Torosaurus latus
| Torosaurus | |
|---|---|
 | |
| Starea de conservare | |
Fosil | |
| Clasificare științifică | |
| Domeniu | Eukaryota |
| Regatul | Animalia |
| Phylum | Chordata |
| Superordine | Dinozauria |
| Ordin | † Ornitischia |
| Familie | † Ceratopsidae |
| Subfamilie | † Chasmosaurinae |
| Trib | † Triceratopsini |
| Tip | † Torosaurus Marsh , 1891 |
| Nomenclatura binominala | |
| † Torosaurus latus Marsh , 1891 | |
| Sinonime | |
| |
| Specii | |
| |
Torosaurus (al cărui nume înseamnă „șopârlă perforată”, referindu-se la deschiderile mari din guler) este un gen dispărut de dinozaur ceratopsid hasmosaurian care a trăit în Cretacicul târziu , cu aproximativ 68-65,95 milioane de ani în urmă, deși este posibil ca „intervalul temporal s-ar fi putut extinde până acum 69 de milioane de ani ( Maastrichtian ) și ale cărui fosile au fost descoperite în interiorul vestic al Americii de Nord , din Saskatchewan până în sudul Texasului . Genul conține două specii : specia tip T. latus și posibila specie T. utahensis . [1]
Torosaurus poseda unul dintre cele mai mari cranii ale oricărui alt animal terestru cunoscut. Întregul craniu, inclusiv gulerul osos mare, a ajuns până la 2,77 metri (9,1 picioare) în lungime. De la cap până la coadă, se crede că animalul are o lungime totală de aproximativ 8-9 m (26-30 de picioare) [2] [3] , pentru 4-6 tone. Torosaurus se distinge de Triceratopsul contemporan pentru gulerul său mai îngust și alungit cu deschideri mari (fenestrae), pentru oasele lungi squamosal care alcătuiesc partea inferioară a gulerului și prezența a cinci sau mai multe perechi de osteodermi sau noduli osoși (epoccipital ) pe spatele gulerului. [4] O altă caracteristică a Torosaurus a fost absența cornului nazal lung văzut în Triceratops prorsus , care seamănă mai mult cu starea văzută în Triceratops horridus cu un corn mai scurt. [4] Trei specii de Torosaurus au fost numite de-a lungul anilor: T. latus , T. gladius și T. utahensis . Cu toate acestea, specia T. gladius nu mai este considerată o specie validă.
În 2010, validitatea Torosaurusului ca gen a fost pusă la îndoială. [5] Un studiu histologic al oaselor fosile combinat cu o investigație a formei gulerului a concluzionat că Torosaurus a reprezentat forma matură a Triceratops , în care oasele specimenelor tipice Triceratops aparțineau exemplarelor imature și a arătat semne ale unei dezvoltări timpurii a deschiderile gulerului Torosaurus . În timpul creșterii, suprafața gulerului s-ar fi întins considerabil și ar apărea deschideri parietale. [6] [7] [8] Cu toate acestea, în 2011, 2012 și 2013, studiile privind caracteristicile externe ale specimenelor fosile cunoscute au arătat că diferențele morfologice dintre cele două genuri exclud sinonimia lor. Principalele probleme cu această ipoteză sunt lipsa unor forme tranzitorii bune, existența aparentă a sub- adulților autentici Torosaurus , diferite proporții craniene independente de maturare și afirmația că formarea deschiderilor în guler la un stadiu adult nu face parte din una normală.secvență de maturare ceratopsică. [4] Cu toate acestea, dezbaterea rămâne încă deschisă în rândul paleontologilor, până la descoperirea de noi date. [9] [10]
Descriere

Toate exemplarele care se referă la Torosaurus sunt animale mari, comparabile cu exemplarele mai mari de Triceratops . Datorită gulerului osos alungit, craniul Torosaurus este unul dintre cele mai mari cranii ale oricărui alt animal terestru cunoscut. Hatcher a estimat o lungime de 2,2 metri pentru craniul exemplarului YPM 1830 și 2,35 metri pentru exemplarul YPM 1831. [11] În 1933, Richard Swann Lull a mărit aceste estimări la 2,4 metri și 2,57 metri. [12] Pe baza acestor estimări, Torosaurus a fost listat ca fiind animalul terestru cu cel mai mare craniu cunoscut. Cu toate acestea, în 1998 Thomas Lehman a susținut că un specimen Pentaceratops posedă un craniu parțial care, în viață, ar fi avut o lungime de 2,9 metri. [13] Acest lucru a fost din nou pus la îndoială de Nicholas Longrich, care, în 2011, a descris acest specimen ca pe un nou gen pe care l-a numit Titanoceratops , concluzionând că craniul său a fost supradimensionat în lungime. [14] Mai mult, în 2006 Andrew Farke a subliniat că noile cranii pe care le-a descris erau în medie chiar mai lungi decât cele două cranii originale ale lui Hatcher: exemplarul MOR 1122 are o lungime de aproximativ 2,52 metri, în timp ce MOR 981 are o lungime de 2,77 metri . [15]

În 2006, Farke a stabilit câteva trăsături de diagnostic ale Torosaurusului . Gulerul este extrem de lung în comparație cu restul craniului. Marginea posterioară, parietală a gulerului avea zece sau mai multe epiparietale ( osteoderme triunghiulare pe marginea parietală a gulerului). Nu există o linie mediană epiparietală; în consecință, nu există osteodermi care se încadrează în limita parietal-squamosal. Osul parietal este subțire și traversat de două deschideri, fenestra parietală, sub formă de deschideri ovale circulare sau transversale. Osul parietal este cu aproximativ 20% mai lat decât este lung. Farke a identificat o singură trăsătură în care T. latus diferă atât de Triceratops horridus cât și de T. utahensis : osul scuamosal avea o creastă vizibilă pe margine cu parietalul combinat cu o depresiune longitudinală profundă paralelă cu acesta. [15]
Farke a subliniat, de asemenea, că exemplarele cunoscute de Torosaurus sunt destul de variabile. De exemplu, coarnele supraorbitale mari (plasate peste orificiile ochiului ) pot fi mari și curbate spre față, ca în exemplarul MOR 981, sau scurte și drepte ca în exemplarele MOR 1122 și ANSP 15191. Poziția coarnelor diferă de specimen cu specimen: acestea se găsesc adesea chiar deasupra orbitelor, în timp ce în exemplarul YPM 1831 își au originea pe marginea posterioară a orbitei. În mod similar, forma cornului nazal este extrem de variabilă: YPM 1831 și, într-o măsură mai mică, YPM 1830 au un corn nazal drept, dar MOR 981, ANSP 15192 și mai ales MOR 1122 posedă doar o umflătură scăzută pe osul nazal. Forma gulerului diferă, de asemenea, de la individ la individ: ANSP 15192 și YPM 1830 au un guler curbat în sus în spate, dar gulerul YPM 1831 este aproape plat, deși acesta poate fi un artefact al restaurării fosilelor. Gulerul YPM 1831 are, de asemenea, o formă de inimă, cu o crestătură mijlocie clară, în timp ce marginea din spate a celorlalte exemplare este dreaptă. Chiar și proporțiile gulerului sunt destul de variabile: în YPM 1831 raportul lungime-lățime este de 1,26, dar MOR 981 are o suprafață de 2,28 ori mai mare decât lățimea. Numărul epiparietalelor este dificil de estimat, deoarece majoritatea fosilelor par să le fi pierdut. MOR 981 și MOR 1122 au 10 și, respectiv, 12 epiparietale. YPM 1831 a fost restaurat cu un fontanel pe acoperișul craniului, care este posibil autentic. Farke a concluzionat, de asemenea, că gradul de variabilitate nu a depășit cel arătat de genurile înrudite. [15]
Farke a subliniat că, în afară de guler, nu s-au găsit diferențe sistematice între Torosaurus și Triceratops . Toate exemplarele Torosaurus sunt similare prin faptul că le lipsește un corn nazal alungit și o canelură arterială orizontală la baza anterioară a cornului nazal, dar fosilele Triceratops cu aceeași combinație de trăsături nu sunt neobișnuite. [15] Hunt, în 2008, a concluzionat că T. utahensis , spre deosebire de T. latus, dar similar cu Triceratops , posedă o linie mediană epiparietală. [16]
Clasificare
În 1891, Marsh l-a plasat pe Torosaurus în familia Ceratopsidae , în Ceratopsia (în greacă: „fețe cu coarne”), [17] un grup de dinozauri erbivori cu ciocuri ca de papagal care au prosperat în America de Nord și Asia în timpul Cretacicului târziu .
Datorită gulerului mare, înalt și îngust, Torosaurus este clasificat în mod tradițional ca Chasmosaurinae , în special ca membru evoluat al liniei Anchiceratops sau Arrhinoceratops . Apoi a fost plasat într-o ramură diferită de Triceratops, care în 1980 a fost considerată un membru al Centrosaurinae , datorită gulerului său scurt și larg. Cu toate acestea, în anii 1990 , o analiză cladistică mai completă a arătat că ambele genuri erau hasmosaurine. Analizele recente arată invariabil o relație strânsă între Torosaurus și Triceratops . [15] [18]
Următoarea cladogramă urmează studiile lui Longrich (2014), care a numit o nouă specie de Pentaceratops și a inclus aproape toate speciile de Chasmosaurini: [19]

| Chasmosaurinae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Descoperire și specii
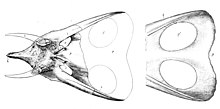
În 1891, la doi ani după denumirea Triceratops , o pereche de cranii ceratopside cu gulere alungite și deschideri mari au fost găsite în sud-estul Wyoming , în județul Niobrara , de către John Bell Hatcher. Angajatorul lui Hatcher, paleontologul profesor Othniel Charles Marsh , le-a inventat genul Torosaurus . [17]
Numele Torosaurus este adesea tradus prin „șopârlă de taur”, datorită și puterii craniului, de la substantivul latin taurus sau de la taurul spaniol , dar mult mai probabil derivă din verbul grecesc τορέω / toreo sau „a străpunge” ). [20] Aluzia este la fenestrele sau deschiderile („asemănătoare unei ferestre”) prezente în guler, care sunt utilizate în mod tradițional pentru a distinge animalul, de Triceratops, care are un craniu mai scurt și mai solid. O mare parte din confuzia asupra etimologiei numelui provine din faptul că Marsh nu a explicat-o niciodată în mod explicit în articolele sale.
Au fost identificate două specii de Torosaurus :
- T. latus Marsh, 1891 ( specie tip ). Numele specific, latus , înseamnă „cel lat” în latină , cu referire la gulerul animalului;
- T. utahensis (Gilmore, 1946) Lawson, 1976
O altă specie a fost considerată ulterior identică cu T. latus :
- T. gladius Marsh, 1891 Numele specific, gladius , înseamnă „sabie” în latină, cu referire la forma alungită a squamosalului.
Specia T. latus se bazează pe holotipul YPM 1830, un craniu parțial. Holotipul lui T. gladius a fost exemplarul YPM 1831, un craniu mai mare. Ambele fosile au fost găsite în formațiunea Lance, datând din Maastrichtian . Specimene fosile similare găsite în Wyoming , Montana , Dakota de Sud , Dakota de Nord , Colorado , Utah și Saskatchewan au fost denumite de atunci genul Torosaurus . Specimenele care pot fi identificate cu o anumită certitudine față de gen includ: ANSP 15192, un individ mai mic din Dakota de Sud descoperit de Edwin Harris Colbert , în 1944; [21] MPM VP6841, un schelet parțial cu craniu, montat acum la Milwaukee ; SMM P97.6.1, un craniu fără bot; și două cranii parțiale din Formația Hell Creek găsite în 2002: MOR 981, descoperit în 1998 și MOR 1122, din 2001. [22] Au fost găsite fragmente în regiunea Big Bend din Texas care ar putea fi atribuite genului, precum și unele fragmente în bazinul San Juan din New Mexico . [23] Paleontologii au observat că exemplarele Torosaurus nu sunt foarte frecvente în înregistrările fosile; Exemplarele Triceratops sunt mult mai abundente.
Specia Torosaurus utahensis a fost inițial descrisă ca Arrhinoceratops utahensis de Charles Whitney Gilmore , în 1946, pe baza USNM 15583, un fragment de guler găsit în comitatul Emery , Utah . [24] În 1976, a fost redenumit Torosaurus utahensis de Douglas Lawson. [25] O recenzie de Robert Sullivan și colab. (2005) [26] și-au confirmat atribuirea genului Torosaurus , deși această specie era ceva mai veche decât T. latus . În 2008, Rebecca Hunt a trimis materiale suplimentare considerabile acestei specii. [16] Nu s-au publicat încă cercetări pentru a considera T. utahensis ca un nou gen sau, așa cum sa sugerat pentru T. latus , faza de creștere a adulților pentru unele specii de Triceratops . [6]
Posibilă sinonimie cu Triceratops
De-a lungul anilor a apărut posibilitatea ca Torosaurus și Triceratops să fie două etape de creștere ale aceluiași animal. În timpul Maastrichtianului din Laramidia au existat două genuri strâns legate de hasmosaurine care împărtășeau același habitat . Singura diferență vizibilă dintre ele a fost forma gulerului. Această ipoteză se bazează pe faptul că nu se cunosc exemplare tinere de Torosaurus , ci dimpotrivă au fost găsite numeroase exemplare tinere de Triceratops . Acesta din urmă se deosebește de alte hasmosaurine prin faptul că menține caracteristicile juvenile chiar și ca adult, cum ar fi scuamosele mici, un caz de pedomorfoză . În 2009, John Scannella, investigând ontogeneza dinozaurilor din Formația Hell Creek din Montana , a concluzionat că aceste date ar putea fi explicate prin ipoteza că Triceratops și Torosaurus au reprezentat diferite etape de creștere ale unui singur gen. În acest punct de vedere, exemplarele de Torosaurus ar fi indivizi complet maturi de Triceratops și, prin urmare, Torosaurus ar fi un sinonim junior al Triceratops , deoarece acesta din urmă are prioritate ca nume. [27]


În 2010, Scanella și Jack Horner , mentorul lui Scannella la Universitatea de Stat din Montana, au publicat cercetări privind modelele de creștere a craniului a treizeci și opt de exemplare (douăzeci și nouă de Triceratops și nouă de Torosaurus ) recuperate în formația Hell Creek, concluzionând că Torosaurus chiar face reprezintă forma adultă a Triceratopsului . [6] Horner a subliniat că gulerul craniilor ceratopsisului este alcătuit din os metaplastic , care are caracteristica de a putea să se lungească și să se scurteze în timp, extinzându-se și reabsorbindu-se pentru a forma forme noi. O dezvoltare semnificativă a acestui lucru poate fi văzută și la acele cranii identificate deja ca Triceratops , unde, potrivit lui Horner, „orientarea cornului este curbată înapoi la exemplarele tinere și apoi se îndoaie la adulți”. Aproximativ 50% din toate craniile sub-adulte Triceratops au două zone subțiri în guler care corespund plasării „deschiderilor” în gulerele Torosaurus , care sunt înconjurate de os granular matur, sugerând că aceste deschideri s-au dezvoltat pentru a ușura craniul, a cărui greutate ar fi crescut la indivizii adulți Triceratops , cu gulere mai lungi. Această ipoteză a făcut parte dintr-un argument și mai larg al lui Horner conform căruia multe presupuse specii de dinozauri ar putea fi de fapt etape de creștere ale altor specii cunoscute. [28] [29] Conform ipotezei lui Horner, prin urmare gulerul indivizilor mai în vârstă de Triceratops s-ar fi lungit considerabil, aplatizându-se și lărgindu-se la marginea din spate. În același timp, ar apărea ferestre parietale, asemănându-se astfel cu forma tipică a gulerului hasmosaurinelor. [6]
Scanella și Horner au recunoscut că nu toate datele ar putea fi ușor explicate prin ipotezele lor. Pentru aceasta au propus câteva ipoteze auxiliare. Una dintre problemele care apar în identificarea materialului Torosaurus ca ultimul stadiu adult al Triceratopsului (denumit stadiu „toromorf”), ne- am aștepta ca fosilele Torosaurus să fie destul de frecvente, în timp ce de fapt sunt destul de rare. Acest lucru a fost explicat de mortalitatea ridicată a subadultilor și de posibilitatea ca animalele vechi și pe deplin dezvoltate să trăiască de preferință la altitudini mari, unde eroziunea împiedică fosilizarea . O a doua problemă a fost gama de dimensiuni a exemplarelor Torosaurus , care părea să sugereze existența unui Torosaurus subadult autentic. Cu toate acestea, Horner și Scanella au declarat că structura osoasă a acestor exemplare a indicat o vârstă complet matură, iar diferențele de mărime au fost rezultatul variațiilor individuale. O a treia obiecție posibilă a fost lipsa aparentă de forme de tranziție între indivizi cu și fără deschideri parietale; aceste ferestre sunt întotdeauna perfect modelate și nu ca perforațiile incipiente. Pentru a explica acest lucru, Horner și Scanella au indicat USNM 2412, holotipul controversatului Nedoceratops , ca exemplu al acestei forme de tranziție. Trăsăturile morfologice problematice de acest fel ar reflecta pur și simplu ființa sa în stadiile incipiente ale transformării sale într-un „toromorf”. O ultimă obiecție la această ipoteză a fost dată de numărul de epoccipitale, de osteodermi sau de noduli osoși de pe marginea gulerului. Cinci epiparietale sunt de obicei prezente în Triceratops , inclusiv un osteoderm de linie mediană; la Torosaurus , în schimb, există zece sau doisprezece, fiind absenți pe linia mediană epiparietală. Numărul de episquamosals pe marginea laterală a gulerului diferă, de asemenea: cinci în Triceratops , șase sau șapte în Torosaurus . Acest lucru a fost explicat prin presupunerea că numărul epoccipitalelor a crescut în timpul creșterii. Mai mult, s-a subliniat că atât numărul, cât și poziția osteodermelor sunt variabile în Triceratops , după cum demonstrează exemplarul MOR 2923, care are șase epiparietale, dar nu are o linie mediană. [6]
Concluziile lui Scannella și Horner nu au fost acceptate în unanimitate. Mai mulți experți, deși au admis posibilitatea că ipoteza „toromorfului” este corectă, au negat că acest lucru este probabil. Ipoteza a fost contestată direct de un articol din 2011 al lui Andrew Farke și un alt articol din 2012 al lui Nicholas Longrich. În 2011, Farke a redescris problematica Nedoceratops hatcheri ca fiind un individ în vârstă sau bolnav de felul său, împotriva identificării lui Scannella și Horner care au susținut identificarea lui Triceratops . Farke a remarcat că găurile neregulate din gulerul lui Nedoceratops , departe de a fi un os subțire, erau înconjurate de umflături îngroșate, sugerând o origine patologică . Farke a concluzionat în continuare că mai multe date erau greu de conciliat cu dezvoltarea propusă de la Triceratops la Torosaurus . În general, numărul epoccipitalelor din ceratopide nu crește odată cu creșterea gulerului. Chiar dacă numărul episoadelor epoccipitale este adesea variabil, pare să nu existe nicio relație cu dimensiunea, deoarece de multe ori tinerii prezintă deja numărul maxim de epoccipitale; se pare că este o variație individuală și nu este legată de ontogeneză. În mod similar, în Ceratopsia în general, deschiderile gulerului sunt prezente de la naștere și nu se formează odată cu creșterea individului. Zonele osoase subțiri din gulerul Triceratops , locația presupusă a găurilor incipiente, a explicat Farke, sunt locuri probabil de atașament muscular. Nu ar exista o relație consistentă între deschideri și o structură osoasă granulară. Multe exemple de Triceratops au gulere cu o suprafață profund venată, ceea ce indică o vârstă considerabilă; pentru a susține schimbarea formei gulerului, osul ar trebui să întinereze din nou, apoi să devină din nou granulat pentru a începe formarea deschiderilor. O secvență pe care Farke a considerat-o puțin probabilă. În cele din urmă, Farke a subliniat că exemplarul YPM 1831, în ciuda dimensiunilor sale mari, nu era încă pe deplin crescut, așa cum arată suturile încă nelegate și structura osoasă netedă, făcându-l astfel un autentic Torosaurus sub-adult. [9]

În același an, Scanella și Horner au răspuns la unele dintre criticile lui Farke. Aceștia au recunoscut că USNM 2412, având în vedere patologiile sale, nu era un candidat ideal pentru o formă de tranziție, dar au subliniat că, pe lângă umflături, găurile de pe gulerul său erau, de asemenea, mărginite de oase granulare și subțiate. Luând în considerare toate datele, au considerat că este mai probabil ca Nedoceratops să reprezinte mai degrabă un individ cu Triceratops decât un gen în sine. De asemenea, au arătat câteva specimene Triceratops care arată exact combinația de os striat venos, granular și juvenil pe care Farke a considerat-o puțin probabilă. De asemenea, au respins ideea că zonele osoase subțiri de pe gulerele Triceratops erau locuri de atașament muscular, deoarece osul din aceste puncte nu arăta suprafața rugoasă tipică atașamentului muscular. Pentru a explica diferența în numărul de epoccipitale, Horner și Scanella au oferit două explicații suplimentare: vârfurile osteodermelor persoanelor în vârstă în timpul vieții s-ar putea să se fi erodat, astfel încât fiecare osteoderm să dea impresia că sunt prezenți doi. În acest fel, cele cinci sau șase epiparietale ale Triceratopsului s-ar fi putut dubla până la zece sau doisprezece, exact cantitatea văzută în exemplarele de Torosaurus . Alternativ, exemplarele de Torosaurus latus , care au fost găsite în straturi mai vechi, ar putea, într-un proces de anageneză , să reprezinte o fază inițială a evoluției Triceratopsului . Cel mai vechi specimen databil, MOR 1122, are douăsprezece epiparietale, în timp ce MOR 981 mai tânăr are zece, ceea ce pare să indice o secvență evolutivă în care numărul de epiparietale a scăzut treptat. [30]

În 2012, Longrich a studiat problema aplicând principiul falsului . Din orice ipoteză științifică valabilă se pot face predicții cu ajutorul cărora pot fi verificate. Longrich a susținut că ipoteza „toromorfului” implica trei astfel de predicții. În primul rând, dacă Torosaurus ar fi identic cu Triceratops , fosilele lor ar fi fost găsite împreună, în aceleași locuri. De fapt, intervalele geografice ale ambelor nu coincid perfect: în nordul îndepărtat nu au fost găsite fosile Torosaurus , în timp ce în sud au fost găsite doar exemplare Torosaurus utahensis . Cu toate acestea, această situație ar putea fi un artefact al relativității rare a rămășițelor Torosaurus și rezultatul eșantionării imperfecte. Longrich a concluzionat apoi că ipoteza a fost coroborată de prima predicție. În al doilea rând, ipoteza a prezis că toate exemplarele Torosaurus erau adulte, în timp ce niciun exemplar Triceratops nu ar putea reprezenta un individ complet matur. Potrivit lui Longrich, ultimul punct nu fusese încă stabilit. Desigur, în 2011 Horner a publicat un studiu histologic care arată că toate exemplarele Triceratops examinate posedă o structură osoasă sub-adultă [31], dar eșantionarea a fost prea mică pentru a permite o generalizare validă pentru toate fosilele Triceratops . Pentru a testa mai bine predicția, Longrich a propus o listă de douăzeci și patru de trăsături ale craniului extern, prin care este posibil să se verifice exemplarele în ceea ce privește nivelul lor de fuziune a elementelor craniului și, prin urmare, creșterea și maturizarea lor. Treizeci și șase specimene au fost examinate aplicând aceste criterii. S-a constatat că fuziunea a avut loc de obicei într-o anumită secvență, oferind informații suplimentare despre vârsta lor. De fapt, conform acestor criterii, majoritatea exemplarelor Torosaurus erau foarte vechi. Cu toate acestea, au existat două excepții. Micul individ ANSP 15192 era, deși adult, încă relativ tânăr, dovadă fiind lipsa fuziunii oaselor botului. Cel mai tânăr exemplar a fost YPM 1831, care avea botul, epiugalul și condilul occipital încă neafuzat; în plus, el pierduse toate osteodermele gulerului, aparent pentru că nu erau încă topite, în timp ce marginea gulerului avea un aspect exterior, tineresc, osos. Pe de altă parte, Longrich a descoperit că zece dintre craniile Triceratops examinate ajunseseră la același nivel de maturare ca exemplarele mai vechi de Torosaurus . Longrich a concluzionat că al doilea test de predicție a infirmat ipoteza. A treia predicție a fost că se pot găsi forme de tranziție între Torosaurus și Triceratops . Longrich a considerat că afirmația că zonele osoase subțiri ale gulerelor Triceratops au fost precursorii fenestrelor parietale ale lui Torosaurus , ca fiind cele mai puternice dovezi ale unei faze de tranziție. Cu toate acestea, el a subliniat că aceste structuri diferă în locație: depresiunile Triceratops se află într-o parte a gulerului situată pe osul squamosal, în timp ce găurile Torosaurus sunt complet înconjurate de osul parietal. În plus, depresiunile Triceratops sunt mărginite de un os mult mai gros, în timp ce găurile Torosaurus sunt înconjurate de os subțire. Longrich a concluzionat că ipoteza a eșuat în ceea ce privește a treia predicție. Fiind infirmată de două dintre cele trei predicții, ipoteza a trebuit să fie respinsă. [4]
Longrich a sugerat, de asemenea, câteva obiecții suplimentare față de ipoteza „toromorfului”: nu se cunosc forme tranzitorii în ceea ce privește numărul de epiparietale. Mai mult decât atât, este dificil să se indice cum numărul lor ar putea crește pe măsură ce ocupă marginea completă a gulerului și că, într-un proces de creștere metaplastică, și-ar crește pur și simplu dimensiunea împreună cu restul gulerului. Ideea divizării osteodermelor prin eroziune fusese stabilită doar pentru episquamosal, dar niciodată pentru epiparietale. Torosaurus are un squamosal îngroșat în partea internă și concav în suprafața externă, în timp ce squamosalul Triceratops este concav în partea internă și plat în partea superioară; formele tranzitorii sunt necunoscute. Squamosalul Torosauros este, de asemenea, indiferent de dimensiunea absolută, mai alungit. Longrich a subliniat că, atunci când exemplarele Torosaurus și Triceratops sunt combinate pentru a crea o singură secvență de creștere, așa cum făcuseră Scanella și Horner, exemplarele Torosaurus , ANSP 15192 și YPM 1831, erau anormale pentru linia de regresie, deoarece squamosalul lor era mai alungit decât ar putea fi explicată prin alometrie . Longrich a recunoscut că faptul că Horner, în studiul său histologic, a reușit să găsească doar exemplare sub-adulte de Triceratops , a fost sugestiv, dar a oferit o explicație alternativă că Triceratops diferea de rudele sale cele mai apropiate prin menținerea unei structuri osoase relativ tinere până la bătrânețe. Pe de altă parte, remodelarea osoasă nu este o figură fiabilă pentru stabilirea maturității, în lumina studiilor experimentale care arată că diferențele în condițiile de deformare mecanică ale diferitelor oase pot modifica semnificativ rata sau gradul unei astfel de remodelări și pot genera iluzia vechilor. tesut osos. Longrich știa că Scanella și Horner vor încerca să contracareze al doilea test al ipotezelor lor, susținând că rezultatele sale sunt compensate de variații individuale. Secondo Longrich, l'importanza di questo fattore era tuttavia limitata: ad esempio, la differenza nelle dimensioni tra ANSP 15192 e YPM 1831 poteva essere spiegata dal dimorfismo sessuale , in cui il primo esemplare era probabilmente una giovane femmina adulta, mentre il secondo era un maschio subadulto. [4]

Nel 2013, Farke e Leonardo Maiorino pubblicarono una ricerca morfometrica, un'analisi statistica del morfospazio (spazio della forma) che descriveva la variazione dei crani di Torosaurus , Triceratops horridus , Triceratops prorsus e Nedoceratops correlati alla crescita e alla maturazione. Il loro studio concluse che i crani di Torosaurus latus durante la maturazione conservavano una forma diversa da T. horridus e T. prorsus , mentre le ultime due specie mostravano una sovrapposizione nelle loro proporzioni. Ciò è vero anche quando la forma del collare viene ignorata. Venne dimostrato che Nedoceratops , con l'eccezione delle dimensioni, non fosse una forma di transizione plausibile tra Torosaurus e Triceratops horridus . Farke e Maiorino ammisero che il basso numero di esemplari di Torosaurus riduce l'affidabilità di questi risultati, ma conclusero che Torosaurus e Triceratops erano taxa separati, pur consentendo la possibilità di anagenesi, ossia che i vari taxa formassero un'unica cronospecie, una linea di discendenza, in mancanza di buoni dati stratigrafici. [10]
L'ipotesi che gli esemplari di Torosaurus latus possano rappresentare una fase "toromorfa" della crescita di Triceratops , ha sollevato la questione secondo cui anche la seconda specie di Torosaurus , Torosaurus utahensis , possa rappresentare una fase "toromorfa". Questo problema è complicato dalla mancanza di materiale fossile sufficiente, in quanto la maggior parte degli esemplari sono costituiti da ossa isolate. T. utahensis era in gran parte riferito a Torosaurus a causa di squamosali allungati, indicando un lungo collare. Il numero di epiparietali, le dimensioni, la posizione o addirittura l'esistenza di fenestre parietali sono sconosciuti. I ricercatori hanno affermato che diversi Torosaurus giovanili sono stati ritrovati in un letto d'ossa nella Formazione Javelina del Parco nazionale di Big Bend , basando la loro identificazione come Torosaurus cfr. utahensis per via della prossimità di un caratteristico parietale di Torosaurus adulto. [16] Scanella e Horner hanno concluso che solo risultati futuri potrebbero risolvere questo problema. Suggerirono, inoltre, che questo taxon, che estende la gamma di Torosaurus più a sud rispetto a quella di Triceratops , potrebbe rappresentare un genere di chasmosaurino separato o una terza specie di Triceratops . Lo studio morfometrico del 2013 di Farke era inconcludente su questo punto, e il morfospazio di T. utahensis si colloca tra il Triceratops e Torosaurus latus non separandosi abbastanza da entrambi. [10]
Note
- ^ *Hicks, JF, Johnson, KR, Obradovich, JD, Miggins, DP, and Tauxe, L. 2003. Magnetostratigraphyof Upper Cretaceous (Maastrichtian) to lower Eocene strata of the Denver Basin, Colorado. In KR Johnson, RG Raynolds and ML Reynolds (eds), Paleontology and Stratigraphy of Laramide Strata in the Denver Basin, Pt. II., Rocky Mountain Geology 38: 1-27.
- ^ Holtz, Thomas R. Jr. (2011) Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages, Winter 2010 Appendix.
- ^ Paul, GS (2010) The Princeton Field Guide to Dinosaurs , Princeton University Press.
- ^ a b c d e Longrich, NR e Field, DJ,Torosaurus is not Triceratops : Ontogeny in chasmosaurine ceratopsids as a case study in dinosaur taxonomy , in PLoS ONE , vol. 7, n. 2, 2012, p. e32623, Bibcode : 2012PLoSO...732623L , DOI : 10.1371/journal.pone.0032623 , PMC 3290593 , PMID 22393425 .
- ^ Morph-osaurs: How shape-shifting dinosaurs deceived us – life – 28 July 2010 , su newscientist.com , New Scientist, DOI : 10.1080/02724634.2010.483632 . URL consultato il 3 agosto 2010 .
- ^ a b c d e Scannella, J. and Horner, JR (2010). " Torosaurus Marsh, 1891, is Triceratops Marsh, 1889 (Ceratopsidae: Chasmosaurinae): synonymy through ontogeny ." Journal of Vertebrate Paleontology , 30 (4): 1157–1168. DOI : 10.1080/02724634.2010.483632
- ^ Brian Switek, New Study Says Torosaurus=Triceratops , in Dinosaur Tracking , Smithsonian.com. URL consultato il 2 marzo 2011 .
- ^ Horner, Jack. TEDX Talks: " Shape-shifting Dinosaurs ". Nov 2011. Accessed 20 Nov 2012.
- ^ a b Farke, AA (2011) " Anatomy and taxonomic status of the chasmosaurine ceratopsid Nedoceratops hatcheri from the Upper Cretaceous Lance Formation of Wyoming, USA. " PLoS ONE 6 (1): e16196. DOI : 10.1371/journal.pone.0016196
- ^ a b c Maiorino L., Farke AA, Kotsakis T., Piras P.,2013, "Is Torosaurus Triceratops ? Geometric Morphometric Evidence of Late Maastrichtian Ceratopsid Dinosaurs", PLoS ONE 8 (11): e81608. doi:10.1371/journal.pone.0081608
- ^ Hatcher, JB, Marsh OC and Lull, RS, 1907, The Ceratopsia , Monographs of the United States Geological Survey 49 : 1-198
- ^ Lull, RS, 1933, A revision of the Ceratopsia or horned dinosaurs , Memoirs of the Peabody Museum of Natural History 3 (3): 1-175
- ^ Lehman, TM, 1998, "A gigantic skull and skeleton of the horned dinosaur Pentaceratops sternbergi from New Mexico: Journal of Paleontology, 72 (5): 894-906
- ^ Nicholas R. Longrich, 2011, " Titanoceratops ouranous , a giant horned dinosaur from the Late Campanian of New Mexico", Cretaceous Research , 32 : 264-276
- ^ a b c d e Farke, AA "Cranial osteology and phylogenetic relationships of the chasmosaurine ceratopsid Torosaurus latus ", pp. 235-257. In K. Carpenter (ed.). Horns and Beaks: Ceratopsian and Ornithopod Dinosaurs . Indiana Univ. Press (Bloomington), 2006.
- ^ a b c Hunt, Rebecca K. and Thomas M. Lehman. 2008. Attributes of the ceratopsian dinosaur Torosaurus , and new material from the Javelina Formation (Maastrichtian) of Texas. Journal of Paleontology 82(6): 1127–1138.
- ^ a b OC Marsh, 1891, "Notice of new vertebrate fossils", The American Journal of Science, series 3 42 : 265-269
- ^ Scott D. Sampson, Mark A. Loewen, Andrew A. Farke, Eric M. Roberts, Catherine A. Forster, Joshua A. Smith e Alan A. Titus, New Horned Dinosaurs from Utah Provide Evidence for Intracontinental Dinosaur Endimism , in PLoS ONE , 5, vol. 5, n. 9, 2010, pp. e12292, Bibcode : 2010PLoSO...512292S , DOI : 10.1371/journal.pone.0012292 , PMC 2929175 , PMID 20877459 .
- ^ NR Longrich, The horned dinosaurs Pentaceratops and Kosmoceratops from the upper Campanian of Alberta and implications for dinosaur biogeography , in Cretaceous Research , vol. 51, 2014, pp. 292–308, DOI : 10.1016/j.cretres.2014.06.011 .
- ^ Dodson, P. The Horned Dinosaurs . Princeton Univ. Press (Princeton), 1996.
- ^ Colbert, EH and JD Bump, 1947, "A skull of Torosaurus from South Dakota and a revision of the genus:, Proceedings of the Academy of Natural Sciences of Philadelphia , 99 : 93–106
- ^ Farke, A., 2002, "A review of Torosaurus (Dinosauria: Ceratopsidae) specimens from Texas and New Mexico", Journal of Vertebrate Paleontology , 22 : 52A
- ^ Lucas, SG, Mack, GH, Estep, GW, 1998, "The Ceratopsian dinosaur Torosaurus from the Upper Cretaceous McRae Formation, Sierra County, New Mexico", New Mexico Geological Society Guidebook, 49th Field Conference, Las Cruces County II
- ^ CW Gilmore, 1946, "Reptilian fauna of the North Horn Formation of central Utah", United States Department of the Interior Geological Survey Professional Paper 210 -C: 29-53
- ^ DA Lawson, 1976, " Tyrannosaurus and Torosaurus , Maestrichtian dinosaurs from Trans-Pecos, Texas", Journal of Paleontology 50 (1): 158-164
- ^ Sullivan, RM, AC Boere, and SG Lucas. 2005. Redescription of the ceratopsid dinosaur Torosaurus utahensis (Gilmore, 1946) and a revision of the genus. Journal of Paleontology 79:564-582.
- ^ Scannella J., 2009, "And then there was one: synonymy consequences of Triceratops cranial ontogeny", Journal of Vertebrate Paleontology 29 : 177A
- ^ New Analyses Of Dinosaur Growth May Wipe Out One-third Of Species , su sciencedaily.com . URL consultato il 3 agosto 2010 .
- ^ John R Horner e Mark B Goodwin,Extreme Cranial Ontogeny in the Upper Cretaceous Dinosaur Pachycephalosaurus , in PLoS ONE , vol. 4, n. 10, 2009, pp. e7626, Bibcode : 2009PLoSO...4.7626H , DOI : 10.1371/journal.pone.0007626 , PMC 2762616 , PMID 19859556 .
- ^ Scannella, JB & Horner, JR, 2011, "'Nedoceratops': An Example of a Transitional Morphology", PLoS ONE 6 (12): DOI:10.1371/journal.pone.0028705
- ^ Horner, JR, Lamm, ET., 2011, "Ontogeny of the parietal frill of Triceratops : a preliminary histological analysis", Comptes Rendus de l'Academie des Sciences Paris série D 10 : 439–452
Bibliografia
- John B. Scannella, John R. Horner, (2010) Torosaurus Marsh, 1891, is Triceratops Marsh, 1889 (Ceratopsidae: Chasmosaurinae): synonymy through ontogeny. Journal of Vertebrate Paleontology, Volume 30, Issue 4 July 2010, pages 1157 - 1168
Altri progetti
-
 Wikimedia Commons contiene immagini o altri file su Torosaurus latus
Wikimedia Commons contiene immagini o altri file su Torosaurus latus -
 Wikispecies contiene informazioni su Torosaurus latus
Wikispecies contiene informazioni su Torosaurus latus
Collegamenti esterni
- ( EN ) Torosaurus latus , su Enciclopedia Britannica , Encyclopædia Britannica, Inc.
- ( EN ) Torosaurus latus , su Fossilworks.org .