Theropoda
| Teropode | |
|---|---|
 | |
| Clasificare științifică | |
| Domeniu | Eukaryota |
| Regatul | Animalia |
| Phylum | Chordata |
| Superordine | Dinozauria |
| Ordin | Saurischia |
| Clade | Eusaurischia |
| Subordine | Theropoda Marsh , 1881 |
| Subgrupuri [1] | |
Teropodul (Theropoda, din greacă θηρίον care înseamnă „fiară” și πούς, ποδός care înseamnă „picior”) sunt un grup de dinozauri saurischieni . Majoritatea erau carnivore, dar anumite genuri au evoluat în erbivore, omnivore, piscivore și insectivore. Theropods apărut în Carnian (acum 231,4 milioane de ani) [2] și a inclus numai mari carnivore terestre din Jurasic inferior la sfârșitul Cretacicului . Păsările (singurul grup de teropode care au supraviețuit evenimentului KT ) au apărut în timpul Jurasicului superior și astăzi există 10.000 de specii vii.
În 2014, oamenii de știință au publicat detalii cu privire la evoluția păsărilor de la alți dinozauri teropodi. [3] [4] [5] Printre trăsăturile care leagă păsările de ceilalți dinozauri teropodi se numără forcula , sacii de aer, eclozarea ouălor și, cel puțin în celurozauri , penajul .
Paleobiologie
Dieta și dinții

Teropodele prezintă o serie de diete diferite, inclusiv insectivore , erbivore și carnivore . Abilitatea de a se hrăni cu diverse surse de hrană, altele decât carnea, se credea anterior a fi o trăsătură unică păsărilor, dar descoperirile făcute la sfârșitul secolului XX și începutul secolului XXI au arătat că existau chiar teropode primitive cu diete. [6] Stereotipul conform căruia teropodele erau în primul rând carnivore a apărut din faptul că primele exemplare descoperite la sfârșitul secolului al XIX-lea și începutul secolului al XX-lea erau echipate cu dinți ascuțiți, zimțate, unele exemplare prezentând dovezi neechivoce de comportament prădător. Exemplele includ o fosilă Compsognathus cu o șopârlă conservată în stomac și un specimen de Velociraptor conservat care luptă un Protoceratops .
Primii teropode non-aviari confirmați a fi necarnivori au fost terizinozaurii , cunoscuți odinioară sub numele de „Segnosauri”. În trecut se credea că sunt prozauropodi , dar mai târziu s-a demonstrat că erau în schimb teropodi specializați într-o dietă erbivoră. Therizinozaurii posedau abdomenuri enorme pentru digestia legumelor și capete mici cu ciocuri și dinți în formă de frunze. Studii suplimentare asupra teropodelor maniraptoriene au arătat că teropodele nu au fost singurele teropode preistorice care au abandonat carnivorismul. Anumite alte linii de maniraptorieni primitivi demonstrează adaptări la o dietă omnivoră, inclusiv troodontidele (pentru care există dovezi că au mâncat semințele), alvarezsauri , oviraptorosauri și ornitomimozauri . Anumite alte linii, cum ar fi Masiakasaurus și spinosauridele par să se fi specializat într-o dietă piscivoră. [7] [8]
Dieta teropodelor poate fi dedusă prin respectarea morfologiei dinților, [9] semnele dinților pe oasele prăzii și conținutul stomacului. Pe lângă păsările de astăzi, se știe că anumite teropode preistorice, cum ar fi Baryonyx , Lourinhanosaurus și ornitomimozaurii, folosesc gastroliti .
Cântare și penaj

Teropodele mezozoice au arătat o diversificare ridicată în forma pielii. Penele sau structurile asemănătoare penelor erau prezente în majoritatea teropodelor. În afara celurozaurilor, penele ar fi putut fi limitate la exemplare tinere, mici sau limitate la anumite părți ale animalului. Multe teropode mari aveau pielea acoperită cu solzi mici, rotunzi. La anumite specii, aceste solzi au fost intercalate cu osteoderme . Un exemplu al acestui fenomen îl găsim în Carnotaurus ceratosaurus, din care s-au găsit diverse amprente ale pielii sale. [10]
Linii mai îndepărtate de păsări aveau pene relativ scurte compuse din filamente simple. [11] Aceste filamente au fost descoperite și la terizinozauri, care posedau pene uriașe, asemănătoare unui vârf. Alte teropode cu penaj mai larg, precum dromeosauridele , au păstrat solzi doar pe picioare. Anumite specii posedau un amestec de solzi și pene, cum ar fi Scansoriopteryx , care a arătat prezența solzilor sub coadă, [12] și Juravenator ar fi putut fi în principal cu solzi, cu unele suvite împrăștiate pe diferite părți ale corpului. [13] Alte teropode erau complet acoperite de pene, cum ar fi Anchiornis troodontid, care avea și penajul la picioare. [14]
A tăia
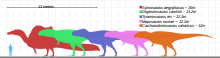
Timp de multe decenii, se credea că cel mai mare teropod era Tiranosaurul . De la descoperirea sa, s-a găsit un număr și mai mare de teropode, inclusiv Spinosaurus , Carcharodontosaurus și Giganotosaurus . [15] Exemplarele originale de Spinosaurus (precum și exemplarele găsite în 2006) susțin ideea că era cu șase metri mai lung decât Tyrannosaurus și o depășea în greutate cu o tonă. [16] În prezent, cel mai mare teropod din zilele noastre este struțul , cu o înălțime de 2,74 metri și o greutate de 63,5-145,15 kilograme. [17]
Cel mai mic teropod non-aviar a fost Anchiornis huxleyi troodontid , cântărind 110 grame și 34 cm lungime. [18] Când sunt incluse păsările, cel mai mic teropod vreodată este colibriul Helenei , cântărind 1,9 grame și 5,5 cm lungime. [19]
Este evident că teropodele au arătat o scădere a dimensiunii lor în ultimii 50 de milioane de ani, de la o medie de 163 kilograme până la 0,8 kilograme, evoluând în cele din urmă la păsări moderne. Aceasta se bazează pe faptul că teropodele au fost singurii dinozauri care au prezentat o scădere progresivă a dimensiunii și că scheletele lor s-au schimbat de patru ori mai repede decât cele ale altor dinozauri. [3] [5]
Postură
Fiind un grup foarte divers de animale, este probabil ca postura teropodelor să varieze între diferite descendențe. [20] Toate teropodele cunoscute sunt bipede, cu membrele anterioare scurte și specializate pentru diferite funcții. La păsările moderne, corpul este de obicei ținut în poziție verticală, cu femurul ținut paralel cu coloana vertebrală și forța locomotivă generată în genunchi. [20]
Chiar înainte de acceptarea conexiunii lor cu păsările, teropodele au fost înțelese a fi bipede în secolul al XIX-lea. În această perioadă, se credea că teropodele, cum ar fi carnosaurii și tiranosauridele, mergeau cu coapsele și spatele ținute în poziție verticală, iar cozile lor trecând pe pământ, într-o poziție „trepied”. [20] Această interpretare a fost pusă la îndoială în anii 1970 prin studii biomecanice. Mai mult, nici o serie de urme nu prezintă semne ale unei cozi de urcare, indicând astfel că teropodii mergeau într-o postură orizontală cu coada ținută paralel cu solul. [20] [21] Cu toate acestea, orientarea picioarelor atunci când au mers animalele rămâne controversată. Unele studii susțin ipoteza tradițională a femurului vertical (cel puțin la exemplarele mari cu cozi mai lungi), [21] în timp ce altele sugerează că genunchiul a fost în mod normal flectat în toate teropodele în timp ce mergea, chiar și la giganți precum tiranosauridele. [22] [23] Este probabil că teropodele preistorice au prezentat o mare varietate de posturi. [20] [24]
Sistem nervos
Deși rare, anumite fosile demonstrează prezența endocraniei complete. Endocrania conservată permite reconstrucția formei creierului printr-o scanare tomografică și un software 3D. Descoperirile acestui proces au o valoare evolutivă ridicată, deoarece permit cercetătorilor să documenteze apariția neurologiei moderne a păsărilor. O creștere a dimensiunii creierului pare să fi avut loc odată cu sosirea coelurozaurilor, proces care a continuat până la păsările de astăzi. [25]
Membrele anterioare

Membrele anterioare scurte proporțional cu corpul și picioarele a fost o trăsătură comună în rândul theropods, în special abelisaurids și tyrannosaurids . Cu toate acestea, această caracteristică nu era universală: spinosauridele și anumiți celurozauri aveau brațe bine dezvoltate. S-a propus că cel puțin un gen, Xuanhanosaurus , era un patruped, deoarece membrele anterioare erau relativ scurte și robuste, dar această ipoteză este acum considerată puțin probabilă. [26] Teropodii arată chiar o altă diversitate în morfologia mâinilor; cea mai comună structură consta din trei degete echipate cu gheare. Anumiți teropode timpurii (cum ar fi Herrerasaurus și Eoraptor ) aveau patru degete și un al cincilea metacarpal redus. Ceratozaurii dețineau de obicei patru degete, în timp ce majoritatea tenanurilor aveau trei. Funcția mâinilor variază în acord cu familia; este probabil ca spinosauridele să-și folosească mâinile pentru a imobiliza peștii, în timp ce se crede că anumite celurozauri mici (cum ar fi dromeosauridele și scansoriopterigidele ) le-au folosit pentru a urca în copaci. Multe păsări, care sunt teropodele de astăzi, le folosesc pentru zbor.
Contrar a ceea ce s-a arătat în arta și cinematografia populară, membrele anterioare ale teropodelor erau relativ inflexibile. [27] Se remarcă în special faptul că teropodele și alte sauriscuri bipede (inclusiv prosauropodele bipede) nu ar putea roti antebrațul într-o poziție înclinată (cu palma în jos). La om, poziția predispusă este atinsă printr-o mișcare a razei în raport cu ulna . În contrast, la dinozaurii saurisque vârful razei de la cot a fost imobilizat într-o canelură din ulna, împiedicând orice mișcare. Încheieturile au fost, de asemenea, limitate în flexibilitate, forțând antebrațul și mâna să se miște ca o singură componentă. [28]
La carnosauri precum Acrocanthosaurus , mâna în sine deținea un nivel destul de ridicat de flexibilitate, cu degetele articulate. Acest lucru se aplică și teropodelor mai primitive, cum ar fi ererasaurii și dilofosaurii. Coelurosaurii au arătat o schimbare în utilizarea antebrațului, cu o mai mare flexibilitate la umăr, permițând astfel ridicarea brațului într-o poziție orizontală. Acest lucru a atins apogeul păsărilor zburătoare moderne. La celurozauri, cum ar fi ornitomimosauridele și dromeosauridele, mâna și degetele și-au pierdut mult din flexibilitate. Dromeosauridele și alți maniraptorieni au arătat o mobilitate crescută la încheietura mâinii, datorită prezenței unui os carpian în formă de semilună, permițând astfel întregii mâini să se aplece înspre antebraț în maniera păsărilor de astăzi. [28]
Paleopatologie

În 2001, a fost publicat un sondaj privind patologiile prezente în oasele dinozaurilor teropode. Trăsături patologice au fost găsite în 21 de genuri din 10 familii. Bolile au fost descoperite la teropode de toate dimensiunile, deși erau mai puțin frecvente la teropode mici, dar acest lucru se poate datora conservării. Prezența patologiilor este mai frecventă în coaste și în vertebrele caudale. În mod ciudat, puține patologii au fost descoperite în oase care au susținut greutatea animalelor, cum ar fi sacrul, femurul și tibia. Lipsa rănilor pe aceste oase indică faptul că selecția naturală a favorizat o creștere a forței lor. Majoritatea patologiilor constau în fracturi și puncții, probabil din cauza mușcăturilor. Anumite patologii indică prezența infecției, care tindea să fie limitată la cele mai mici părți ale scheletului. S-au găsit dovezi ale defectelor congenitale, oferind astfel date utile pentru înțelegerea proceselor evolutive ale animalelor. Sinostoză sau neregularitate în elementele craniene ar putea indica vârsta înaintată a specimenului.[30]
Eu înot
Urmele unui teropod de înot au fost descoperite în formațiunea Feitianshan din Sichuan . [31] Această descoperire susține ipoteza că teropodele erau potrivite pentru înot, capabile să traverseze apă moderat adâncă. Urmele au constat din amprente de gheare, indicând astfel că animalul înota lângă țărm, atingând subgrada doar cu vârfurile ghearelor sale. Urmele arată o progresie coordonată stânga-dreapta, indicând faptul că teropodele erau înotători bine coordonați. [31]
Posibil dans de împerechere

Între 2015 și 2016, în Colorado, o echipă de cercetători și paleontologi de la Universitatea din Colorado Denver au descoperit amprente ciudate de-a lungul unui pat de roci sedimentare datate în Cretacicul inferior . [33] Aceste urme sunt semi-circulare și cu o lățime de până la doi metri și sunt dispuse paralel una cu cealaltă. La prima vedere, paleontologii s-au gândit la baza construirii unui cuib, dar cuiburile de dinozauri au o margine ridicată și un fund plat și au cel mai adesea coji de ouă . Aceste semne din sol sunt alungite și concavă și au urme de gheare , de parcă s-ar fi târât pe pământ. Aceste structuri sunt mai comparabile cu puii produși de păsările terestre îndrăgostite. În timpul leks , păsările masculine de la sol călcă pământul într-un fel de dans în timp ce își arată penele sau fac apeluri de dragoste. [34] Potrivit paleontologilor, aceasta ar fi, prin urmare, prima dovadă fosilă a unui ritual de împerechere dinozaur teropod din lume. Acest lucru sugerează că cel puțin unele grupuri mai evoluate de teropode practicau curtarea elaborată. Având în vedere perioada geologică a urmelor și zona în care au fost găsite, se crede că autorul acestor urme este un exemplu de Acrocanthosaurus . [34]
Dinozauri în chihlimbar
Chihlimbarul este bine cunoscut pentru că a păstrat în mod deosebit numeroase animale mici, în special artropode . Mai puțin cunoscut este faptul că, ocazional, chiar și vertebratele mici pot fi încorporate, chiar dacă doar parțial în chihlimbar: în special, există un număr destul de mare de anurani și șopârle , care oferă o documentare foarte rară și excepțională a formei corpului și a detaliilor anatomice ale părțile moi din aceste animale cunoscute și numai din fosile.
În iunie 2016 , Xing și colab. (2016) [ este necesară citarea ] a publicat descoperirea extraordinară a două nuclee mici de chihlimbar din Cenomanianul din Birmania . În interior erau rămășițele a două aripi mici cu pene . Penele sunt păstrate tridimensional, nu ca urme, și prezintă aranjamentul clasic al primarului și secundarului. Imaginile micro-CT arată că aripile includ și resturi scheletice ale mâinii și antebrațului. Mâna are al treilea metacarpal mai lung decât al doilea, primul deget mai scurt decât al doilea metacarpal și doar o singură falangă în al treilea deget. Această combinație indică faptul că aceste aripi aparțin enantiornithi . [35]
În decembrie același an, Xing și colab. (2016) [ este necesară citarea ] descrie descoperirea incredibilă a unui fragment de coadă de vertebrate, lung de câțiva centimetri, încorporat într-o picătură mare de chihlimbar din Cretacicul mediu din Birmania . [36] Coada păstrează tegumentul, acoperit de o serie densă de pene cu rahis și barbule . Partea scheletică a cozii este prea puțin recunoscută în masa penei, cu toate acestea, datorită scanărilor cu raze X și eliminării digitale a unei părți a penajului, sunt identificate cel puțin trei vertebre, joase și alungite, care totuși nu dau indicii pentru a fi capabil să clasifice corect animalul căruia i-a aparținut. Ca o opțiune mai conservatoare, animalul ar putea fi o archeopterygid sau jeholornithid- grad bazal Avial , sau o mai bazal paravian . Forma conică a cozii exclude atât un oviraptorosaurus . Penajul relativ complex ar tinde să excludă non-maniraptorii. Prin urmare, animalul căruia îi aparține această fosilă trebuie să fie un membru nespecific al maniraptorei . [36] [37]
Evoluţie

În timpul triasicului superior , au existat și au evoluat unul lângă altul o serie de prototeropode primitive și teropode adecvate. Cele mai vechi și primitive forme de teropode sunt reprezentate de genul Eodromaeus și de membrii familiei herrerasauridae găsite în Argentina . Herrerrasaurs a trăit în triasicul superior timpuriu (de la sfârșitul perioadei Carnic până la începutul perioadei norice ). Rămășițele lor au fost găsite atât în America de Nord, cât și în America de Sud și este posibil să fi trăit și în India și Africa de Sud. Herrerrasaurii se caracterizează printr-un set de caractere primitive și avansate. În trecut, unii paleontologi consideraseră că hererasauridele sunt membri ai cladei Theropoda, în timp ce alții speculaseră că erau sauriski bazali sau chiar forme care se diferențiaseră înainte de divergența dintre dinozaurii saurisc și ornitischian. Analizele cladice efectuate după descoperirea lui Tawa , un alt dinozaur triasic, sugerează că hererrasaurii erau teropodi primitivi. [38]
Cele mai vechi și bazale forme care pot fi clasificate cu certitudine printre teropodele reale (uneori numite „Eutheropoda”) sunt reprezentate de celofisoid , un grup de dinozauri cu corp ușor, care poate au obiceiuri sociale de viață și o distribuție largă a habitatului . Grupul include prădători mici, cum ar fi Coelophysis și forme mai mari, cum ar fi Dilophosaurus . Aceste animale au trăit să existe de la Carnianul superior (începutul Triasicului superior) până la Toarcian (sfârșitul Jurassicului inferior). Deși anterior au fost incluși printre ceratozauri , [39] este posibil să reprezinte strămoșii teropodelor ulterioare (făcând astfel grupul parafiletic ). [40] [41] Mai mulți ceratozauri derivați (cum ar fi Ceratosaurus și Carnotaurus ) au apărut în timpul Jurasicului inferior și au supraviețuit până în Jurasicul Superior în Laurasia . Au concurat cu rudele lor tetanuri (mai derivate din punct de vedere anatomic) și - prin linia evolutivă a abelisaurilor - au supraviețuit în Gondwana până la sfârșitul Cretacicului.
Tetanidele sunt mai specializate decât ceratozaurii și se împart între megalosauroizii mai bazali (sau spinosauroizi ) și aveteropodele mai derivate. Megalosauridele au trăit în principal din Jurasicul mijlociu până în Cretacicul inferior. Verii lor spinosauridici au trăit mai ales din Cretacicul inferior până în Cretacicul mediu. După cum indică denumirea lor, aveteropods reprezintă grupul care cuprinde formele cele legate de păsări, și sunt la rândul lor împărțite în allosauroids și celurosaurs (grupul care include păsările înșiși).
Coelurozaurii sunt cel mai divers grup de teropode. Printre celurozaurii Cretacici s-au numărat tiranozauride (cum ar fi Tiranosaurul ), dromeosauridele (precum Velociraptor și Deinonychus , ambele cu caracteristici similare cu cele ale Archeopteryx , cea mai veche pasăre cunoscută [42] [43] ), troodontide , oviraptorosauri , ornitomimosauri (sau " dinozauri de struț "), terizinozauri și păsări, singurul grup de dinozauri care a supraviețuit extincției în masă Cretacic-Paleocen . [44] Deși diferențierea acestor grupuri datează din Jurasicul mijlociu, abia din Cretacicul inferior a început difuzarea lor masivă. Anumiți paleontologi, cum ar fi Gregory S. Paul , au sugerat că unele sau toate formele dintre aceste teropode derivate ar fi putut proveni din dinozauri zburători sau din proto-păsări, cum ar fi Archaeopteryx, care ar fi pierdut capacitatea de a zbura pentru a reveni la un stil de viață terestru. . [45]
Clasificare și taxonomie
Această intrare sau secțiune despre subiectul dinozaurilor nu citează sursele necesare sau cei prezenți sunt insuficienți . |
Clasificare
Următoarea carte arată poziția Theropoda în Dinosauria .
- Sauropodomorpha
- Theropoda
| Dinozauria |
| ||||||||||||||||||||||||||||||||||||||||||
Taxonomie




Următoarea foaie prezintă o clasificare simplificată a principalelor clade Theropoda.
- Theropoda
| Theropoda |
| ||||||||||||||||||||||||||||||||||||||||||||||||
Cladi
Această intrare sau secțiune despre subiectul dinozaurilor nu citează sursele necesare sau cei prezenți sunt insuficienți . |
- † Herrerasauridele , probabil cele mai vechi teropode , includ forme primitive precum Staurikosaurus , Gnathovorax și Herrerasaurus .
- † Celofisoizi , nu se știe cu siguranță dacă sunt ceratozauri primitivi. Include celofiza .
- † Dilofosauride
- † Ceratosaurians , includ Saltriovenator , Ceratosaurus și Abelisauroids . Aceștia din urmă au fost principalii prădători ai emisferei sudice în Cretacicul târziu.
- Tetanuri , cel mai mare grup dintre toate, include Spinosauroids și Neotetanurs ( Carnosaurs și Coelurosaurs ).
- † Megalosauroizi , include și Spinozaurii; se poate numi Spinosauroidea sau Torvosauroidea.
- Neotetanuri , teropode avansate care seamănă cu păsările, dacă apare și Neotheropoda în clade acestea pot fi numite Avetheropods .
- † Allosauroids , include Allosaurus și prădători chinezi, cum ar fi Sinraptor și prădători uriași, cum ar fi Mapusaurus .
- Coelurosaurianii , un grup mare cu diferite forme ca mărime, dietă și habitat.
- † Tiranosaurii , nu sunt la fel de mici ca mulți celurozauri; sunt o cladă foarte mare.
- † Ornitomimozauri , femuri lungi și probabil omnivori; au mulți dinți.
- Maniraptorii , păsările și strămoșii lor cei mai apropiați, variază foarte mult în mărime și pot fi din toate dietele; toate sau majoritatea pot fi pene.
- † Terizinozauri , erbivore mari bipede cu gheare lungi pe picioarele din față; burta umflata.
- † Alvarezsauri , mici insectivore asemănătoare păsărilor cu femururi lungi.
- † Oviraptorosauri , mulți dinți; dieta și modul lor de viață sunt încă studiate.
- † Deinonicosauri , mai des numiți „Raptor” ; gheara mare de secera pe al doilea deget; creier și ochi relativ mari.
- Avialae : acest grup include păsări actuale și dispărute ( Jeholornitidi , Confuciusorniti , Scansoriopterigidi , Aves ).
- † Jeholornithidae
- † Omnivoropterygidae
- † Confuciusornithidae
- † Enantiornithes
- † Hesperornithes
- Aves , păsări tradiționale.
Notă
- ^ Holtz, Thomas R. Jr. (2012). Dinozauri: cea mai completă enciclopedie actualizată pentru iubitorii de dinozauri de toate vârstele , iarna 2011 Anexă .
- ^ Oscar A. Alcober și Martinez, Ricardo N., A new herrerasaurid (Dinosauria, Saurischia) from the Upper Triassic Ischigualasto Formation of nord-vest Argentina , in ZooKeys , vol. 63, nr. 63, 2010, pp. 55–81, DOI : 10.3897 / zookeys.63.550 , PMC 3088398 , PMID 21594020 . [1]
- ^ a b Seth Borenstein, Studiul urmărește evoluția dinozaurilor în păsările timpurii , în AP News , 31 iulie 2014. Adus pe 3 august 2014 (arhivat din original la 8 august 2014) .
- ^ Lee, MichaelS.Y.; Cau, Andrea; Naish, Darren; Dyke, Gareth J., Sustained miniaturization and anatomical innovation in the dinosaurian ancestors of birds , in Science , vol. 345, n. 6196, 1º agosto 2014, pp. 562–566, DOI : 10.1126/science.1252243 . URL consultato il 2 agosto 2014 .
- ^ a b Zoe Gough, Dinosaurs 'shrank' regularly to become birds , BBC , 31 luglio 2014.
- ^ Zanno, LE, Gillette, DD, Albright, LB, and Titus, AL (2009). "A new North American therizinosaurid and the role of herbivory in 'predatory' dinosaur evolution." Proceedings of the Royal Society B , Published online before print July 15, 2009, DOI : 10.1098/rspb.2009.1029 .
- ^ Nicholas R. Longrich e Philip J. Currie, Albertonykus borealis , a new alvarezsaur (Dinosauria: Theropoda) from the Early Maastrichtian of Alberta, Canada: Implications for the systematics and ecology of the Alvarezsauridae , in Cretaceous Research , vol. 30, n. 1, 2009, pp. 239–252, DOI : 10.1016/j.cretres.2008.07.005 .
- ^ TR Holtz, Jr, DL Brinkman e CL Chandler, Dental morphometrics and a possibly omnivorous feeding habit for the theropod dinosaur Troodon , in GAIA , vol. 15, 1998, pp. 159–166.
- ^ Hendrickx, C., & Mateus O. (2014). Abelisauridae (Dinosauria: Theropoda) from the Late Jurassic of Portugal and dentition-based phylogeny as a contribution for the identification of isolated theropod teeth. Zootaxa. 3759, 1-74.
- ^ Bonaparte, Novas, and Coria (1990). " Carnotaurus sastrei Bonaparte, the horned, lightly built carnosaur from the Middle Cretaceous of Patagonia." Contributions in Science (Natural History Museum of Los Angeles County) , 416 : 41 pp.
- ^ UB Göhlich e LM Chiappe, A new carnivorous dinosaur from the Late Jurassic Solnhofen archipelago , in Nature , vol. 440, n. 7082, 2006, pp. 329–332, DOI : 10.1038/nature04579 , PMID 16541071 .
- ^ Czerkas, SA, and Yuan, C. (2002). "An arboreal maniraptoran from northeast China." Pp. 63–95 in Czerkas, SJ (Ed.), Feathered Dinosaurs and the Origin of Flight. The Dinosaur Museum Journal 1 . The Dinosaur Museum, Blanding, USA PDF abridged version
- ^ UB Goehlich, H. Tischlinger e LM Chiappe, Juraventaor starki (Reptilia, Theropoda) ein nuer Raubdinosaurier aus dem Oberjura der Suedlichen Frankenalb (Sueddeutschland): Skelettanatomie und Wiechteilbefunde , in Archaeopteryx , vol. 24, 2006, pp. 1–26.
- ^ X. Xu, Q. Zhao, M. Norell, C. Sullivan, D. Hone, G. Erickson, X. Wang, F. Han e Y. Guo, A new feathered maniraptoran dinosaur fossil that fills a morphological gap in avian origin , in Chinese Science Bulletin , vol. 54, n. 3, 2009, pp. 430–435, DOI : 10.1007/s11434-009-0009-6 . Abstract
- ^ F. Therrien e DM Henderson, [108:MTIBTY 2.0.CO;2 My theropod is bigger than yours...or not: estimating body size from skull length in theropods ], in Journal of Vertebrate Paleontology , vol. 27, n. 1, 2007, pp. 108–115, DOI : 10.1671/0272-4634(2007)27[108:MTIBTY]2.0.CO;2 .
- ^ C. dal Sasso, S. Maganuco, E. Buffetaut e MA Mendez, New information on the skull of the enigmatic theropod Spinosaurus , with remarks on its sizes and affinities , in Journal of Vertebrate Paleontology , vol. 25, n. 4, 2005, pp. 888–896, DOI : 10.1671/0272-4634(2005)025[0888:NIOTSO]2.0.CO;2 .
- ^ Ostrich | African Wildlife Foundation
- ^ Xu, X., Zhao, Q., Norell, M., Sullivan, C., Hone, D., Erickson, G., Wang, X., Han, F. and Guo, Y. (2009). "A new feathered maniraptoran dinosaur fossil that fills a morphological gap in avian origin." Chinese Science Bulletin , 6 pages, accepted November 15, 2008.
- ^ Conservation International (Content Partner); Mark McGinley (Topic Editor). 2008. "Biological diversity in the Caribbean Islands." In: Encyclopedia of Earth. Eds. Cutler J. Cleveland (Washington, DC: Environmental Information Coalition, National Council for Science and the Environment). [First published in the Encyclopedia of Earth May 3, 2007; Last revised August 22, 2008; Retrieved November 9, 2009]. < http://www.eoearth.org/article/Biological_diversity_in_the_Caribbean_Islands >
- ^ a b c d e Hutchinson, JR, The evolution of locomotion in archosaurs , in Comptes Rendus Palevol , vol. 5, 3–4, 2006, pp. 519–530, DOI : 10.1016/j.crpv.2005.09.002 .
- ^ a b BH Newman,Stance and gait in the flesh-eating Tyrannosaurus , in Biological Journal of the Linnean Society , vol. 2, n. 2, 1970, pp. 119–123, DOI : 10.1111/j.1095-8312.1970.tb01707.x .
- ^ K. Padian, PE Olsen, (1989). "Ratite footprints and the stance and gait of Mesozoic theropods." Pp. 231–241 in: DD Gillette, MG Lockley (Eds.), Dinosaur Tracks and Traces , Cambridge University Press, Cambridge.
- ^ GS Paul, Limb design, function and running performance in ostrich-mimics and tyrannosaurs , in Gaia , vol. 15, 1998, pp. 257–270.
- ^ JO Farlow, SM Gatesy, Jr. Holtz, JR Hutchinson e JM Robinson, Theropod locomotion , in Am. Zool. , vol. 40, 2000, pp. 640–663, DOI : 10.1093/icb/40.4.640 .
- ^ "Abstract", in Chure (2001). Pg. 19.
- ^ Rauhut, OW (2003). The Interrelationships and Evolution of Basal Theropod Dinosaurs . Blackwell Publishing, 213 pp. ISBN 0-901702-79-X
- ^ K. Carpenter, Forelimb biomechanics of nonavian theropod dinosaurs in predation , in Senckenbergiana Lethaea , vol. 82, n. 1, 2002, pp. 59–76, DOI : 10.1007/BF03043773 .
- ^ a b P. Senter e JH Robins, Range of motion in the forelimb of the theropod dinosaur Acrocanthosaurus atokensis , and implications for predatory behaviour , in Journal of Zoology, London , vol. 266, n. 3, 2005, pp. 307–318, DOI : 10.1017/S0952836905006989 .
- ^ Senter P, Juengst SL (2016) Record Breaking Pain: The Largest Number and Variety of Forelimb Bone Maladies in a Theropod Dinosaur. PLoS ONE 11(2): 0149140. doi:10.1371/journal.pone.0149140
- ^ Molnar, RE, 2001, Theropod paleopathology: a literature survey: In: Mesozoic Vertebrate Life, edited by Tanke, DH, and Carpenter, K., Indiana University Press, p. 337-363.
- ^ a b Xing LD, Lockley MG, Zhang JP, et al. A new Early Cretaceous dinosaur track assemblage and the first definite non-avian theropod swim trackway from China. Chin Sci Bull, DOI : 10.1007/s11434-013-5802-6
- ^ Martin G. Lockley, Richard T. McCrea, Lisa G. Buckley, Jong Deock Lim, Neffra A. Matthews, Brent H. Breithaupt, Karen J. Houck, Gerard D. Gierliński, Dawid Surmik, Kyung Soo Kim, Lida Xing, Dal Yong Kong, Ken Cart, Jason Martin & Glade Hadden, Theropod courtship: large scale physical evidence of display arenas and avian-like scrape ceremony behaviour by Cretaceous dinosaurs Scientific Reports 6, Article number: 18952
- ^ Copia archiviata , su thenewsindependent.com . URL consultato il 27 gennaio 2016 (archiviato dall' url originale il 3 febbraio 2016) .
- ^ a b https://theconversation.com/mysterious-footprint-fossils-point-to-dancing-dinosaur-mating-ritual-52911
- ^ https://theropoda.blogspot.it/2016/06/i-primi-dinosauri-mesozoici.html
- ^ a b https://theropoda.blogspot.it/2016/12/una-coda-di-dinosauro-conservata.html
- ^ https://theropoda.blogspot.it/2016/12/un-test-sulla-posizione-filogenetica.html
- ^ SJ Nesbitt, ND Smith, RB Irmis, AH Turner, A. Downs e MA Norell, A complete skeleton of a Late Triassic saurischian and the early evolution of dinosaurs , in Science , vol. 326, n. 5959, 2009, pp. 1530–1533, DOI : 10.1126/science.1180350 , PMID 20007898 . .
- ^ Rowe, T., and Gauthier, J., (1990). "Ceratosauria." Pp. 151–168 in Weishampel, DB, Dodson, P., and Osmólska, H. (eds.), The Dinosauria , University of California Press, Berkeley, Los Angeles, Oxford.
- ^ Mortimer, M. (2001). " Rauhut's Thesis ", Dinosaur Mailing List Archives, 4 Jul 2001.
- ^ MT Carrano, SD Sampson e CA Forster, [0510:TOOMKA 2.0.CO;2 The osteology of Masiakasaurus knopfleri , a small abelisauroid (Dinosauria: Theropoda) from the Late Cretaceous of Madagascar ], in Journal of Vertebrate Paleontology , vol. 22, n. 3, 2002, pp. 510–534, DOI : 10.1671/0272-4634(2002)022[0510:TOOMKA]2.0.CO;2 .
- ^ JH Ostrom, Osteology of Deinonychus antirrhopus , an unusual theropod from the Lower Cretaceous of Montana , in Peabody Museum Natural History Bulletin , vol. 30, 1969, pp. 1–165.
- ^ Paul, GS (1988). Predatory Dinosaurs of the World . New York: Simon and Schuster Co. ( ISBN 0-671-61946-2 )
- ^ Dingus, L. and Rowe, T. (1998). The Mistaken Extinction: Dinosaur Evolution and the Origin of Birds . Freeman.
- ^ Paul, GS (2002). Dinosaurs of the Air: The Evolution and Loss of Flight in Dinosaurs and Birds. Baltimore: Johns Hopkins University Press. 472 pp. ( ISBN 0-8018-6763-0 )
- ^ Anderson, Ted R. (2006). Biology of the Ubiquitous House Sparrow: from Genes to Populations. Oxford: Oxford University Press. ISBN 0-19-530411-X .
Bibliografia
- Carrano, MT, Sampson, SD & Forster, CA, (2002), The osteology of Masiakasaurus knopfleri , a small abelisauroid (Dinosauria: Theropoda) from the Late Cretaceous of Madagascar. Journal of Vertebrate Paleontology Vol. 22, #3, pp. 510–534
- Dingus, L. & Rowe, T. (1998), The Mistaken Extinction: Dinosaur Evolution and the Origin of Birds , Freeman
- Kirkland , JI, Zanno, LE, Sampson, SD, Clark, JM & DeBlieux, DD, (2005) A primitive therizinosauroid dinosaurs from the Early Cretaceous of Utah, Nature: Vol. 435, pp. 84–87
- Mortimer, M., (2001) " Rauhut's Thesis ", Dinosaur Mailing List Archives, 4 Jul 2001
- Ostrom, JH (1969). Osteology of Deinonychus antirrhopus, an unusual theropod from the Lower Cretaceous of Montana, Peabody Museum Nat. History Bull. , 30, 1-165
- Paul, GS, (1988) Predatory Dinosaurs of the World Simon and Schuster Co., New York ( ISBN 0-671-61946-2 )
- ----- (2002) Dinosaurs of the Air ( ISBN 0-8018-6763-0 ):
- Rowe, T., & Gauthier, J., (1990) Ceratosauria. 151-168 in Weishampel, DB, Dodson, P., & Osmólska, H. (eds.), The Dinosauria , University of California Press, Berkley, Los Angeles, Oxford.
Altri progetti
-
 Wikimedia Commons contiene immagini o altri file su Theropoda
Wikimedia Commons contiene immagini o altri file su Theropoda -
 Wikispecies contiene informazioni su Theropoda
Wikispecies contiene informazioni su Theropoda
Collegamenti esterni
- ( EN ) Theropoda , su Enciclopedia Britannica , Encyclopædia Britannica, Inc.
- ( EN ) Theropoda , su Fossilworks.org .
| Controllo di autorità | GND ( DE ) 4548235-4 |
|---|