Alternativ, și-ar fi putut folosi botul ghemuit pentru a săpa în pământ pentru rădăcini și tuberculi . [2]
Simțurile
În 1977, paleontologa poloneză Teresa Maryanska a propus că complexele sinusale și cavitatea nazală ale anchilozaurilor au funcția de a ușura craniul și că au adăpostit o glandă nazală dezvoltată sau au acționat ca o cameră de rezonanță vocală. [6] [21] Carpenter respinse queste ipotesi, sostenendo che gli animali tetrapodi emettono suoni attraverso la laringe , non attraverso le narici, e che la riduzione del peso sarebbe stata minima, in quanto gli spazi vuoti rappresentano solo una piccola percentuale del volume del cranio. Anche l'ipotesi della ghiandola nasale era piuttosto improbabile, in quanto i seni non hanno alcuna funzione specifica. [6]
Uno studio del 2011 sui passaggi nasali di Euoplocephalus hanno sostenuto la loro funzione come sistema di calore e bilanciamento dell'acqua, notando il vasto sistema di vasi sanguigni e l'aumento della superficie per la membrana mucosa (usato per il calore e l'acqua di scambio negli animali moderni). I ricercatori hanno quindi risuscitato l'ipotesi della camera di risonanza vocale, paragonabile ai passaggi nasali allungati dell' antilope saiga e alla trachea delle gru e dei cigni . Le ricostruzioni dell'orecchio interno suggeriscono un adattamento per sentire i suoni a basse frequenza, come la risonanza a tono basso dei suoni possibilmente prodotti dai passaggi nasali. Tuttavia l'ipotesi della ghiandola nasale è stata scartata in quanto la regione olfattiva si trova ai lati delle vie respiratorie principale. [22]
La forma delle cavità nasali di Ankylosaurus indicano che il flusso d'aria era unidirezionale, (attraverso i polmoni durante inspirazione e l'espirazione), sebbene la bidirezionale sia possibile. [6] L'allargata regione olfattiva degli anchilosauridi indica un senso dell'olfatto molto sviluppato, [22] e la posizione delle orbite di Ankylosaurus suggerire una buona visione stereoscopica . [6]
Locomozione

Le ricostruzioni della muscolatura degli arti anteriori degli ankylosauri, fatta da Coombs nel 1978, suggerisce che gli arti anteriori portassero la maggior parte del peso dell'animale, soprattutto mentre l'animale si nutriva. Inoltre, Coombs ha suggerito che gli anchilosauri fossero capaci di scavare per cercare il cibo, sebbene le loro unghie simili a zoccoli limitassero le loro capacità. La grande e pesante mole dell' Ankylosaurus lo rendevano con tutta probabilità un animale lento e pigro, [23] [24] capace però di movimenti agili e rapidi in caso di necessità. [25]
Ontogenesi
Studi su esemplari di Pinacosaurus di diverse età hanno scoperto che durante lo sviluppo ontogenetico , le costole dei giovani ankylosauri erano fuse con le vertebre. Gli arti anteriori erano molto più robusti degli arti posteriori, ulteriore prova che gli arti anteriori portavano la maggior parte del peso. Durante questa fase della crescita gli osteodermi che formavano la corazza erano ancora in via di formazione e ossificazione, rendendo inoltre l'animale debole agli attacchi dei predatori.[26] Le piastre ossee del cranio, si indurivano poco a poco cominciando prima dal centro del muso e il bordo posteriore; a poco a poco l'ossificazione si estendeva verso le regioni centrali. Sul resto del corpo, il processo di ossificazione progrediva dal collo fino a raggiungere la coda. [27]
Difese

Gli osteodermi degli anchilosauridi erano molto più sottili rispetto a quelle degli altri anchilosauri, e sembrano essere stati rafforzati da dei cuscini distribuiti in modo casuale di collagene e fibre. Questi erano strutturalmente simili alle fibre di Sharpey, ed erano inseriti direttamente nel tessuto osseo, una caratteristica unica degli ankylosauridi. Questo avrebbe fornito agli anchilosauridi un'armatura non solo resistente agli attacchi e ai morsi dei predatori ma anche leggera. [28] Anche le palpebre erano corazzate e rivestite dalle ossa palpebrali, che avrebbero fornito una protezione aggiuntiva. [29] Nel 1982, Carpenter ha suggerito che l'armatura, fortemente vascolarizzata, può anche aver avuto un ruolo nella termoregolazione dell'animale, come nei moderni coccodrilli . [9]
La mazza caudale dell' Ankylosaurus era un'ottima arma offensiva, in grado di produrre un impatto abbastanza forte da rompere le ossa di un possibile aggressore. I tendini della coda erano parzialmente ossificati e non erano molto elastici, trasmettendo alla coda una grande forza per essere usata come arma. [6] Nel 1979, Coombs ha suggerito che i diversi muscoli degli arti posteriori avessero il controllato sull'oscillazione della coda, e che spinta con violenza la mazza caudale avrebbe certamente potuto rompere il metatarso di grandi teropodi come Tyrannosaurus . [24]
Uno studio del 2009 ha stimato che gli anchilosauridi avrebbero potuto far oscillare la coda a 100 gradi lateralmente. Tuttavia, lo studio ha anche riscontrato che solo le grandi mazze caudali degli anchilosauridi erano in grado di rompere le ossa, mentre gli anchilosauri con mazze caudali medie e piccole non ne erano in grado. Nonostante la fattibilità di far oscillare la coda, i ricercatori non hanno potuto determinare se tutti gli anchilosauridi utilizzassero la coda come difesa contro potenziali predatori, nei combattimento intraspecifica o entrambi i casi. [30] Nel 1993, Tony Thulborn ha proposto che la mazza caudale degli ankylosauridi avrebbe agito principalmente come mezzo di distrazione: vedendola agitarsi un predatore avrebbe potuto attaccare la coda scambiandola per la testa in modo che l'anchilosauro potesse colpirlo. Questa teoria potrebbe anche portare a credere che le mazze caudali di questi animali fossero colorate per attirare l'attenzione del predatore o che avessero la stessa colorazione del capo. [31] Sebbene questa idea non sia stata completamente scartata, Carpenter la respinse, in quanto la forma della mazza di questi animali è molto variabile anche tra gli esemplari della stessa specie. [6]
Paleoecologia

L' Ankylosaurus visse tra i 70 ei 65 milioni di anni fa, nella fase finale, o Maastrichtiana , del Cretaceo superiore . Fu uno degli ultimi ultimi generi di dinosauri apparsi prima dell'evento d' estinzione di massa del Cretaceo-Paleogene . L'esemplare tipo dell'animale proviene dalla formazione Hell Creek , nel Montana, mentre altri esemplari sono stati ritrovati nelle formazioni Lance e Ferris, nel Wyoming , nella Formazione Scollard in Alberta e nella formazione Frenchman nello Saskatchewan , tutte formazioni risalenti alla fine del Cretaceo. [2] [4] [32]
I fossili di Ankylosaurus sono piuttosto rari in questi sedimenti e la distribuzione dei suoi resti suggerisce che fosse ecologicamente raro, o limitato agli altopiani delle formazioni piuttosto che alle pianure costiere, dove i suoi resti avrebbero avuto maggiore possibilità di fossilizzarsi. Nello stesso habitat dell'animale, visse anche un altro anchilosauro nodosauride , chiamato Denversaurus (precedentemente denominato Edmontonia sp. ), tuttavia sembra che i due animali non siano mai entrati in competizione per il cibo o il territorio, in quanto quest'ultimo sembra aver preferito le pianure. Inoltre lo stretto muso del Denversaurus suggerisce che quest'ultimo avesse una dieta più selettiva rispetto ad Ankylosaurus , indicando un'ulteriore separazione ecologica . [2] [6]
Con il suo basso centro di gravità, l' Ankylosaurus non sarebbe stato in grado di abbattere alberi come fanno oggi gli elefanti. Era anche incapace di masticare la corteccia, per cui è improbabile che si nutrisse di essa. Una volta adulto, l' Ankylosaurus sembrava prediligere una vita solitaria (anche se alcuni ankylosauri sembrano essersi radunati in gruppi da giovani). Quindi, sebbene fosse un grande erbivoro con requisiti energetici simili, è improbabile che l' Ankylosaurus fosse in grado di modificare il paesaggio del suo ecosistema, come fanno oggi gli elefanti; questo ruolo di " ingegneri dell'ecosistema " era forse ricoperto dagli hadrosauridi . [2]

Le formazioni in cui sono stati rinvenuti fossili di Ankylosaurus rappresentano diverse sezioni della sponda occidentale del mare interno occidentale che divideva l'America settentrionale in occidentale e orientale durante il Cretaceo, che all'epoca rappresentavano ampie pianure costiere che si estendevano dalla costa ovest alle neonate Montagne Rocciose . Queste formazioni sono composte in gran parte da arenaria e argilla , attribuite ad ambienti delle pianure alluvionali . [5] [33] [34] Le regioni in cui sono stati trovati i fossili di Ankylosaurus e altri ankylosauri del tardo Cretaceo avevano un clima caldo subtropicale / temperato , ma anche monsonico , con piogge occasionali, tempeste tropicali e incendi boschivi . [20] Nella formazione di Hell Creek, crescevano diversi tipi di piante, principalmente angiosperme , con conifere meno comuni, felci e cicadee . Un'abbondanza di foglie fossili trovate in dozzine di siti diversi indica che l'area era in gran parte boschiva. [35] L' Ankylosaurus condivideva il suo habitat con altri dinosauri che includevano i ceratopsidi Triceratops e Torosaurus , il parksosauride Thescelosaurus , l' hadrosauride Edmontosaurus , il nodosauride Denversaurus , il pachycephalosauro Pachycephalosaurus , ei teropodi Anzu , Struthiomimus , Ornithomimus , Dakotaraptor , Acheroraptor , diversi troodontidi , e Tyrannosaurus . [3] [4]
Nella cultura di massa
L' Ankylosaurus è una delle creature addomesticabili presenti nel videogioco Ark: Survival Evolved .
Galleria d'immagini

Cranio

Mazza caudale

Ricostruzione antiquata dello scheletro basata sui primi ritrovamenti del 1908

Modello di Anchilosauro


Note
- ^ ( EN ) Definition of ankylosaurus , su merriam-webster.com . URL consultato il 2 marzo 2018 .
- ^ a b c d e f g h i j k l m n VM Arbour e JC Mallon, Unusual cranial and postcranial anatomy in the archetypal ankylosaur Ankylosaurus magniventris , in FACETS , vol. 2, n. 2, 2017, pp. 764–794, DOI : 10.1139/facets-2017-0063 .
- ^ a b Bigelow, P., Cretaceous 'Hell Creek Faunal Facies'; Late Maastrichtian , su scn.org . URL consultato il 24 marzo 2014 (archiviato dall' url originale il 24 gennaio 2007) .
- ^ a b c Weishampel, DB; Barrett, PM; Coria, RA; Le Loeuff, J.; Xu X.; Zhao X.; Sahni, A.; Gomani, EMP; Noto, CR, Dinosaur Distribution , in Weishampel, DB; Dodson, P.; Osmolska, H.. (a cura di), The Dinosauria (2nd) , University of California Press, 2004, pp. 517 –606, ISBN 0-520-24209-2 .
- ^ a b Eberth, DA, Edmonton Group , in Currie, PJ; Padian, K. (a cura di), The Encyclopedia of Dinosaurs , Academic Press, 1997, pp. 199 –204, ISBN 978-0-12-226810-6 .
- ^ a b c d e f g h i j k l m n o p q r s t u v Carpenter, K., Redescription of Ankylosaurus magniventris Brown 1908 (Ankylosauridae) from the Upper Cretaceous of the Western Interior of North America , in Canadian Journal of Earth Sciences , vol. 41, n. 8, 2004, pp. 961–86, Bibcode : 2004CaJES..41..961C , DOI : 10.1139/e04-043 .
- ^ a b c d Arbour, VM e Currie, PJ, Systematics, phylogeny and palaeobiogeography of the ankylosaurid dinosaurs , in Journal of Systematic Palaeontology , vol. 14, n. 5, 2015, pp. 1–60, DOI : 10.1080/14772019.2015.1059985 .
- ^ Coombs, W., Teeth and taxonomy in ankylosaurs , in Carpenter, K. Currie, PJ (a cura di), Dinosaur systematics: Approaches and perspectives , Cambridge University Press, 1990, pp. 269–79, ISBN 978-0-521-43810-0 .
- ^ a b c Carpenter, K., Skeletal and dermal armor reconstruction of Euoplocephalus tutus (Ornithischia: Ankylosauridae) from the Late Cretaceous Oldman Formation of Alberta , in Canadian Journal of Earth Sciences , vol. 19, n. 4, 1982, pp. 689–97, Bibcode : 1982CaJES..19..689C , DOI : 10.1139/e82-058 .
- ^ VM Arbour e PJ Currie, Ankylosaurid dinosaur tail clubs evolved through stepwise acquisition of key features , in Journal of Anatomy , vol. 227, n. 4, 2015, pp. 514–23, DOI : 10.1111/joa.12363 , PMID 26332595 .
- ^ Brown, B., The Ankylosauridae, a new family of armored dinosaurs from the Upper Cretaceous , in Bulletin of the American Museum of Natural History , 24, 1908, pp. 187–201.
- ^ Osborn, HF, Two Lower Cretaceous dinosaurs of Mongolia , in American Museum Novitates , vol. 95, 1923, pp. 1–10.
- ^ Coombs, W., The families of the ornithischian dinosaur order Ankylosauria ( PDF ), in Journal of Paleontology , vol. 21, n. 1, 1978, pp. 143–170. URL consultato il 2 marzo 2018 (archiviato dall' url originale l'8 luglio 2015) .
- ^ a b RS Thompson, JC Parish, SCR Maidment e PM Barrett, Phylogeny of the ankylosaurian dinosaurs (Ornithischia: Thyreophora) , in Journal of Systematic Palaeontology , vol. 10, n. 2, 2012, pp. 301–312, DOI : 10.1080/14772019.2011.569091 .
- ^ VM Arbour, PJ Currie e D. Badamgarav, The ankylosaurid dinosaurs of the Upper Cretaceous Baruungoyot and Nemegt formations of Mongolia , in Zoological Journal of the Linnean Society , vol. 172, n. 3, 2014, pp. 631–652, DOI : 10.1111/zoj.12185 .
- ^ a b Hill, RV, D'Emic, MD, Bever, GS e Norell, MA, A complex hyobranchial apparatus in a Cretaceous dinosaur and the antiquity of avian paraglossalia , in Zoological Journal of the Linnean Society , vol. 175, n. 4, 2015, pp. n/a, DOI : 10.1111/zoj.12293 .
- ^ K. Carpenter, Baby dinosaurs from the Late Cretaceous Lance and Hell Creek formations and a description of a new species of theropod , in Rocky Mountain Geology , vol. 20, n. 2, 1982, pp. 123–134.
- ^ ( EN ) Haas, G., On the jaw musculature of ankylosaurs , in American Museum Novitates , vol. 2399, 1969, pp. 1–11.
- ^ Rybczynski, N. e Vickaryous, MK, Chapter 14: Evidence of Complex Jaw Movement in the Late Cretaceous Ankylosaurid, Euoplocephalus tutus (Dinosauria: Thyreophora) , in K. Carpenter (a cura di), The Armored Dinosaurs , Indiana University Press, 2001, pp. 299–317, ISBN 978-0-253-33964-5 .
- ^ a b A. Ősi, E. Prondvai, J. Mallon e ER Bodor, Diversity and convergences in the evolution of feeding adaptations in ankylosaurs (Dinosauria: Ornithischia) , in Historical Biology , 2016, pp. 1–32, DOI : 10.1080/08912963.2016.1208194 .
- ^ Maryanska, T., Ankylosauridae (Dinosauria) from Mongolia ( PDF ), in Palaeontologia polonica , vol. 37, 1977, pp. 85–151.
- ^ a b Miyashita, T., Arbour VM; Witmer LM; Currie, PJ, The internal cranial morphology of an armoured dinosaur Euoplocephalus corroborated by X-ray computed tomographic reconstruction ( PDF ), in Journal of Anatomy , vol. 219, n. 6, 2011, pp. 661–75, DOI : 10.1111/j.1469-7580.2011.01427.x , PMC 3237876 , PMID 21954840 (archiviato dall' url originale il 24 settembre 2015) .
- ^ Coombs, W., Forelimb muscles of the Ankylosauria (Reptilia, Ornithischia) , in Journal of Paleontology , vol. 52, n. 3, 1978, pp. 642–57, JSTOR 1303969 .
- ^ a b Coombs, W., Osteology and myology of the hindlimb in the Ankylosauria (Reptillia, Ornithischia) , in Journal of Paleontology , vol. 53, n. 3, 1979, pp. 666–84, JSTOR 1304004 .
- ^ Coombs, WP, Theoretical aspects of cursorial adaptations in dinosaurs , in The Quarterly Review of Biology , vol. 53, n. 4, 1978, pp. 393–418, DOI : 10.1086/410790 .
- ^ M Burns, T Tumanova e P Currie, Postcrania of juvenile Pinacosaurus grangeri (Ornithischia: Ankylosauria) from the Upper Cretaceous Alagteeg Formation, Alag Teeg, Mongolia: implications for ontogenetic allometry in ankylosaurs , in Journal of Paleontology , vol. 89, n. 1, Jan 2015, pp. 168–182, DOI : 10.1017/jpa.2014.14 .
- ^ Currie, PJ; Badamgarav, D.; Koppelhus, EB; Sissons, R.; Vickaryous, MK, Hands, feet, and behaviour in Pinacosaurus (Dinosauria: Ankylosauridae) , in Acta Palaeontologica Polonica , vol. 56, n. 3, 2011, pp. 489–504, DOI : 10.4202/app.2010.0055 .
- ^ Scheyer, TM e Sander, PM, [0874:hoaoif 2.0.co;2 Histology of ankylosaur osteoderms: implications for systematics and function ], in Journal of Vertebrate Paleontology , vol. 24, n. 4, 2004, pp. 874–93, DOI : 10.1671/0272-4634(2004)024[0874:hoaoif]2.0.co;2 , JSTOR 4524782 .
- ^ Coombs W., The Bony Eyelid of Euoplocephalus (Reptilia, Ornithischia) , in Journal of Paleontology , vol. 46, n. 5, 1972, pp. 637–50, JSTOR 1303019 . .
- ^ Arbour, VM,Estimating impact forces of tail club strikes by ankylosaurid dinosaurs , in PLoS ONE , vol. 4, n. 8, 2009, p. e6738, Bibcode : 2009PLoSO...4.6738A , DOI : 10.1371/journal.pone.0006738 , PMC 2726940 , PMID 19707581 .
- ^ Thulborn, T., Mimicry in ankylosaurid dinosaurs , in Record of the South Australian Museum , 27, 1993, pp. 151–58.
- ^ Vickaryous, MK, Maryanska, T.; Weishampel, DB, Ankylosauria , in Weishampel, DB, Dodson, P. e Osmólska, H. (a cura di), The Dinosauria , University of California Press, 2004, pp. 363 –92, ISBN 0-520-24209-2 .
- ^ Lofgren, DF, Hell Creek Formation , in Currie, PJ e Padian, K. (a cura di), The Encyclopedia of Dinosaurs , Academic Press, 1997, pp. 302 –03, ISBN 978-0-12-226810-6 .
- ^ Breithaupt, BH, Lance Formation" , in Currie, PJ; Padian, K. (a cura di), The Encyclopedia of Dinosaurs , Academic Press, 1997, pp. 394 –95, ISBN 978-0-12-226810-6 .
- ^ Johnson, KR, Hell Creek Flora , in Currie, PJ e Padian, K. (a cura di), The Encyclopedia of Dinosaurs , Academic Press, 1997, pp. 300 –02, ISBN 978-0-12-226810-6 .
Altri progetti
-
 Wikimedia Commons contiene immagini o altri file su Ankylosaurus magniventris
Wikimedia Commons contiene immagini o altri file su Ankylosaurus magniventris -
 Wikispecies contiene informazioni su Ankylosaurus magniventris
Wikispecies contiene informazioni su Ankylosaurus magniventris
Collegamenti esterni
- Ankylosaurus magniventris , su sapere.it , De Agostini .
- ( EN ) Ankylosaurus magniventris , su Enciclopedia Britannica , Encyclopædia Britannica, Inc.
- ( EN ) Ankylosaurus magniventris , su Fossilworks.org .
| Controllo di autorità | LCCN ( EN ) sh85005327 |
|---|