Coelophysis bauri
| Celofiza | |
|---|---|
 | |
| Starea de conservare | |
Fosil | |
| Clasificare științifică | |
| Domeniu | Eukaryota |
| Regatul | Animalia |
| Phylum | Chordata |
| Superordine | Dinozauria |
| Ordin | Saurischia |
| Subordine | Theropoda |
| Familie | † Coelophysidae |
| Tip | † Celofiza Cope , 1889 |
| Nomenclatura binominala | |
| † Coelophysis bauri Cope, 1889 | |
| Sinonime | |
| Specii | |
| |
Coelophysis (al cărui nume înseamnă „formă goală la interior“ , cu referire la forma gol a vertebrelor sale [1] ) este un gen pe cale de disparitie dinozaur theropod coelophyside trăit între Triasic superior și jurasic inferior , aproximativ 212 milioane la 196 de milioane de ani ( Norian - Sinemurian ) [2] , în ceea ce este acum sud-vestul Statelor Unite [3] , Africa de Sud și Zimbabwe . [4] Deși numărul speciilor atribuite genului este destul de variabil în funcție de autor, cele mai recente studii indică faptul că genul conține o singură specie [5] , C. bauri, atribuită inițial genului Coelurus de Edward Drinker Cope , în 1887 și descrise de acesta din urmă în 1889. numele Longosaurus și Rioarribasaurus sunt sinonime ale Coelophysis. De asemenea, un alt tip de celofilă , Megapnosaurus , este considerat un sinonim al genului, deși studii mai recente au re-autentificat validitatea. [5]
Coelofiza era un mic biped carnivor , ușor de construit, care putea atinge o lungime maximă de 3 metri (9,8 picioare). Este unul dintre cele mai vechi genuri de dinozauri cunoscuți, ale căror rămășițe fosile au fost găsite în diferite părți ale lumii, în formațiunile triasice târzii și jurasice timpurii. Este, de asemenea, unul dintre cele mai cunoscute genuri de dinozauri, datorită abundenței exemplarelor fosile.
Descriere

Coelofiza este unul dintre genurile de dinozauri mai bine cunoscute prin abundența grației exemplarelor fosile aparținând speciei C. bauri, dezvăluind că era un dinozaur de constituție ușoară care putea ajunge până la 3 metri (9,8 picioare) în lungime [6] , pentru o înălțimea până la laturile a puțin mai mult de un metru. [6] Paul (1988) a estimat că greutatea formei gracile este de 15 kg și greutatea formei solide la 20 kg (44 lbs) [7] , ceea ce ar indica un animal ușor, agil și rapid. [8] În ciuda faptului că este unul dintre cei mai vechi dinozauri cunoscuți, morfologia sa era deja foarte avansată în comparație cu creaturi precum Herrerasaurus și Eoraptor . Bustul este conforma Coelophysis la forma de baza a theropod corpului, în timp ce centura scapulară prezintă unele caracteristici distinctive: C. bauri a avut un forcula , prezent în toate theropods au evoluat și la păsări, iar cel mai vechi cunoscut un astfel de dinozaur veche. Coelofiza a păstrat, de asemenea, condiția ancestrală de a deține patru degete pe mână, deși doar trei erau operaționale, în timp ce a patra a fost probabil încorporată în țesuturile mâinii. [9]
Coelofiza avea șolduri înguste, membrele anterioare adaptate pentru apucarea prăzii și picioarele înguste. [10] Chiar și pelvisul și membrele posterioare sunt ușoare variații ale planului corpului de bază al teropodelor, prezentând o cupă membrele deschise și o dreaptă sub corp, caracteristici care definesc Dinosauria . Membrul posterior s-a terminat cu un picior cu trei degete, un deget ridicat care a servit drept pinten. Gâtul și coada erau lungi și subțiri [3], iar acesta din urmă avea o structură neobișnuită în prezigapofisi a vertebrelor caudale, care formau o rețea semi-rigidă, dând mai multă rigiditate cozii și limitând mișcarea verticală. [11]
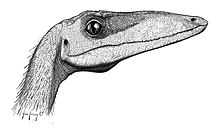
Animalul avea un cap lung și îngust (aproximativ 270 mm), cu ochi mari orientați spre înainte, care îi confereau o vedere stereoscopică și, prin urmare, o percepție excelentă a adâncimii. Rinehart și colab. (2004) au descris inelul sclerotic complet găsit la un tânăr C. bauri (specimenul NMMNH P-4200) și l-au comparat cu datele referitoare la inelele sclerotice ale reptilelor și păsărilor și au concluzionat că Coelofiza este un prădător diurn , care a folosit în principal vedere la vânătoare. [12] Studiul a relevat, de asemenea, că viziunea Coelophysis era superioară viziunii majorității șopârlelor și era comparabilă cu cea a păsărilor de pradă moderne, cu o putere acomodativă ridicată. Datele sugerează, de asemenea, o viziune nocturnă slabă, ceea ce ar însemna că acest dinozaur avea o pupilă mai degrabă rotundă decât o înclinată. [8]
Botul era alungit și echipat cu ferestre mari care ajutau la reducerea greutății craniului, în timp ce tijele înguste de os păstrau integritatea structurală a craniului. Gâtul avea o curbă sigmoidă pronunțată. Deși aveți craniu complet în C. bauri, s-ar putea obține puține dovezi, deoarece craniul multor indivizi este zdrobit. [10] Spre deosebire de alte teropode, ornamentele craniene ale Coelophysis nu erau situate în partea superioară a craniului, ci pe marginea dorsolaterală a oaselor nazale și lacrimale din craniu, unde erau prezente creste osoase scăzute, ridicate lateral, direct deasupra antorbitalului. fenestra. [13]

Un diagnostic este o declarație a trăsăturilor anatomice ale unui organism (sau grup) care îl disting în mod colectiv de toate celelalte organisme. Unele, dar nu toate, caracteristicile unui diagnostic sunt și autapomorfii. Autapomorfia este o caracteristică anatomică distinctă, care este unică pentru o anumită organizație sau grup. Conform Ezcurra (2007) și Bristowe și Raath (2004), celofiza se poate distinge în funcție de următoarele caracteristici: absența unui proces rostral decalat al maxilarului; pătratul este puternic caudal; o mică fereastră mandibulară externă, care reprezintă 9-10% din lungimea mandibulară; [14] iar lungimea anteroposterioră a procesului lacrimal ventral este mai mare de 30% din înălțimea sa. [15]
Mai mulți paleontologi consideră că Coelophysis bauri este același dinozaur ca și Coelophysis rhodesiensis (anterior Syntarsus sau, alternativ, Megapnosaurus ), însă acest lucru a fost infirmat de următoarele: Downs (2000) a concluzionat că C. bauri diferă de C. rhodesiensis în lungimea colului uterin, proporțiile proximale și distale ale membrelor posterioare și anatomia vertebrală caudală proximală; [16] Tykoski și Rowe (2004) au ajuns la concluzia că C. bauri diferă de C. rhodesiensis deoarece nu are o groapă la baza procesului nazal al premaxilei ; [10] și Bristowe și Raath (2004) au ajuns la concluzia că C. bauri diferă de C. rhodesiensis prin faptul că are un rând de dinți maxilari cei mai lungi. [15]
Clasificare

Coelofiza este un tip taxonomic compus posibil din două specii: specia tip C. bauri și posibila specie C. rhodesiensis (aceasta din urmă poate reprezenta de fapt genul Megapnosaurus ). [5] [17] Alte două specii au fost descrise de Cope, C. longicollis și C. willistoni, dar nu sunt considerate valide și sunt considerate în prezent sinonime pentru C. bauri. Specia C. rhodesiensis, provenind din Jurasicul inferior al Zimbabwe , face probabil parte din acest complex generic. [17] O a treia specie posibilă este Coelophysis kayentakatae, denumită anterior genul Megapnosaurus. [18] [19] Cu toate acestea, opiniile cu privire la numărul și validitatea speciilor de celofiză variază de la autor la autor.
La începutul anilor 1990 a avut loc o dezbatere cu privire la caracteristicile de diagnostic ale primelor exemplare găsite în comparație cu exemplarele din cariera Ghost Ranch. Unii paleontologi au fost de părere că exemplarele originale nu erau diagnostice și, prin urmare, că numele C. bauri nu ar putea fi aplicat niciunui exemplu suplimentar. Prin urmare, exemplarele din Ghost Ranch au fost numite Rioarribasaurus. [20] Cu toate acestea, deoarece specimenele Ghost Ranch erau acum cunoscute sub numele de Coelophysis în cea mai mare parte a literaturii științifice, ar fi fost incomod pentru cercetători să folosească numele Rioarribasaurus , astfel încât s-a depus o petiție pentru a transfera specimenul de tip Coelophysis din fragmentul original. exemplar la unul dintre exemplarele cele mai bine conservate din Ghost Ranch. Acest lucru l-ar face permanent Rioarribasaurus un sinonim al celofizei, în special un sinonim obiectiv junior. [21] În cele din urmă, Comisia Internațională pentru Nomenclatura Zoologică (ICZN) a adoptat această moțiune, a scăpat cu totul de numele Rioarribasaurus , declarându-l un nomen refuseum , sau "nume respins", în timp ce Coelophysis a fost ales un nomen conservandum ("nume păstrat" ). [22]
Într-o situație care afectează mulți taxoni de dinozauri, fosilele descoperite ulterior au fost inițial clasificate ca noi genuri, deși în realitate ar putea fi o specie de Coelophysis. De exemplu, descoperirea în 1911 a prof. Mignon Talbot, [23] numită Podokesaurus Holyokensis, a fost considerată mult timp ca fiind legată de Coelophysis [24] , iar unii oameni de știință moderni consideră Podokesaurus sinonim cu Coelophysis. [17] Un alt exemplu al formațiunii Portland din bazinul Hartford, aflat acum la Boston Museum of Science, a fost raportat în Coelophysis. [25] [26] Acest specimen constă din turnări în gresie ale unui pubis, o tibie, trei coaste și o cutie vertebrală și probabil provine dintr-o carieră din Middletown, Connecticut. Cu toate acestea, atât tipul de specimen Podokesaurus , cât specimenul din Middletown sunt considerate în general teropode nedeterminate. [27]
În 1999 , Sullivan și Lucas (1999) au sugerat că specimenul de tip original (AMNH 2706) a fost atribuit a ceea ce se credea a fi un celofisoid abia descoperit, eucoelophysis baldwini . [28] Cu toate acestea, studii suplimentare au arătat că eucoelofiza este de fapt o bază de referință ornitodiro , poate chiar un dinozaur, legată de enigmaticul Silesaurus . [29]
Genul a fost numit de Syntarsus Raath, în 1969, specia tip Syntarsus rhodesiensis Africa și ulterior aplicat rămășițelor nord-americane, numit Syntarsus kayentakatae . [30] Genul a fost apoi redenumit de entomologul american Dr. Michael Ivie ( Universitatea de Stat din Montana din Bozeman ), de dr. Australian polonez Adam Ślipiński și de dr. Polonez Piotr Węgrzynowicz (Muzeum Ewolucji Instytutu Zoologii PAN din Varșovia ), cei trei oameni de știință care au descoperit că numele genului Syntarsus fusese deja folosit pentru un gândac colydiine descris în 1869. [31] Mulți paleontologi nu au apreciat numele Megapnosaurus , în parte, deoarece taxonomiștii sunt, în general, așteptați ca autorii originali ai unui nume să corecteze orice erori. în munca lor. Raath era conștient de omonimie printre dinozaurii Syntarsus și Syndarsus gândac, dar grupul care a numit Megapnosaurus a fost condus să creadă că Raath a murit și, prin urmare, nu a putut să-și corecteze greșeala și a procedat în consecință. Mortimer (2012) a subliniat că „paleontologii ar fi putut reacționa mai pozitiv dacă numele substitut (Megapnosaurus) nu ar fi fost atât de ironic, rezultând, de fapt, în„ șopârlă mare moartă ”. [32] Yates (2005) a analizat Coelophysis și Megapnosaurus , concluzionând că cele două genuri sunt aproape identice, și a sugerat că Megapnosaurus era , probabil , un sinonim al Coelophysis. [33] în 2004, Raath a fost co-autor a două articole în care susținea că Megapnosaurus (fostă Syntarsus) a fost un sinonim junior Coelophysis . [17] Megapnosaurus a fost considerat de Paul (1988) și Downs (2000) ca fiind congenerico cu Coelophysis. [7] [16] apoi, în 1993, Paul a sugerat că Coelophysis ar trebui plasat în Megapnosaurus (cunoscut pe atunci sub numele de Syntarsus) obține în jurul valorii de confuzie menționată mai sus taxonomia . [34] Downs (2000) a examinat Camposaurus și a concluzionat că a fost un sinonim junior al Coelophysis, din cauza asemănării sale cu unele exemplare de Coe lophysis of Ghost Ranch. [16] Cu toate acestea, o reevaluare a holotype Camposaurus Martin Ezcurra și Stephen Brusatte publicat în 2011 a arătat o pereche de autapomorfie nell'olotipo, indicând faptul că C. arizonensis nu era sinonim cu C. bauri, deși era o rudă apropiată de C. rodesensis . [35]
Mai jos este o cladogramă pe primele teropode, bazată pe analiza efectuată de filogenetic Sues și colab. (2011): [36]
| Theropoda |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
În 2020, Marsh și Rowe, în analiza lor, decid să păstreze denumirea generică Syntarsus atât pentru exemplarele QG 1 cât și pentru MNA V2623, precum și pentru exemplarele respective atribuite acestor taxoni, spre deosebire de Coelophysis sau Megapnosaurus , datorită relațiilor sistematice cu interiorul Coelophysoidea în flux. Prin urmare, congenerozitatea sau nevoia de Megapnosaurus nu ar fi susținută dacă Coelophysis bauri, Syntarsus rhodesiensis și Syntarsus kayentakatae nu ar forma clade respective, după cum reiese din analiza lor filogenetică. [37]
Ezcurra și colab. (2021) arată că Megapnosaurus rhodesiensis era destul de îndepărtat atât de Coelophysis bauri (acum doar specii din genul Coelophysis), cât și de „Syntarsus” kayentakatae (care nu sunt clasate în prezent într-un fel bun). În această analiză, cele mai apropiate rude ale lui M. rhodesiensis sunt Camposaurus , segisaurus și Lucianovenator . [5]
Sinonime
Sinonimele celofizei includ de obicei:
- Podokesaurus ?
Talbot, în 1911 [17] - Longosaurus
Welles, 1984 [38] - Rioarribasaurus
Hunt & Lucas, 1991 [38] - Gojirasaurus ?
Tâmplar, 1997 [39] - Camposaurus ?
Hunt și colab., 1998 [40] - Megapnozaur ?
Ivie, Ślipiński și Węgrzynowicz, 2001 [17]
Sinonimele speciei C. bauri, cu toate acestea, includ:
- Coelurus bauri
Cope, 1887 - Tanystropheus bauri
(Cope, 1887) - Coelurus longicollis
Cope, 1887 - Tanystropheus longicollis
(Cope, 1887) - Coelophysis longicollis
(Cope, 1887) - Longosaurus longicollis
(Cope, 1887) - Tanystropheus willistoni
Cope, 1887 - Coelophysis willistoni
(Cope, 1887) - Rioarribasaurus colberti
Hunt & Lucas, 1991
Istoria descoperirii

Numită în 1887, specia tip de Coelophysis a fost inițial numită ca un fel de Coelurus . [21] Edward Drinker Cope a numit Coelophysis pentru prima dată în 1889 pentru a numi un nou gen, în afara lui Coelurus și Tanystropheus, pe care C. bauri fusese clasificat anterior pentru bauri C., C. și C. willistoni longicollis. [41] Un colecționar de amatori de fosile care lucra pentru Cope, David Baldwin, găsise primele rămășițe ale dinozaurului în 1881 în Chinle Formation din nord-vestul New Mexico. [42] La începutul anului 1887, exemplarele Cope affibiò colectate în două specii noi, C. și C. bauri longicollis de gen Coelurus . Mai târziu, în 1887, materialul Cope a fost repartizat unui alt tip, Tanystropheus. Doi ani mai târziu, Cope și-a corectat clasificarea după ce a observat diferențele dintre vertebre și a numit genul Coelophysis , cu C. bauri ca specie tip, [41] care a fost numit, bauri , de la Georg Baur, un anatomist comparativ ale cărui idei erau similare. la Cope's. [42] [43] Numele derivă din grecescul Coelophysis κοῖλος / koilos (care înseamnă „cablu”) și φύσις / physis (care înseamnă „formă”), apoi „formă goală” o referință la carierele vertebrelor animalului. [41] [44] Cu toate acestea, aceste prime descoperiri au fost prea puțin conservate pentru a oferi o imagine completă a acestui nou dinozaur.

În 1947, o expediție în New Mexico , condusă de Edwin H. Colbert , a făcut o descoperire senzațională. Un „cimitir” substanțial de fosile de Coelophysis a fost găsit de George Whitaker, asistentul lui Colbert, pe o proprietate cunoscută sub numele de Ghost Ranch , lângă descoperirea originală, unde s-au găsit sute de schelete de Coelophysis de diferite vârste și dimensiuni. . Colbert a efectuat un studiu cuprinzător [13] al tuturor fosilelor găsite până la acea dată și le-a atribuit coelofizei. Exemplarele Ghost Ranch au fost atât de numeroase, inclusiv multe exemplare bine conservate și complet articulate, încât unul dintre ele a devenit de atunci „ exemplarul de tip , pentru întregul gen, înlocuind exemplarul original, slab conservat. [22]
„Syntarsus” rhodesiensis a fost descris pentru prima dată de Raath (1969) și atribuit Podokesauridae . [30] Taxonul „Podokesauridae” a fost abandonat deoarece tipul său a fost distrus într-un incendiu și nu mai poate fi comparat cu noile descoperiri. De-a lungul anilor, paleontologii au atribuit astfel de ceratosauridae (Welles, 1984), Procompsognathidae (Parrish și Carpenter, 1986) și ceratosauria (Gauthier, 1986). În 2004, „Syntarsus” a fost recunoscut ca sinonim cu Coelophysis de la Tykoski și Rowe (2004). Ulterior, Ezcurra și Novas (2007) și Ezcurra (2007) au concluzionat că „Syntarsus” era sinonim cu Coelophysis, care este consensul științific actual. [45]
Paleobiologie
Dietă

Dinții Coelofizei au avut forma tipică prezentă la mulți dinozauri prădători: similar cu lamele, curbate, ascuțite și zimțate cu zimțări fine atât pe marginea din față, cât și pe cea din spate. Dinții lui indică o dietă carnivoră , compusă probabil din animale mici, precum insecte , mamifere și șopârle , ale căror rămășițe au fost găsite în aceleași locuri. [46] De asemenea, este posibil ca aceste animale să vâneze în haite pentru a doborî pradă mai mare. [7] C. bauri prezintă aproximativ 26 de dinți pe maxilar și 27 de dinți pe maxilarul inferior. [13] Carpenter (2002) a examinat biomecanica membrelor anterioare ale diferitelor teropode, având tendința de a evalua utilitatea sa în prădare. În studiul său, el a concluzionat că partea din față a membrelor Coelophysis era flexibilă și avea o bună libertate de mișcare, dar structura lor osoasă sugerează că erau relativ slabe. Caracteristici precum picioarele anterioare „slabe” și dinții mici par să sugereze că Coelophysis predase în primul rând animalele mai mici. Rinehart și colab. Coelofiza a fost de acord că un „vânător agil de pradă mică”. [8] Carpenter a identificat chiar și trei modele distincte care utilizează membrele anterioare ale teropodelor și a observat că Coelophysis face parte din modelul „gripper”, comparativ cu alți dinozauri care se aflau în cadrul modelelor „grip ferm” sau „gripper cu brațe lungi”. [47]

De asemenea, s-a sugerat că C. bauri era un canibal , pe baza descoperirii unor presupuse exemplare juvenile găsite „în interiorul” cavităților abdominale ale unor exemplare Ghost Ranch. [44] Cu toate acestea, în 2002, Robert J. Gay a demonstrat că acești presupuși minori au fost interpretați greșit. Mai multe exemplare de „coelophysidi juvenile” erau de fapt reptile mici , crurotarsi ca Hesperosuchus și dromomeron . [48] Interpretarea lui Gay a fost susținută de un studiu realizat în 2006 de Nesbitt și colab. . [49] Cu toate acestea, în 2009, au apărut noi dovezi care să susțină un comportament canibalistic atunci când pregătirea ulterioară a matricei recuperate anterior a relevat prezența materialului regurgitat în și în jurul specimenului bucal NMMNH P-44551. Acest material a inclus fragmente de dinți și oase maxilare pe care Rinehart și colab. considerat „identic morfologic” cu o tânără Coelofiză. [8] Mai mult, canibalismul nu este un comportament neobișnuit la arzauri , la fel ca la crocodili , mai ales în situații stresante sau lipsa de hrană.
În 2010, Gay a examinat oasele exemplarelor găsite în cavitatea toracică AMNH 7224 și a calculat că volumul total al acestor oase a fost de 17 ori mai mare decât estimarea maximă a exemplarului de volum de stomac Coelophysis . Gay a observat că volumul total ar fi fost și mai mare dacă s-ar lua în considerare și țesuturile moi. Această analiză a relevat și absența urmelor dentare pe oase și absența coroziunii din acizii stomacului. În cele din urmă, Gay a demonstrat că presupusele oase juvenile canibalizate au fost depuse stratigrafic sub animalul mai mare care ar fi devorat-o. Luate împreună, aceste date sugerează că specimenul AMNH 7224 nu a canibalizat exemplarul mai mic și că rămășițele acestuia din urmă au fost „zdrobite” sub exemplarul mai mare în urma unei suprapuneri aleatorii a indivizilor de diferite dimensiuni. [50]
Comportament social
Descoperirea a peste 1000 de exemplare de carieră de Coelophysis la Whitaker din Ghost Ranch , sugerează că aceste animale manifestă un comportament gregar. [51] Această masivă congregație de rămășițe a făcut să creadă că turmele uriașe ale Coelophysis cutreieră pământul, [44] când, de fapt, nu există dovezi directe ale comportamentului turmei; depozitele indică doar faptul că un număr mare de Coelophysis, împreună cu alte animale triasice, au fost îngropate împreună. Unele dintre dovezile oferite de tafononomia site-ului indică faptul că aceste animale s-ar fi putut aduna împreună pentru a mânca sau bea dintr-o baltă de apă într-o secetă sau pentru a hrăni un puiet de ouă de pește, iar apoi să fie îngropate în timpul unei inundații catastrofale [44]. [51] sau din cauza secetei. [44]
Cu 30 de exemplare de C. rhodesiensis adunate în Zimbabwe , unii paleontologi au sugerat că Coelophysis a fost într-adevăr un animal gregar. Din nou, nu există dovezi directe ale comportamentului turmei și s-a sugerat, de asemenea, că acești indivizi au fost, de asemenea, victime ale inundațiilor fulgerătoare, care au fost destul de frecvente în această perioadă. [30] [52] [53]
Creștere și dimorfism sexual

Rinehart (2009) a evaluat creșterea ontogenică a Coelophysis utilizând date colectate de lungimea femurului și a concluzionat că Coelophysis tânără crește rapid, în special în primul an de viață. [8] Conform estimărilor sale, aceste animale au atins probabil maturitatea sexuală între al doilea și al treilea an de viață și au atins dimensiunea lor maximă, cu puțin peste 3 metri lungime, al optulea an. Acest studiu a identificat, de asemenea, patru etape distincte de creștere: 1 an, 2 ani, 4 ani și 7+ ani. [8] Potrivit rapoartelor, de la naștere, aceste animale au fost autosuficiente și au putut să se descurce singure fără îngrijire parentală extinsă. [44]
Au fost identificate două forme sau „morf” ale Coelophysis: o formă mai subțire, ca specimenul AMNH 7223, și o formă puțin mai robustă, ca exemplarele AMNH 7224 și NMMNH P-42200. Proporțiile scheletice au fost diferite între aceste două forme; [54] forma gracilă are craniul și gâtul mai lung, membrele anterioare mai scurte și coloanele neuronale sacre contopite; forma robustă are un craniu și un gât mai scurt, membrele anterioare mai lungi și coloanele neuronale sacre neacordate. [13] Din punct de vedere istoric, s-au avansat mai multe ipoteze că aceste două forme reprezintă un fel de dimorfism în populația Coelophysis, probabil dimorfism sexual . [7] [13] [55] [56] Raath a fost de acord că dimorfismul în celofiză este evidențiat de dimensiunea și structura membrelor anterioare. [55] Rinehart și colab. el a studiat 15 indivizi și a fost de acord că două morfuri erau prezente, chiar și la cele mai tinere exemplare, sugerând că dimorfismul sexual la aceste animale era prezent și la o vârstă fragedă, înainte de maturitatea sexuală. Rinehart a concluzionat că forma mai scăzută a reprezentat femelele, în timp ce forma puternică a reprezentat masculii, pe baza diferențelor dintre vertebrele sacrale ale formei gracile, care au permis o mai mare flexibilitate pentru ouă. [8] O altă întrebare care susține această ipoteză a venit dintr-o analiză care arată cum fiecare formă cuprindea 50% din populație, așa cum v-ați aștepta într-un raport de 50/50 sexe. [57]
Cu toate acestea, cercetări recente au arătat că C. bauri și C. rhodesiensis au avut o creștere foarte variabilă între indivizi, cu unele exemplare care au fost mai mari în stadiul lor juvenil în comparație cu adulții mai tineri care au atins deja maturitatea sexuală; acest lucru indică faptul că prezența presupusă a unor forme distincte este pur și simplu rezultatul unei variații individuale. Această creștere extrem de variabilă a fost probabil ancestrală pentru dinozauri, dar ulterior s-a pierdut și ar fi putut oferi acestor dinozauri timpurii un avantaj evolutiv în supraviețuirea provocărilor dure de mediu din timpul lor. [58]
Reproducere
Prin compilarea și analiza unei baze de date cu aproape trei duzini de păsări și reptile și compararea cu datele existente despre anatomia Coelophysis, Rinehart și colab. (2009) au tras următoarele concluzii. S-a estimat că o coelofiză medie a ouălor a fost de 31 până la 33,5 mm prin diametrul său mai mic și că fiecare femelă depune între 24 și 26 de ouă pentru incubație. Unele dovezi sugerează, de asemenea, că cel puțin o îngrijire parentală a fost prezentă pentru a hrăni tinerii în primul lor an de viață, unde ar fi atins 1,5 metri lungime până la sfârșitul primei lor etape de creștere. Coelofiza a investit în reproducere la fel de multă energie ca și alte reptile dispărute de dimensiunea sa aproximativă. [8] [59]
Paleopatologie
Într-un studiu din 2001 realizat de Bruce Rothschild și alți paleontologi, oasele au fost examinate de 14 picioare legate de Coelophysis pentru semne de fractură de stres, dar niciunul nu a fost găsit. [60]
La C. rhodesiensis s-au observat fracturi vindecate ale tibiei și metatarsului , dar sunt foarte rare. O parte din a doua coastă sacrală într-un specimen de Syntarsus rhodesiensis prezintă semne de asimetrie fluctuantă. Asimetria fluctuantă apare din tulburările de dezvoltare și este mai frecventă la populațiile stresate și, prin urmare, poate fi informativă despre calitatea condițiilor în care a trăit un dinozaur.[61]
Paleoecologie

Dintre exemplarele de Coelophysis au fost găsite și recuperate din Formația Chinle din New Mexico și Arizona , cel mai notoriu la cariera Ghost Ranch (Whitaker), membru Rock Point [8] printre alte cariere din membrul de sub Pădurea Petrificată, ale cărei sedimente au fost datate cu aproximativ 212 de milioane de ani în urmă, făcându-i parte din faza mediului norian al triasicului superior . [2] [44]
C. rhodesiensis a fost găsit în formațiunea superioară Elliot din provinciile Cape și din statul liber al Africii de Sud , precum și în golul albiului oaselor chitake ale râului la Formația Forest Sandstone din Zimbabwe .
Ghost Ranch a fost situat în apropierea ecuatorului în urmă cu peste 200 de milioane de ani și avea un climat cald, aproape musonal, cu precipitații sezoniere abundente. Hayden Quarry, un nuovo sito di scavo a Ghost Ranch, nel Nuovo Messico, ha portato alla luce una collezione diversificata di materiale fossile che includeva le prime prove di dinosauri e dinosauromorfi dello stesso periodo. La scoperta indica che i due gruppi vivevano insieme durante il primo periodo del Triassico, circa 235 milioni di anni fa. [62]
Therrien e Fastovsky (2001) hanno esaminato il paleoambiente di Coelophysis e altri primi teropodi del Petrified Forest National Park in Arizona, e hanno determinato che questo genere viveva in un ambiente costituito da pianure alluvionali contrassegnate da stagioni secche e umide distinte. C'era una grande competizione durante i periodi più secchi, quando gli animali lottavano per l'acqua nei letti dei fiumi che si stavano prosciugando. [63]
Nelle sezioni superiori della Formazione Chinle dove è stato ritrovato Coelophysis , i dinosauri erano più rari e finora sono noti solo Chindesaurus e Daemonosaurus . [64] La fauna terrestre è invece dominata da altri rettili come il rincocefalo Whitakersaurus [65] ; lo pseudosucho Revueltosaurus [66] ; gli aetosauri Desmatosuchus [66] , Typothorax [66] e Heliocanthus [66] ; il crocodilomorfo Hesperosuchus [66] ; i " rauisuchi " Shuvosaurus [66] , Effigia [67] e Vivaron [68] ; insieme ad altri componenti rari come il dinosauriforme Eucoelophysis [69] , e l' amniote Kraterokheirodon . [66] Nei corsi d'acqua troviamo il fitosauro Machaeroprosopus [66] ; l' arcosauromorfo Vancleavea [66] ; gli anfibi Apachesaurus [66] e Koskinonodon [70] ; ei pesci Reticulodus [66] , Arganodus [66] e Lasalichthyes . [66]
Si pensa che la Formazione Elliot superiore sia stata un'antica pianura alluvionale. I fossili dei prosauropodi Massospondylus e Ignavusaurus sono stati recuperati dalla Formazione di Elliot superiore, che vanta la fauna più diversificata al mondo di dinosauri ornitischi del primo Giurassico, tra cui Abrictosaurus , Fabrosaurus , Heterodontosaurus e Lesothosaurus , tra gli altri. La Formazione Forest Sandstone era il paleoambiente di coccodrilli protosuchidi, sfenodonti, il dinosauro Massospondylus e resti indeterminati di un altro prosauropode. Paul (1988) ha notato che Megapnosaurus rhodesiensis viveva tra le dune del deserto e le oasi e cacciava giovani prosauropodi. [71]
Tafonomia
La moltitudine di esemplari depositatisi così vicini gli uni agli altri a Ghost Ranch fu probabilmente il risultato di un'inondazione improvvisa, che spazzò via un gran numero di Coelophysis e li seppellì rapidamente e contemporaneamente. In effetti, sembra che tali inondazioni fossero all'ordine del giorno durante questo periodo della storia della Terra e, in effetti, la foresta pietrificata della vicina Arizona è il risultato di un blocco di tronchi d'albero conservato che sono stati trascinati via da una di queste inondazioni. La cava Whitaker a Ghost Ranch è considerata un sito monotassico perché presenta più individui di un singolo taxon. La qualità della conservazione e la gamma ontogenica (età) dei campioni hanno contribuito a rendere Coelophysis uno dei più noti di tutti i generi di dinosauro. [8] [72]
I 30 esemplari di C. rhodesiensis ritrovati insieme nello Zimbabwe sono probabilmente anch'essi il risultato di un'alluvione improvvisa, che ha spazzato via un gran numero di Coelophysis e seppellito rapidamente e contemporaneamente i loro corpi. [30] [52] [53]
Icnologia

Edwin H. Colbert suggerì che le impronte di teropodi riferite all' icnogenere Grallator , rinvenute nella valle del fiume Connecticut attraverso il Connecticut e il Massachusetts, potrebbero appartenere a Coelophysis . [73] Le impronte provengono dal Supergruppo di Newark datato dal Triassico superiore all'inizio del Giurassico. Le impronte mostrano chiaramente le dita II III e IV ma non I o V. Questa condizione è strana per le impronte della loro età. Si presumeva che queste dita fossero tozze e vestigiali, non toccando il suolo quando il dinosauro camminava o correva. Per questo è stato ipotizzato che appartenessero in realtà ad un saurischiano primitivo non identificato simile a Coelophysis da David B. Weishampel e L. Young. [74] Nella valle sono stati trovati anche resti scheletrici simili a Coelophysis , a sostegno dell'idea che una specie simile a Coelophysis possa aver lasciato queste impronte. [75]
Significato culturale
Coelophysis è stato il secondo dinosauro nello spazio, dopo Maiasaura ( STS-51-F ). [76] Un cranio di Coelophysis del Carnegie Natural History Museum era a bordo della Space Shuttle Endeavour missione STS-89 quando ha lasciato l'atmosfera il 22 gennaio 1998. È stato anche preso sulla stazione spaziale Mir , prima di essere riportato sulla Terra. [76] [77]
Essendo stato nominato più di 100 anni fa, Coelophysis è uno dei dinosauri più conosciuti in letteratura. È stato designato come fossile ufficiale dello stato del Nuovo Messico nel 1981 ed è ora il logo del New Mexico Museum of Natural History. [21] [78]
Note
- ^ RE: Afrovenator pronunciation , su dml.cmnh.org . URL consultato il 26 maggio 2019 .
- ^ a b Randall B. Irmis, Roland Mundil, Jeffrey W. Martz e William G. Parker, High-resolution U–Pb ages from the Upper Triassic Chinle Formation (New Mexico, USA) support a diachronous rise of dinosaurs , in Earth and Planetary Science Letters , vol. 309, 3–4, 15 Settembre 2011, pp. 258–267, Bibcode : 2011E&PSL.309..258I , DOI : 10.1016/j.epsl.2011.07.015 .
- ^ a b Richard M. Gaines, Coelophysis , ABDO Publishing Company, 2001, p. 4, ISBN 978-1-57765-488-9 .
- ^ Weishampel, David B; et al. (2004). "Dinosaur distribution (Early Jurassic, Africa)." In: Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.): The Dinosauria, 2nd, Berkeley: University of California Press. Pp. 535–536. ISBN 0-520-24209-2 .
- ^ a b c d ( EN ) Martín D Ezcurra, Richard J Butler, Susannah CR Maidment, Ivan J Sansom, Luke E Meade e Jonathan D Radley, A revision of the early neotheropod genus Sarcosaurus from the Early Jurassic (Hettangian–Sinemurian) of central England , in Zoological Journal of the Linnean Society , vol. 191, n. 1, 1º Gennaio 2021, pp. 113–149, DOI : 10.1093/zoolinnean/zlaa054 , ISSN 0024-4082 .
- ^ a b Hilde L. Schwartz e Gillette, David D., Geology and taphonomy of the Coelophysis quarry, Upper Triassic Chinle Formation, Ghost Ranch, New Mexico , in Journal of Paleontology , vol. 68, n. 5, 1994, pp. 1118–1130, DOI : 10.1017/S0022336000026718 , JSTOR 1306181 .
- ^ a b c d Gregory S. Paul , Predatory Dinosaurs of the World , Simon & Schuster , 1988, pp. 260 , ISBN 978-0-671-61946-6 .
- ^ a b c d e f g h i j k Rinehart, LF; Lucas, SG; Heckert, AB; Spielmann, JA & Celesky, MD, The paleobiology of Coelophysis bauri (Cope) from the Upper Triassic (Apachean) Whitaker quarry, New Mexico, with detailed analysis of a single quarry block , in New Mexico Museum of Natural History & Science, A Division of the Department of Cultural Affairs Bulletin , vol. 45, 2009, pp. 260.
- ^ Larry F. Rinehart, Spencer G. Lucas e Adrian P. Hunt, Furculae in the Late Triassic theropod dinosaur Coelophysis bauri , in Paläontologische Zeitschrift , vol. 81, n. 2, 2007, pp. 174–180, DOI : 10.1007/bf02988391 .
- ^ a b c Tykoski, RS & Rowe, T. (2004). "Ceratosauria": In: The Dinosauria , Seconda, Berkeley, University of California Press, 6 Novembre 2004, pp. 47–70, ISBN 978-0-520-24209-8 .
- ^ Gay, RJ, An unusual adaptation in the caudal vertebrae of Coelophysis bauri (Dinosauria: Theropoda) , in PaleoBios , vol. 21, 2001, pp. 55.
- ^ Rinehart, LF; Heckert, AB; Lucas, SG & Hunt, AP, The sclerotic ring of the Late Triassic theropod dinosaur Coelophysis , in New Mexico Geological Society Spring Meeting , vol. 26, 2004, pp. 64.
- ^ a b c d e Colbert, E., The Triassic Dinosaur Coelophysis , in Museum of Northern Arizona Bulletin , vol. 57, 1989, pp. 160.
- ^ Ezcurra, MD, The cranial anatomy of the coelophysoid theropod Zupaysaurus rougieri from the Upper Triassic of Argentina , in Historical Biology , vol. 19, n. 2, 2007, pp. 185–202, DOI : 10.1080/08912960600861467 .
- ^ a b Bristowe, A. & Raath, MA, A juvenile coelophysoid skull from the Early Jurassic of Zimbabwe, and the synonymy of Coelophysis and Syntarsus ( PDF ), in Palaeontologia Africana , vol. 40, 2004, pp. 31–41, ISSN 0078-8554 .
- ^ a b c Downs, A. (2000). " Coelophysis bauri and Syntarsus rhodesiensis compared, with comments on the preparation and preservation of fossils from the Ghost Ranch Coelophysis quarry": In: Dinosaurs of New Mexico , in New Mexico Museum of Natural History Bulletin , vol. 17, pp. 33–37.
- ^ a b c d e f Bristowe, A. & MA Raath, A juvenile coelophysoid skull from the Early Jurassic of Zimbabwe, and the synonymy of Coelophysis and Syntarsus .(USA) , in Palaeontologica Africana , vol. 40, n. 40, 2004, pp. 31–41.
- ^ Coelophysoidea , su theropoddatabase.com . URL consultato il 5 febbraio 2016 .
- ^ Facial variation in Coelophysis bauri and the status of Megapnosaurus (Syntarsus) , su nmstatefossil.org . URL consultato il 5 febbraio 2016 .
- ^ Adrian P. Hunt e Spencer G. Lucas, Rioarribasaurus , a new name for a Late Triassic dinosaur from New Mexico (USA) , in Paläontologische Zeitschrift , vol. 65, n. 1/2, 1991, pp. 191–198, DOI : 10.1007/bf02985783 .
- ^ a b c Colbert, EH; Charig, AJ; Dodson, P.; Gillette, DD; Ostrom, JH & Weishampel, DB, Coelurus bauri Cope, 1887 (currently Coelophysis bauri ; Reptilia, Saurischia): Proposed replacement of the lectotype by a neotype ( PDF ), in Bulletin of Zoological Nomenclature , vol. 49, n. 4, 1992, pp. 276–279.
- ^ a b International Commission on Zoological Nomenclature, Opinion 1842: Coelurus bauri Cope, 1887 (currently Coelophysis bauri ; Reptilia, Saurischia): lectotype replaced by a neotype ( PDF ), in Bulletin of Zoological Nomenclature , vol. 53, n. 2, 1996, pp. 142–144.
- ^ Talbot, M., Podokesaurus holyokensis , a new dinosaur from the Triassic of the Connecticut Valley , in American Journal of Science , 4, vol. 31, n. 186, 1911, pp. 469–479, Bibcode : 1911AmJS...31..469T , DOI : 10.2475/ajs.s4-31.186.469 .
- ^ E. Colbert, The Triassic dinosaur genera Podokesaurus and Coelophysis , in American Museum Novitates , vol. 2168, 1964, pp. 1–12.
- ^ E. Colbert e Baird, D., Coelurosaur bone casts from the Connecticut Valley Triassic , in American Museum Novitates , vol. 1901, 1958, pp. 1–11.
- ^ PR Getty e AM Bush, Sand pseudomorphs of dinosaur bones: Implications for (non-) preservation of tetrapod skeletal material in the Hartford Basin, USA , in Palaeogeography, Palaeoclimatology, Palaeoecology , vol. 302, 3–4, 2011, pp. 407–414, DOI : 10.1016/j.palaeo.2011.01.029 .
- ^ Olshevsky, G., A revision of the parainfraclass Archosauria Cope, 1869, excluding the advanced Crocodylia ( PDF ), in Mesozoic Meanderings 2 , 1991, pp. 196.
- ^ Robert M. Sullivan e Spencer G. Lucas, Eucoelophysis baldwini , a new theropod dinosaur from the Upper Triassic of New Mexico, and the status of the original types of Coelophysis , in Journal of Vertebrate Paleontology , vol. 19, n. 1, 1999, pp. 81–90, DOI : 10.1080/02724634.1999.10011124 .
- ^ Sterling J. Nesbitt, Randall B. Irmis e William G. Parker, A critical re-evaluation of the Late Triassic dinosaur taxa of North America , in Journal of Systematic Palaeontology , vol. 5, n. 2, 2007, pp. 209–243, DOI : 10.1017/S1477201907002040 .
- ^ a b c d Raath (1969). "A new Coelurosaurian dinosaur from the Forest Sandstone of Rhodesia." Arnoldia Rhodesia. 4 (28): 1–25.
- ^ MA Ivie, SA Ślipiński e P. Węgrzynowicz, Generic homonyms in the Colydiinae (Coleoptera: Zopheridae) , in Insecta Mundi , vol. 15, 2001, pp. 63–64.
- ^ Mickey Mortimer, Coelophysoidea , su archosaur.us , 2012 (archiviato dall' url originale il 4 maggio 2013) .
- ^ Yates, AM, A new theropod dinosaur from the Early Jurassic of South Africa and its implications for the early evolution of theropods , in Palaeontologia Africana , vol. 41, 2005, pp. 105–122.
- ^ Paul GS (1993). "Are Syntarsus and the Whitetaker theropod the same genus?": In: SG Lucas e M. Morales (a cura di), The Nonmarine Triassic , in New Mexico Museum of Natural History Bulletin , vol. 3, pp. 397–402.
- ^ MD Ezcurra e Brusatte, SL, Taxonomic and phylogenetic reassessment of the early neotheropod dinosaur Camposaurus arizonensis from the Late Triassic of North America , in Palaeontology , vol. 54, n. 4, 2011, pp. 763–772, DOI : 10.1111/j.1475-4983.2011.01069.x .
- ^ Sues, HD, Nesbitt, SJ, Berman, DS e Henrici, AC,A late-surviving basal theropod dinosaur from the latest Triassic of North America , in Proceedings of the Royal Society B , vol. 278, n. 1723, 2011, pp. 3459–3464, DOI : 10.1098/rspb.2011.0410 , PMC 3177637 , PMID 21490016 .
- ^ AD Marsh e TB Rowe, A comprehensive anatomical and phylogenetic evaluation of Dilophosaurus wetherilli (Dinosauria, Theropoda) with descriptions of new specimens from the Kayenta Formation of northern Arizona , in Journal of Paleontology , vol. 94, n. 78, 2020, pp. 1–103, DOI : 10.1017/jpa.2020.14 .
- ^ a b Matthew T. Carrano, Roger BJ Benson e Scott D. Sampson, The phylogeny of Tetanurae (Dinosauria: Theropoda) , in Journal of Systematic Palaeontology , vol. 10, n. 2, 2012, pp. 211–300, DOI : 10.1080/14772019.2011.630927 .
- ^ Sterling J. Nesbitt, Randall B. Irmis e William G. Parker, A critical re-evaluation of the Late Triassic dinosaur taxa of North America ( PDF ), in Journal of Paleontology , vol. 5, n. 2, 2007, pp. 209–243, DOI : 10.1017/s1477201907002040 .
- ^ Nesbitt, Irmis and Parker, 2007. A critical re-evaluation of the Late Triassic dinosaur taxa of North America. Journal of Systematic Palaeontology. 5(2), 209–243.
- ^ a b c Cope, ED, On a new genus of Triassic Dinosauria , in The American Naturalist , vol. 23, n. 271, 1889, pp. 621–633, DOI : 10.1086/274979 .
- ^ a b Glut, DF, Dinosaurs, the Encyclopedia, Supplement 1 , McFarland & Company, Inc., 1999, pp. 442, ISBN 978-0-7864-0591-6 .
- ^ Cope, ED (1887). "The Dinosaurian Genus Coelurus". The American Naturalist. xxi 5: 367-369.
- ^ a b c d e f g Haines, T. & Chambers, P., The Complete Guide to Prehistoric Life , Firefly Books, 2007, pp. 70 –71, ISBN 978-1-55407-181-4 .
- ^ Tykoski, RS, and Rowe, T., 2004, Ceratosauria, Chapter Three: In: The Dinosauria, Second Edition, edited by Weishampel, DB, Dodson, P., and Osmolska, H., California University Press, p. 47-70.
- ^ Benton, M., Dinosaur and other prehistoric animal Fact Finder , Kingfisher Books Ldt., 1993, pp. 256, ISBN 978-0-86272-949-3 .
- ^ Carpenter, K., Forelimb bio-mechanics of non-avian theropod dinosaurs in predation , in Senckenbergiana Lethaea , vol. 82, 2002, pp. 59–76, DOI : 10.1007/bf03043773 .
- ^ Gay, RJ, The myth of cannibalism in Coelophysis bauri , in Journal of Vertebrate Paleontology , vol. 22, n. 3, 2002, pp. 57A.
- ^ SJ Nesbitt, A. H Turner, G. M Erickson e M. A Norell,Prey choice and cannibalistic behaviour in the theropod Coelophysis , in Biology Letters , 22, vol. 2, n. 4, 2006, pp. 611–614, DOI : 10.1098/rsbl.2006.0524 , PMC 1834007 , PMID 17148302 .
- ^ Gay, RJ, Notes on Early Mesozoic Theropods (First ed.) , Lulu press, 2010, pp. 9–24, ISBN 978-0-557-46616-0 .
- ^ a b Schwartz, HL & Gillette, DD, Geology and taphonomy of the Coelophysis quarry, Upper Triassic Chinle Formation, Ghost Ranch, New Mexico , in Journal of Paleontology , vol. 68, n. 5, 1994, pp. 1118–1130, DOI : 10.1017/S0022336000026718 , JSTOR 1306181 .
- ^ a b G. Bond, 1965. Some new fossil localities in the Karroo System of Rhodesia. Arnoldia, Series of Miscellaneous Publications, National Museum of Southern Rhodesia 2(11):1–4
- ^ a b MA Raath, 1977. The Anatomy of the Triassic Theropod Syntarsus rhodesiensis (Saurischia: Podokesauridae) and a Consideration of Its Biology. Department of Zoology and Entomology, Rhodes University, Salisbury, Rhodesia 1–233
- ^ Gregory S. Paul,The Princeton Field Guide to Dinosaurs , Princeton University Press, 2010, p. 74 , ISBN 978-0-691-13720-9 .
- ^ a b Carpenter, K. e Currie, PJ, Dinosaur Systematics: Approaches and Perspectives , Cambridge, Cambridge University Press, 1990, pp. 91–105, ISBN 978-0-521-43810-0 .
- ^ Gay, R. (2005). "Sexual Dimorphism in the Early Jurassic Theropod Dinosaur Dilophosaurus and a Comparison with Other Related Forms": In: Carpenter, K. (a cura di), The Carnivorous Dinosaurs , Bloomington, Indiana University Press, 2005, pp. 277–283, ISBN 978-0-253-34539-4 .
- ^ Rinehart, LF, Lucas, SG e Heckert, AB, Preliminary statistical analysis defining the juvenile, robust and gracile forms of the Triassic dinosaur Coelophysis , in Journal of Vertebrate Paleontology , vol. 21, 2001, pp. 93A, DOI : 10.1080/02724634.2001.10010852 .
- ^ CT Griffin e SJ Nesbitt, Anomalously high variation in postnatal development is ancestral for dinosaurs but lost in birds , in Proceedings of the National Academy of Sciences of the United States of America , vol. 113, n. 51, 2016, pp. 14757–14762, Bibcode : 2016PNAS..11314757G , DOI : 10.1073/pnas.1613813113 .
- ^ Glut, DF, Dinosaurs, the Encyclopedia, Supplement 7 , McFarland & Company, Inc., 2012, pp. 866, ISBN 978-0-7864-4859-3 .
- ^ Rothschild, B.; Tanke, DH & Ford, TL (2001). "Theropod stress fractures and tendon avulsions as a clue to activity": In: DH Tanke e K. Carpenter (a cura di), Mesozoic Vertebrate Life , Indiana University Press, 2001, pp. 331–336, ISBN 978-0-253-33907-2 .
- ^ Molnar, RE, 2001, Theropod paleopathology: a literature survey: In: Mesozoic Vertebrate Life, edited by Tanke, DH, and Carpenter, K., Indiana University Press, p. 337-363.
- ^ Donna Braginetz, A new species of dinosauromorph (lower left) was among the mixed assemblage of dinosaurs and dinosauromorphs found at Hayden Quarry in Ghost Ranch, NM , su amnh.org , American Museum of Natural History, 2007. URL consultato il 31 Marzo 2013 .
- ^ F. Therrien e DE Fastovsky, Paleoenvironments of early theropods, Chinle Formation (Late Triassic), Petrified Forest National Park, Arizona , in PALAIOS , 3, vol. 15, n. 3, 2000, pp. 194–211, Bibcode : 2000Palai..15..194T , DOI : 10.1669/0883-1351(2000)015<0194:POETCF>2.0.CO;2 .
- ^ H. -D. Sues, SJ Nesbitt, DS Berman e AC Henrici,A late-surviving basal theropod dinosaur from the latest Triassic of North America , in Proceedings of the Royal Society B: Biological Sciences , vol. 278, n. 1723, 2011, pp. 3459–64, DOI : 10.1098/rspb.2011.0410 , PMC 3177637 , PMID 21490016 .
- ^ AB Heckert, SG Lucas, LF Rinehart e AP Hunt, A new genus and species of sphenodontian from the Ghost Ranch Coelophysis Quarry (Upper Triassic: Apachean), Rock Point Formation, New Mexico, USA , in Palaeontology , vol. 51, n. 4, 2008, pp. 827–845, DOI : 10.1111/j.1475-4983.2008.00786.x .
- ^ a b c d e f g h i j k l m Irmis, RB (2005). "The vertebrate fauna of the Upper Triassic Chinle Formation in northern Arizona": In: SJ Nesbitt, WG Parker e RB Irmis (a cura di), Guidebook to the Triassic formations of the Colorado Plateau in northern Arizona: Geology, Paleontology, and History ( PDF ), in Mesa Southwest Museum Bulletin , vol. 9, pp. 63–88. URL consultato il 2 Gennaio 2014 (archiviato dall' url originale il 4 ottobre 2012) .
- ^ Benson, RJ; Brusatte, S.; Clack, J.; Anderson, J.; Dennis-Bryan, K.; Hone, D.; Duffin, C.; Johanson, Z.; Milner, A.; Naish, D.; Parsons, K.; Prothero, D.; Xu, X., Prehistoric Life , Londra, Dorling Kindersley, 2012, pp. 214–215, ISBN 978-0-7566-9910-9 .
- ^ Emily J. Lessner, Michelle R. Stocker, Nathan D. Smith, Alan H. Turner, Randall B. Irmis e Sterling J. Nesbitt,A new rauisuchid (Archosauria, Pseudosuchia) from the Upper Triassic (Norian) of New Mexico increases the diversity and temporal range of the clade , in PeerJ , vol. 4, 2016, pp. e2336, DOI : 10.7717/peerj.2336 , PMC 5018681 , PMID 27651983 .
- ^ Weishampel, DB; Dodson, P.; & Osmólska, H., The Dinosauria (Second ed.) , Berkeley, University of California Press, 2004, pp. 518–521, ISBN 978-0-520-24209-8 .
- ^ Mueller, BD, [225:KBAMAR 2.0.CO;2 Koskinonodon Branson and Mehl, 1929, a replacement name for the preoccupied temnospondyl Buettneria Case, 1922 ], in Journal of Vertebrate Paleontology , vol. 27, n. 1, 2007, pp. 225, DOI : 10.1671/0272-4634(2007)27[225:KBAMAR]2.0.CO;2 .
- ^ Paul, GS, 1988, Predatory Dinosaurs of the World, a complete Illustrated guide: New York Academy of sciences book, 464pp.
- ^ Glut, DF, Dinosaurs, the Encyclopedia, Supplement 4 , McFarland & Company, Inc., 2006, pp. 749, ISBN 978-0-7864-2295-1 .
- ^ Edwin H. Colbert , The Age of Reptiles , WW Norton & Company , 1965, pp. 97 , ISBN 978-0-486-29377-6 .
- ^ Weishampel, DB e Young, L., Dinosaurs of the East Coast , Johns Hopkins University Press, 1998, ISBN 978-0-8018-5217-6 .
- ^ Dinosaur footprints of the Connecticut River Valley , su nashdinosaurtracks.com , Nash Dinosaur Track Site and Rock Shop. URL consultato il 16 Giugno 2015 (archiviato dall' url originale il 1º gennaio 2017) .
- ^ a b Chure, D., dino bones in space – was it a PR thing , su dml.cmnh.org , Cleveland Museum of Natural History, 2009. URL consultato il 12 Novembre 2011 .
- ^ Steve Parker, Dinosaurs: the complete guide to dinosaurs , Firefly Books, 2003, ISBN 978-1-55297-772-9 .
- ^ https://geoinfo.nmt.edu/publications/periodicals/earthmatters/17/n2/em_v17_n2.pdf
Altri progetti
-
 Wikimedia Commons contiene immagini o altri file su Coelophysis
Wikimedia Commons contiene immagini o altri file su Coelophysis -
 Wikispecies contiene informazioni su Coelophysis
Wikispecies contiene informazioni su Coelophysis
Collegamenti esterni
- Coelophysis nella Dino Directory
- sul ritrovamento dei celofisi a Ghost Ranch e su quello che potrebbe essere accaduto.