Aenocyon dirus
| Enocion | |
|---|---|
 | |
| Starea de conservare | |
Fosil | |
| Clasificare științifică | |
| Domeniu | Eukaryota |
| Regatul | Animalia |
| Phylum | Chordata |
| Clasă | Mammalia |
| Ordin | Carnivore |
| Familie | Canide |
| Subfamilie | Caninae |
| Trib | canini |
| Subtrib | Canin |
| Tip | Enocion |
| Specii | † A. dirus |
| Nomenclatura binominala | |
| † Aenocyon dirus Leidy , 1858 | |
| Sinonime | |
| |
Enocionul ( Aenocyon dirus Leidy , 1858 ) este o specie dispărută de canid de lupin care a trăit în timpul Pleistocenului (între 200.000 și 10.000 de ani în urmă) în America și Asia de Est . Deși s-a crezut de multă vreme că este un membru al genului Canis , analizele genomice ale rămășițelor nefosilizate indică faptul că este de fapt o descendență care s-a îndepărtat de alte canide de lupin acum 5,7 milioane de ani, demonstrând o rudenie mai strânsă cu șacalii africani . Sunt recunoscute două subspecii: A. d. dirus , care a locuit zonele de la est de diviziunea continentală și A. d. guildayi din Mexic și California, mai mici și cu membrele mai scurte, dar cu dinții mai lungi.
Enocionul avea cam dimensiunea unui lup cenușiu mare astăzi, putând ajunge la 68 de kilograme. Craniul și dentiția lui erau similare cu cele ale lupului cenușiu, dar dinții lui erau mai groși și mai ascuțiți, iar forța mușcăturii sale asupra caninilor era mai mare decât cea a caninilor de astăzi. Se consideră că aceste caracteristici au fost adaptări la eliminarea imenselor erbivore din Pleistocenul inferior . În America de Nord, prada includea cai , leneși , mastodonti , bizoni și cămile . La fel ca canidele mari de astăzi, este posibil să fi vânat în haite, cu o structură socială primitivă și tehnici de vânătoare cooperative.
Rămășițele sale au fost găsite în numeroase habitate, inclusiv câmpii, câmpuri, anumite zone forestiere din America de Nord, savane din America de Sud și stepele din Asia de Est. Majoritatea înregistrărilor fosile au fost obținute în gropile de gudron Rancho La Brea din Los Angeles . Înregistrările fosile se găsesc în zone înalte de la nivelul mării până la 2 255 de metri. Specia era aparent rară deasupra paralelului 42 nord , cu doar cinci exemplare găsite dincolo de această latitudine în America de Nord. Această limitare areală pare a fi datorată temperaturii, disponibilității prăzii sau habitatului impus speciei de apropierea de ghețarii Laurentidi și Cordillerani. Cel puțin o rămășiță fosilă a fost găsită în nord-estul Chinei, ceea ce indică faptul că unele exemplare au reușit să traverseze Podul Bering , dar specia nu a reușit niciodată să prospere în Asia ca și pe continentul de origine, probabil din cauza concurenței cu hienele peșterii .
Dispariția sa s-a încheiat în perioada cuaternară , împreună cu prada principală. Dependența sa de prada mare a fost citată ca fiind cauza morții sale, împreună cu schimbările climatice și competiția cu alți prădători, inclusiv cu oamenii care s-au răspândit pe continent la sfârșitul ultimei ere glaciare prin reducerea resurselor alimentare ale animalului . [1] Un alt factor poate fi incapacitatea enocionului de a se încrucișa cu alte specii de canide mai adaptabile. Cele mai recente rămășițe ale sale datează de 9.440 de ani.
Taxonomie

Începând cu anii 1850, au fost găsite numeroase fosile de canide mari cu identitate taxonomică incertă. Primul exemplar care ar fi asociat ulterior cu A. dirus [3] a fost găsit în 1854 în Evansville , Indiana . O mandibulă fosilizată a fost obținută de omul de știință Joseph Granville Norwood de la colecționarul Francis A. Linck. Paleontologul Joseph Leidy a stabilit că specimenul reprezenta un fel de lup dispărut și l-a numit Canis primaevus . [4] Scrisorile pe care Leidy le-a primit de la Norwood sunt încă păstrate împreună cu tipul nomenclatural la academia de științe naturale. În 1857, în timp ce explora râul Niobrara, Nebraska, Leidy a descoperit vertebrele unui canid dispărut, pe care l-a numit Canis dirus în anul următor. [5] Denumirea de C. primaevus a fost ulterior aruncată în favoarea lui C. indianensis , deoarece C. primaevus a fost deja folosit pentru cuonul alpin de astăzi . [6]
În 1876, după descoperirea și denumirea C. mississippiensis , Joel Asaph Allen a avansat ipoteza că C. indianensis și C. dirus erau de fapt aceleași specii. Întrucât descoperirile atribuibile celor trei exemplare au fost rare, s-a decis păstrarea denumirilor provizorii până la descoperirea unor materiale suplimentare care să servească la clarificarea problemei identității lor. [7] În 1912, paleontologul John Campbell Merriam a decis să clasifice toate exemplarele ca sinonime ale lui C. dirus , primul dintre numele propuse pentru specie. [8] În conformitate cu propunerea lui Merriam, C. indianensis a fost retrogradată în rolul de sinonim pentru C. dirus în 1915. [9] În 1918, Merriam a propus clasificarea animalului sub un nou gen , Aenocyon (din grecescul Aenos : teribil, și cyon : lup); [10] Cu toate acestea, nu toți au acceptat noua clasificare, iar propunerea a rămas un subiect controversat în rândul paleontologilor. [11] În 1972, C. ayersi și A. dirus erau sinonime cu C. dirus . [12] [13] În cele din urmă, toți taxonii deja descriși erau sinonimi cu C. dirus în 1979 de paleontologul Ronald M. Nowak. [14]
În 1984, Björn Kurten a propus denumirea de exemplare din California și Mexic, notabile pentru membrele lor scurte și dinții alungiți, C. d. guildayi , în timp ce cele de la est de diviziunea continentală , care demonstrează membrele mai lungi și dinții mai scurți, cum ar fi C. d. dirus . [15] [16] [17] [18]
Morfologie
Cei trei paleontologi Xiaoming Wang, Richard H. Tedford și RM Nowak au propus că A. dirus derivă din canidul mare C. armbrusteri . [19] [20] [21] Nowak însuși a afirmat că există exemplare găsite din peștera Cumberland din Maryland care par să aparțină unui stadiu intermediar între C. armbrusteri și A. dirus . [22] [23] C. chihliensis din China poate fi strămoșul comun atât al lui C. armbrusteri, cât și al lupului gri de astăzi. [19] Apariția bruscă a C. armbrusteri în America de Nord în Pleistocenul superior indică faptul că a fost un animal care a imigrat din Asia, ca în cazul lupului cenușiu din timpul Pleistocenului . [19] Unii paleontologi nu sunt de acord cu această teorie, argumentând în schimb că C. armbrusteri este o formă arhaică de lup cenușiu. [24] În acord cu această teză, în 1993 paleontologul Goulet a afirmat că A. dirus era de fapt o variantă a lupului gri. [25] Nowak, Kurten și alții au contestat această clasificare, afirmând că A. dirus nu era o derivare a lupului gri. [26] [27] Ulterior s-a avansat ipoteza că A. dirus evoluase paralel cu lupul cenușiu. [28]
A. dirus a trăit de la Pleistocenul superior la Holocenul inferior (acum 125.000-10.000 de ani) în America . [15] Cele mai multe fosile ale subspeciei A. d. dirus poate fi urmărit în urmă cu 125.000-75.000 de ani în urmă, în timp ce cele din A. d. guildayi , pe lângă faptul că sunt mai mici, sunt mai recente. Prin urmare, s-a propus ca A. d. guildayi este derivat din A. d. dirus . [15] [18] Există, totuși, exemplare de A. dirus de clasificare dubioasă, datând de acum 250.000 de ani. Înregistrările de fosile găsite în patru situri din zona Hay Springs (județul Sheridan, Nebraska) au fost denumite Aenocyon dirus nebrascensis , dar nu aveau o descriere. Aceste rămășițe au fost clasificate ulterior sub denumirea de C. armbrusteri . [14] O descriere a exemplarelor publicate ulterior le-a clasificat ca aparținând speciei A. dirus , deși prezentau trăsături comune cu C. armbrusteri . [19] Este posibil ca exemplarele din Nebraska să reprezinte cele mai arhaice forme ale speciei. [19] [29]
O fosilă găsită din așa-numita hală a cailor din peștera Salamandrei din Dealurile Negre din Dakota de Sud poate reprezenta cel mai vechi exemplar de A. dirus descoperit până în prezent. [12] [30] Animalul a fost catalogat ca C. cf. C. dirus , dată fiind identitatea sa incertă. [31] O fosilă ecvină găsită în peșteră a dat o dată radiometrică de acum 252.000 de ani și s-a presupus că exemplarul lui A. dirus era de aceeași vârstă. [30] [31] Atât C. armbrusteri, cât și A. dirus prezintă o serie de caracteristici comune , sugerând că originea lui A. dirus datează de acum aproximativ 250.000 de ani în zonele deschise ale continentului central și apoi s-a extins spre est. și înlocuiește în totalitate strămoșul său C. armbrusteri . [19] Acest lucru ar indica faptul că specia a apărut acum 250.000 de ani în California și Nebraska, pentru a coloniza ulterior Canada, restul Statelor Unite, Mexic, Venezuela, Ecuador, Bolivia și Peru; [19] identitatea acestor fosile antice rămâne totuși controversată. [32]
În America de Sud, fosilele de A. dirus care pot fi urmărite până la Pleistocenul superior au fost găsite pe coastele vestice și nordice, dar nu au fost găsite exemplare în Argentina, unde au trăit C. gezi și C. nehringi . [19] Un studiu a arătat că A. dirus este o specie morfologic mai derivată în comparație cu canidele americane anterioare din genul Canis , fiind mai mare decât C. nehringi , cu molari mai dezvoltați. [27] Pe baza acestor observații, unii cercetători au propus o origine sud-americană pentru specie. [14] [26] [30] În 2009, s-a propus că C. armbrusteri a fost strămoșul comun al lupilor nord-americani și sud-americani. [19] În anul următor, un alt studiu a furnizat dovezi că A. dirus și C. nehringi sunt de fapt aceleași specii, indicând faptul că prima a ajuns în America de Sud din nord. [28]
Analiza ADN
Încercările de extragere a ADN-ului din fosilele găsite în gropile de gudron nu au avut succes. În 1992 s-a încercat extragerea ADN-ului mitocondrial dintr-un specimen de A. d. guildayi pentru a clarifica relația sa cu canidele de astăzi, dar proiectul a eșuat, deoarece s-a dovedit imposibil să scoată gudronul de pe oase. [33] Încercarea de a extrage ADN dintr-un mamut columbian a permis cercetătorilor să înțeleagă că componentele organice ale asfaltului pătrund în oase, făcând astfel imposibilă colectarea materialului genetic. [34]
În 2020, o echipă de oameni de știință a analizat cinci probe de ADN datând de 50.000-12-900 de ani, comparându-le cu cele a opt canide din zilele noastre. Rezultatele au arătat că enocionul s-a despărțit de alte canide de lupin în urmă cu aproximativ 5,7 milioane de ani și că nu s-a încrucișat niciodată cu lupii și coioții cu care a coexistat timp de cel puțin 10.000 de ani. Deoarece arborele filogenetic rezultat a indicat că enocionul era mai bazal decât canidele lupinului analizate, s-a propus redenumirea speciei cu vechiul nume generic Aenocyon . [35]
Cladograma de bază, bazată pe date genomice, se bazează pe filogenia detectată de studiul Lindblad-Toh din 2005,[36] modificat pentru a încorpora descoperirile recente despre genul Canis , [37] [38] clasificarea șacalilor africani ca membri ai genul Lupulella , [39] și restabilirea genului Aenocyon . [35]
| Canidele lupinului |
| ||||||||||||||||||||||||||||||
Descriere
Dimensiuni
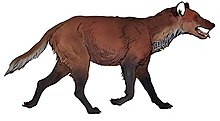
Oenocione a fost considerată odată cel mai mare membru al genului Canis (care include lupi, câini, coioți și șacali aurii, printre altele), [20] dar un studiu genetic făcut în 2021 a arătat că face parte din genul propriu, Aenocyon , și că asemănările cu lupii reali au fost rezultatul evoluției convergente . [35]
Un studiu privind lungimea și circumferința femurilor lui A. d. guildayi și A. d. dirus a indicat că, în viață, greutatea medie a primei trebuia să fie de 60 de kilograme, față de 68 de kilograme din a doua. [18] Un studiu suplimentar al rezistenței humerusului a arătat că greutatea maximă ar putea ajunge la 110 kilograme. [40]
Proporțiile membrelor lui A. d. dirus au fost comparabile cu cele ale lupului, în timp ce cele ale lui A. d. guildayi erau mai scurți. [41] [42] Rămășițele scheletice complete ale enocitelor masculine sunt ușor de identificat, deoarece au un os al penisului distinct de cel al altor canide. [30] [41]
Craniul și dinții
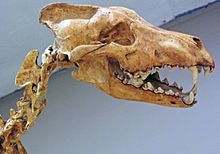
Un studiu al dentiției lui A. d. guildayi a dovedit că era un canid mai specializat decât lupii nord-americani. [27] Mușcătura sa pe dinții canini a fost proporțional cea mai mare dintre canide, depășind cea a licaonei , a lupului gri , a dhole și a dingo . S-a estimat atunci că mușcătura de pe carnassiale era superioară celei a canidelor de astăzi. Deoarece forța mușcăturii în canide este proporțională cu mărimea prăzii, superioritatea enocionului din acest punct de vedere pare să demonstreze că s-a hrănit cu pradă mare. [43]
Un studiu al măsurătorilor craniene și al musculaturii mandibulare a enocionului a arătat că nu au existat diferențe notabile față de lupul cenușiu. Dentiția superioară a fost aceeași cu cea a lupului gri, cu excepția dimensiunilor mai mari, iar al patrulea premolar al enocionului a fost mai mare și mai ascuțit. Maxilarul enocionului a fost prevăzut cu un mușchi temporal mai mare, care i-a permis să muște cu o forță mai mare decât lupul gri. Premolarii inferiori au fost în mod similar mai mari decât cei ai lupului gri [44], iar primul molar a fost chiar mai ascuțit. [8] [26] [44] Dinții săi canini erau mai puternici decât cei ai oricărui canid modern de dimensiuni similare. [45] Aceste diferențe indică faptul că mușcătura enocionului era mai puternică decât cea a lupului cenușiu și că era mai potrivită pentru reținerea prăzilor mari. [44] [46]
Ecologie
Comportament

Având în vedere lipsa unui dimorfism sexual marcat în enocion, se presupune că a format perechi monogame, la fel ca și lupul cenușiu. [47] Mărimea și dentiția hipercarnivoră indică faptul că a mâncat pradă mare și, prin urmare, ar fi trebuit să trăiască în haite. [47][48] [49] Exemplarele multiple găsite în gropile de gudron La Brea par să confirme ipoteza. [50] [51] Din moment ce mamiferele carnivore sociale tind să mănânce pradă la fel de grea ca și masa totală a prădătorilor care le atacă, [52] [53] o turmă de oenociones ar fi fost capabilă să omoare prada cu greutatea de 300-600 de kilograme, cum ar fi bizoni .[48] [49] [54] Deși unii savanți au propus că enocionul a fost un scuturator, bazat pe spargerea frecventă a dinților în exemplarele fosile, gama sa largă și membrele subțiri indică faptul că a fost un prădător activ. La fel ca lupul, el ar folosi dinții post-carnassiali pentru a rupe oasele pentru a extrage măduva. [54]
Dietă

Analiza izotopilor a fost utilizată pentru a identifica anumite elemente chimice care permit cercetătorilor să facă inferențe despre dieta faunei găsite în gropile de gudron. Un studiu efectuat asupra colagenului extras din probele găsite în exemplarele La Brea ar demonstra că enocionul a concurat cu Smilodon fatalis și leul american pentru aceeași pradă, care a inclus pseudocamelul , bizonul Pleistocen , antilocapra pitică , vestul cal și leneșul de pe pământ . Rămășițele mamut și mastodont columbian sunt relativ rare în jurul orașului La Brea. [55]
Studiile ulterioare ale izotopilor găsiți în fosilele de oenocion, vechi de 10.000 de ani, arată că prada lui preferată era calul, în timp ce leneșul, mastodontul, bizonul și cămila erau mai rare. [46] [56] Aceasta arată că enocionul nu s-a specializat în predarea unei singure pradă, ci că a vânat sau s-a hrănit cu carcasele oricărui erbivor disponibil la sfârșitul Pleistocenului târziu. [56]
Concurenți
Înainte de apariția enocionului , America de Nord a fost invadată de xenocion , un mare canid hipercarnivor, strămoș al câinilor sălbatici și cuonului de astăzi . Această specie a fost întotdeauna rară și este probabil că nu ar putea supraviețui competiției cu enocionul. [57]
Analizele stabile ale izotopilor oferă dovezi că enocionul a mâncat aceeași pradă ca Smilodon și leul american . [55] [58] Alți concurenți contemporani au inclus Arctodus simus , puma , coiotul și lupul gri . [58]
După colonizarea Asiei de Est, oenocionul ar întâlni probabil hiena rupestră , cel mai comun și dominant prădător din zonă. Este probabil că concurența cu hiena a însemnat că enocionul a fost întotdeauna rar în Asia. [59]
Zona și habitatul
Resturile de enochion au fost descoperite într-o gamă largă de habitate, inclusiv câmpii, câmpuri și zone montane împădurite din America de Nord, savane din America de Sud și stepe din Asia de Est. Descoperirea rămășițelor în aceste zone indică faptul că enocionul era un animal din câmpiile deschise. [14]
În Statele Unite, au fost raportate fosile de enocione în Arizona, California, Florida, Idaho, Indiana, Kansas, Kentucky, Missouri, Nebraska, New Mexico, Oregon, Pennsylvania, Carolina de Sud, South Dakota, Texas, Utah, Virginia, Virginia de Vest , Wyoming, [30] și Nevada. [60] Identitatea fosilelor raportate mai la nord de California nu este confirmată. [32] În Mexic, rămășițele speciei au fost raportate în El Cedazo din Aguascalientes , Comondú în Baja California Sur , El Cedral din San Luis Potosi , cariera El Tajo lângă Tequixquiac , statul Mexic , Valsequillo în Puebla , Lago din Chapala în Jalisco , cariera Loltun din Yucatán , Potrecito în Sinaloa , cariera San Josecito lângă Aramberri în Nuevo León și Térapa în Sonora . [46]
În America de Sud, rămășițe de enocione au fost raportate doar în statul Falcón din Venezuela , provincia Talara din Peru și în Departamentul Tarija din Bolivia și niciuna dintre ele nu datează de peste 17.000 de ani. Dacă speciile ar fi de origine nord-americană, este probabil că a colonizat America de Sud prin coridorul andin, un pod natural propus ca pasaj pentru mai multe mamifere care au migrat din America Centrală. [27] [30]
În Pleistocenul superior, enocionul a reușit să traverseze Podul Bering pentru a coloniza Eurasia, așa cum a demonstrat în 2020 descoperirea unei maxilare fosilizate găsite într-o mină lângă Harbin, în nord-estul Chinei. Această descoperire a respins ipotezele anterioare conform cărora temperaturile scăzute erau o barieră în calea enocionului, deoarece specia ar trebui să traverseze calotele de gheață la paralela 42 nord pentru a ajunge la pod. [59]
Extincţie
Declinul enocionului a început în urmă cu aproximativ 12.700 de ani, [61] [62] probabil după dispariția prăzii sale mari. [30] [63] [64][65] Este probabil ca enocionul să se fi găsit, așadar, în dezavantaj față de lupul gri mai fragil și mai adaptabil. [66] În plus, spre deosebire de lup, care s-ar fi putut salva de la dispariție prin încrucișarea cu alți membri ai genului său, cum ar fi coioții și câinii, lipsa altor specii strâns legate de enocion a împiedicat-o pe aceasta din urmă să-și răspândească genele. absența semenilor. [35] Cele mai recente rămășițe geologice au fost găsite din peștera Brynjulfson din județul Boone, Missouri [26] [66] și La Mirada din California , datând de 9.440 și respectiv 10.690 de ani. [66]
Notă
- ^ Viața în preistorie - De la originile Pământului la Homo Sapiens , Hobby & Work.
- ^ Muzeul paginii, vizualizați colecțiile de la Rancho La Brea , pe Muzeul de istorie naturală din fundația județului Los Angeles . Adus la 19 decembrie 2016 (arhivat din original la 25 ianuarie 2017) .
- ^ Canis dirus listare completă , pe Fossilworks , Universitatea Macquarie, Australia. Adus pe 19 decembrie 2016 .
- ^ J. Leidy, Notificare a unor oase fosile descoperite de domnul Francis A. Lincke, pe malurile râului Ohio, Indiana , în Proceedings: Academy of Natural Sciences din Philadelphia , n. 7, 1854, p. 200.
- ^ J. Leidy, Observație despre rămășițe de vertebrate dispărute, din Valea râului Niobrara, colectate în timpul Expediției de explorare din 1857, în Nebraska, sub comanda Lieut. GK Warren, Top SUA. Eng., De Dr. FV Hayden, Geolog la Expediție, Proceedings , în Academia de Științe Naturale din Philadelphia , vol. 10, 1858, p. 21.
- ^ J. Leidy, Fauna de mamifere dispărute din Dakota și Nebraska, incluzând o relatare a unor forme aliate din alte localități, împreună cu un sinopsis al rămășițelor de mamifere din America de Nord , în Jurnalul Academiei de Științe Naturale din Philadelphia , vol. 7, 1869, p. 368.
- ^ JA Allen, Descrierea unor rămășițe ale unei specii de lup dispărute și a unei specii de cerbi dispăruți din regiunea de plumb din partea superioară a Mississippi , în American Journal of Science , s3-11, n. 61, 1876, pp. 47-51, DOI : 10.2475 / ajs.s3-11.61.47 .
- ^ a b JC Merriam, Fauna din Rancho La Brea, Partea II. Canide. ( TXT ), în Memoriile Universității din California , vol. 1, 1912, pp. 217-273.
- ^ EL Troxell, Fosile de vertebrate din Rock Creek, Texas , în American Journal of Science , vol. 189, 1915, pp. 613-618.
- ^ JC Merriam, Notă privind poziția sistematică a lupilor din grupul Canis dirus , în Buletinul Departamentului de Geologie al Universității din California , vol. 10, 1918, p. 533.
- ^ Marc Stevenson, 9, în Roberta LS Hall (ed.), Wolf and Man: Evolution in Parallel , New York, Academic Press Inc, 1978, p. 180, ISBN 978-0-12-319250-9 .
- ^ a b EL Lundelius și CJ Bell, 7 The Blancan, Irvingtonian, and Rancholabrean Mammal Ages ( PDF ), în Michael O. Woodburne (ed.), Cretaceu târziu și Cenozoic Mammals of North America: Biostratigraphy and Geochronology , Columbia University Press, 2004, p. 285, ISBN 978-0-231-13040-0 . Accesat la 9 martie 2017 (Arhivat din original la 8 iunie 2017) .
- ^ Lundelius 1972 , Fossilworks , Universitatea Macquarie, Australia. Adus pe 19 decembrie 2016 .
- ^ a b c d Ronald M. Nowak, North American Quaternary Canis , 6, Monografia Muzeului de Istorie Naturală, Universitatea din Kansas, 1979, p. 106, DOI : 10.5962 / bhl.title.4072 , ISBN 978-0-89338-007-6 .
- ^ a b c B. Kurten, Geographic differentiation in the Rancholabrean dire wolf (Canis dirus Leidy) in North America , in HH Genoways and MR Dawson (eds), Contributions in Quaternary Vertebrate Paleontology: A Volume in Memorial to John E. Guilday , Publicație specială 8, Muzeul de istorie naturală Carnegie, 1984, pp. 218 -227, ISBN 978-0-935868-07-4 .
- ^ X. Wang, pleistocenul rău lup rămâne din râul Kansas, cu note despre lupi groaznici în Kansas , în Occasional Papers of the Museum of Natural History, University of Kansas , vol. 137, 1990, pp. 1-7.
- ^ RG Dundas, The dispise of the Late Pleistocene Dire Wolf (Canis dirus): Un model pentru evaluarea extincțiilor carnivore (disertație de doctorat) , Universitatea din California la Berkeley), 1994, pp. 1-491.
- ^ a b c William Anyonge și Chris Roman, [209: NBMEFC 2.0.CO; 2 Noi estimări de masă corporală pentru Canis dirus, lupul dispărut al Pleistocenului ], în Journal of Vertebrate Paleontology , vol. 26, 2006, pp. 209-212, DOI : 10.1671 / 0272-4634 (2006) 26 [209: NBMEFC] 2.0.CO; 2 .
- ^ a b c d e f g h i Richard H. Tedford, Xiaoming Wang și Beryl E. Taylor, Filogenetic Systematics of the North American Fossil Caninae (Carnivora: Canidae) ( PDF ), în Buletinul Muzeului American de Istorie Naturală , vol. 325, 2009, pp. 1-218, DOI : 10.1206 / 574.1 .
- ^ a b Xiaoming Wang și Richard H. Tedford, Dogs: Their Fossil Relatives and Evolutionary History , Columbia University Press, New York, 2008, pp. 1-232, ISBN 978-0-231-13529-0 .
- ^ Canis ambrusteri , despre Fossilworks , Universitatea Macquarie, Australia. Adus la 11 iulie 2016 .
- ^ Ronald M. Nowak și Nicholas E. (Brusco) Federoff, Statutul sistematic al lupului italian Canis lupus , în Acta Theriologica , vol. 47, nr. 3, 2002, pp. 333-338, DOI : 10.1007 / BF03194151 .
- ^ RM Nowak, 9-Wolf evolution and taxonomy , în L. David Mech și Luigi Boitani (eds), Wolves: Behavior, Ecology and Conservation , University of Chicago Press, 2003, ISBN 978-0-226-51696-7 .
- ^ RA Martin și SD Webb,mamifere din Pleistocenul târziu din fauna Devil's Den, județul Levy , în SD Webb (eds), Mamifere din Pleistocen din Florida , Gainesville, University Presses din Florida, 1974, pp. 114 –145, ISBN 978-0-8130-0361-0 .
- ^ GD Goulet, Comparația variației craniului temporal și geografic între lupii gri nearctici, moderni, holoceni și tardivi ai pleistocenului (Canis lupus) și Canis selectat (teza de masterat) , Universitatea din Manitoba, Winnipeg, 1993, pp. 1-116.
- ^ a b c d B. Kurten și E. Anderson, mamifere pleistocene din America de Nord , Columbia University Press, New York, 1980, pp. 1-442, ISBN 978-0-231-03733-4 .
- ^ a b c d A. Berta, Evoluția cuaternară și biogeografia marilor canide sud-americane (Mammalia: Carnivora) , în University of California Publications in Geological Sciences , n. 132, 1988, pp. 1-49.
- ^ a b Francisco J. Prevosti, Filogenia marilor canide sud-americane dispărute (Mammalia, Carnivora, Canidae) folosind o abordare „dovadă totală” , în Cladistics , vol. 26, n. 5, 2010, pp. 456-481, DOI : 10.1111 / j.1096-0031.2009.00298.x .
- ^ C. Frick, fauna înghețată din Alaska , în Istoria naturală , n. 30, 1930, pp. 71-80.
- ^ a b c d e f g RG Dundas, Înregistrări cuaternare ale lupului teribil , Canis dirus , în America de Nord și de Sud ( PDF ), în Boreas , vol. 28, nr. 3, 1999, pp. 375-385, DOI : 10.1111 / j.1502-3885.1999.tb00227.x .
- ^ a b JI Mead, C. Manganaro, CA Repenning and LD Agenbroad, Early Rancholabrean mammifers from Salamander Cave, Black Hills, South Dakota , in KM Stewart and KL Seymour (eds), Palaeoecology and Palaeoenvironments of Cenozoic Late Mamifere, Tributes to the Cariera lui CS (Rufus) Churcher , University of Toronto Press, Toronto, Ontario, Canada, 1996, pp. 458-482, ISBN 978-0-8020-0728-5 .
- ^ a b Julie A. Meachen, Alexandria L. Brannick și Trent J. Fry,morfotipul lupului beringian dispărut găsit în SUA continentală Are implicații pentru migrația și evoluția lupului , în Ecology and Evolution , vol. 6, nr. 10, 2016, p. 3430, DOI : 10.1002 / ece3.2141 , PMC 4870223 , PMID 27252837 .
- ^ DN Janczewski, N Yuhki, DA Gilbert, GT Jefferson și SJ O'Brien, Inferență filogenetică moleculară din fosile de pisici cu dinți de sabie din Rancho La Brea ( PDF ), în Proceedings of the National Academy of Sciences din Statele Unite ale Americii , vol. 89, nr. 20, 1992, pp. 9769-73, cod bib : 1992PNAS ... 89.9769J , DOI : 10.1073 / pnas.89.20.9769 , PMC 50214 , PMID 1409696 .
- ^ David A. Gold, Jacqueline Robinson, Aisling B. Farrell, John M. Harris, Olaf Thalmann și David K. Jacobs,tentativă de extracție a ADN-ului dintr-un mamut columbian Rancho La Brea (Mammuthus columbi): Perspective pentru ADN-ul antic din depozitele de asfalt , în Ecologie și evoluție , vol. 4, nr. 4, 2014, pp. 329-36, DOI : 10.1002 / ece3.928 , PMC 3936381 , PMID 24634719 .
- ^ a b c d Perri, Angela R.; Mitchell, Kieren J.; Mouton, Alice; Álvarez-Carretero, Sandra; Hulme-Beaman, Ardern; Haile, James; Jamieson, Alexandra; Meachen, Julie; Lin, Audrey T.; Schubert, Blaine W.; Ameen, Carly; Antipina, Ekaterina E.; Bover, Pere; Brace, Selina; Carmagnini, Alberto; Carøe, Christian; Samaniego Castruita, Jose A.; Chatters, James C.; Dobney, Keith; Dos Reis, Mario; Evin, Allowen; Gaubert, Philippe; Gopalakrishnan, Shyam; Gower, Graham; Heiniger, Holly; Helgen, Kristofer M.; Kapp, Josh; Kosintsev, Pavel A.; Linderholm, Anna; Ozga, Andrew T.; Presslee, Samantha; Salis, Alexander T.; Saremi, Nedda F.; Shew, Colin; Skerry, Katherine; Taranenko, Dmitry E.; Thompson, Mary; Sablin, Mikhail V.; Kuzmin, Yaroslav V.; Collins, Matthew J.; Sinding, Mikkel-Holger S.; Gilbert, M. Thomas P.; Stone, Anne C.; Shapiro, Beth; Van Valkenburgh, Blaire; Wayne, Robert K.; Larson, Greger; Cooper, Alan; Frantz, Laurent AF (2021). "Dire wolves were the last of an ancient New World canid lineage" . Nature . doi:10.1038/s41586-020-03082-x.
- ^ ( EN ) K. Lindblad-Toh, CM Wade, TS Mikkelsen, EK Karlsson, DB Jaffe, M. Kamal, M. Clamp, JL Chang, EJ Kulbokas, MC Zody, E. Mauceli, X. Xie, M. Breen, RK Wayne, EA Ostrander, CP Ponting, F. Galibert, DR Smith, PJ Dejong, E. Kirkness, P. Alvarez, T. Biagi, W. Brockman, J. Butler, CW Chin, A. Cook, J. Cuff, MJ Daly, D. Decaprio e S. Gnerre, Genome sequence, comparative analysis and haplotype structure of the domestic dog, in Nature, vol. 438, nº 7069, 2005, pp. 803–819, Bibcode:2005Natur.438..803L, DOI:10.1038/nature04338, PMID 16341006 .
- ^ ( EN ) Klaus-Peter Koepfli, John Pollinger, Raquel Godinho, Jacqueline Robinson, Amanda Lea, Sarah Hendricks, Rena M. Schweizer, Olaf Thalmann, Pedro Silva, Zhenxin Fan, Andrey A. Yurchenko, Pavel Dobrynin, Alexey Makunin, James A. Cahill, Beth Shapiro, Francisco Álvares, José C. Brito, Eli Geffen, Jennifer A. Leonard, Kristofer M. Helgen, Warren E. Johnson, Stephen J. o'Brien, Blaire Van Valkenburgh e Robert K. Wayne, Genome-wide Evidence Reveals that African and Eurasian Golden Jackals Are Distinct Species , in Current Biology , vol. 25, n. 16, 2015, p. 2158, DOI : 10.1016/j.cub.2015.06.060 , PMID 26234211 .
- ^ ( EN ) Lamjed Amri, Saverio Bartolini Lucenti, Moncef Saïd Mtimet, Narjess Karoui-Yaakoub, Sergio Ros-Montoya, Maria-Patrocinio Espigares, Mabrouk Boughdiri, Nebiha Bel Haj Ali e Bienvenido Martínez-Navarro, Canis othmanii sp. nov. (Carnivora, Canidae) from the early Middle Pleistocene site of Wadi Sarrat (Tunisia) , in Comptes Rendus Palevol , vol. 16, n. 7, 2017, p. 774, DOI : 10.1016/j.crpv.2017.05.004 .
- ^ Alvares, Francisco; Bogdanowicz, Wieslaw; Campbell, Liz AD; Godinho, Rachel; Hatlauf, Jennifer; Jhala, Yadvendradev V.; Kitchener, Andrew C.; Koepfli, Klaus-Peter; Krofel, Miha; Moehlman, Patricia D.; Senn, Helen; Sillero-Zubiri, Claudio; Viranta, Suvi; Werhahn, Geraldine (2019). "Old World Canis spp. with taxonomic ambiguity: Workshop conclusions and recommendations. CIBIO. Vairão, Portugal, 28th - 30th May 2019" (PDF). IUCN/SSC Canid Specialist Group. Retrieved 6 March 2020.
- ^ Boris Sorkin, A biomechanical constraint on body mass in terrestrial mammalian predators , in Lethaia , vol. 41, n. 4, 2008, pp. 333-347, DOI : 10.1111/j.1502-3931.2007.00091.x .
- ^ a b Adam Hartstone-Rose, Robert G. Dundas, Bryttin Boyde, Ryan C. Long, Aisling B. Farrell e Christopher A. Shaw, The Bacula of Rancho La Brea [ collegamento interrotto ] , a cura di John M. Harris, Science Series 42, Contributions in Science (A special volume entitled La Brea and Beyond: the Paleontology of Asphalt-Preserved Biotas in commemoration of the 100th anniversary of the Natural History Museum of Los Angeles County's excavations at Rancho La Brea), Natural History Museum of Los Angeles County, 15 settembre 2015, pp. 53-63.
- ^ C. Stock e JF Lance, The relative lengths of limb elements in Canis dirus , in Bulletin of the Southern California Academy of Sciences , vol. 47, 1948, pp. 79-84.
- ^ S. Wroe, C. McHenry e J. Thomason,Bite club: Comparative bite force in big biting mammals and the prediction of predatory behaviour in fossil taxa , in Proceedings of the Royal Society B: Biological Sciences , vol. 272, n. 1563, 2005, pp. 619-25, DOI : 10.1098/rspb.2004.2986 , PMC 1564077 , PMID 15817436 .
- ^ a b c W. Anyonge e A. Baker, Craniofacial morphology and feeding behavior in Canis dirus, the extinct Pleistocene dire wolf , in Journal of Zoology , vol. 269, n. 3, 2006, pp. 309-316, DOI : 10.1111/j.1469-7998.2006.00043.x .
- ^ B. Van Valkenburgh e CB Ruff,Canine tooth strength and killing behaviour in large carnivores , in Journal of Zoology , vol. 212, n. 3, 1987, pp. 379-397, DOI : 10.1111/j.1469-7998.1987.tb02910.x .
- ^ a b c John-Paul Hodnett, Mead Jim e A. Baez, Dire Wolf, Canis dirus (Mammalia; Carnivora; Canidae), from the Late Pleistocene (Rancholabrean) of East-Central Sonora, Mexico , in The Southwestern Naturalist , vol. 54.1, marzo 2009, pp. 74-81, DOI : 10.1894/CLG-12.1 .
- ^ a b Van Valkenburgh, Blaire; Sacco, Tyson (2002). "Sexual dimorphism, social behavior, and intrasexual competition in large Pleistocene carnivorans". Journal of Vertebrate Paleontology. 22: 164–169. doi:10.1671/0272-4634(2002)022[0164:SDSBAI]2.0.CO;2
- ^ a b Van Valkenburgh, B.; Koepfli, K.-P. (1993). "Cranial and dental adaptations to predation in canids". In Dunston, N.; Gorman, JL (editori). Mammals as Predators. Oxford University Press, Oxford. pp. 15–37.
- ^ a b Van Valkenburgh, B. (1998). "The decline of North American predators during the late Pleistocene". In Saunders, JJ; Styles, BW; Baryshnikov, GF (eds.). Quaternary Paleozoology in the Northern Hemisphere. Illinois State Museum Scientific Papers, Springfield. pp. 357–374. ISBN 978-0-89792-156-5 .
- ^ McHorse, Brianna K.; Orcutt, John D.; Davis, Edward B. (2012). "The carnivoran fauna of Rancho La Brea: Average or aberrant?". Palaeogeography, Palaeoclimatology, Palaeoecology. 329–330: 118–123. Bibcode:2012PPP...329..118M. doi:10.1016/j.palaeo.2012.02.022
- ^ Carbone, C.; Maddox, T.; Funston, PJ; Mills, MGL; Grether, GF; Van Valkenburgh, B. (2009). "Parallels between playbacks and Pleistocene tar seeps suggest sociality in an extinct sabretooth cat, Smilodon". Biology Letters. 5 (1): 81–5. doi:10.1098/rsbl.2008.0526. PMC 2657756. PMID 18957359
- ^ Sorkin, Boris (2008). "A biomechanical constraint on body mass in terrestrial mammalian predators". Lethaia. 41 (4): 333–347. doi:10.1111/j.1502-3931.2007.00091.x.
- ^ Earle, M. (1987). "A flexible body mass in social carnivores". The American Naturalist. 129 (5): 755–760. doi:10.1086/284670. S2CID 85236511
- ^ a b Anyonge, William; Roman, Chris (2006). "New body mass estimates for Canis dirus, the extinct Pleistocene dire wolf". Journal of Vertebrate Paleontology. 26: 209–212. doi:10.1671/0272-4634(2006)26[209:NBMEFC]2.0.CO;2
- ^ a b Joan Brenner Coltrain, John M. Harris, Thure E. Cerling, James R. Ehleringer, Maria-Denise Dearing, Joy Ward e Julie Allen, Rancho La Brea stable isotope biogeochemistry and its implications for the palaeoecology of late Pleistocene, coastal southern California , in Palaeogeography, Palaeoclimatology, Palaeoecology , vol. 205, n. 3-4, 2004, pp. 199-219, DOI : 10.1016/j.palaeo.2003.12.008 .
- ^ a b K. Fox-Dobbs, JK Bump, RO Peterson, DL Fox e PL Koch, Carnivore-specific stable isotope variables and variation in the foraging ecology of modern and ancient wolf populations: Case studies from Isle Royale, Minnesota, and La Brea ( PDF ), in Canadian Journal of Zoology , vol. 85, n. 4, 2007, pp. 458-471, DOI : 10.1139/Z07-018 .
- ^ Wang, Xiaoming; Tedford, Richard H. (2008). Dogs: Their Fossil Relatives and Evolutionary History. Columbia University Press, New York. p. 60. ISBN 978-0-231-13529-0 .
- ^ a b DeSantis, LRG; Schubert, BW; Schmitt-Linville, E.; Ungar, P.; Donohue, S.; Haupt, RJ (September 15, 2015). John M. Harris (ed.). "Dental microwear textures of carnivorans from the La Brea Tar Pits, California and potential extinction implications". Contributions in Science (A Special Volume Entitled la Brea and Beyond: The Paleontology of Asphalt-Preserved Biotas in Commemoration of the 100th Anniversary of the Natural History Museum of Los Angeles County's Excavations at Rancho la Brea). Science Series 42. Natural History Museum of Los Angeles County: 37–52.
- ^ a b Dan, Lu; Yang, Yangheshan; Lia, Qiang; Nia, Xijun (1 October 2020). "A late Pleistocene fossil from Northeastern China is the first record of the dire wolf (Carnivora: Canis dirus) in Eurasia". Quaternary International. doi:10.1016/j.quaint.2020.09.054
- ^ Eric Scott e Kathleen B. Springer,First records of Canis dirus and Smilodon fatalis from the late Pleistocene Tule Springs local fauna, upper Las Vegas Wash, Nevada , in PeerJ , vol. 4, 2016, pp. e2151, DOI : 10.7717/peerj.2151 , PMC 4924133 , PMID 27366649 .
- ^ FR O'Keefe, EV Fet e JM Harris, Compilation, calibration, and synthesis of faunal and floral radiocarbon dates, Rancho La Brea, California ( PDF ), in Contributions in Science , vol. 518, 2009, pp. 1-16. URL consultato il 28 giugno 2017 (archiviato dall' url originale il 3 luglio 2017) . Archiviato il 3 luglio 2017 in Internet Archive .
- ^ F.Robin O'Keefe, Wendy J. Binder, Stephen R. Frost, Rudyard W. Sadlier e Blaire Van Valkenburgh, Cranial morphometrics of the dire wolf, Canis dirus, at Rancho La Brea: temporal variability and its links to nutrient stress and climate , in Palaeontologia Electronica , vol. 17, n. 1, 2014, pp. 1-24.
- ^ RW Graham e JI Mead, Environmental fluctuations and evolution of mammalian faunas during the last deglaciation in North America , in WF Ruddiman e HE Wright (a cura di), North America and Adjacent Oceans During the Last Deglaciation , Geological Society of America K-3, Boulder, Colorado, 1987, pp. 371-402, ISBN 978-0-8137-5203-7 .
- ^ AD Barnosky, The Late Pleistocene extinction event as a paradigm for widespread mammal extinction , in Stephen K. Donovan (a cura di), Mass Extinctions: Processes and Evidence , Columbia University Press , New York, 1989, pp. 235-255, ISBN 978-0-231-07091-1 .
- ^ Blaire Van Valkenburgh e Fritz Hertel, Tough Times at La Brea: Tooth Breakage in Large Carnivores of the Late Pleistocene ( PDF ), in Science , New Series, vol. 261, n. 5120, 1993, pp. 456-459, Bibcode : 1993Sci...261..456V , DOI : 10.1126/science.261.5120.456 , PMID 17770024 .
- ^ a b c Elaine Anderson, Chapter 2: Who's who in the Pleistocene , in Paul S. Martin e Richard G. Klein (a cura di), Quaternary Extinctions: A Prehistoric Revolution , University of Arizona Press , Tucson, 1984, p. 55, ISBN 978-0-8165-1100-6 .
Altri progetti
-
 Wikimedia Commons contiene immagini o altri file su Canis dirus
Wikimedia Commons contiene immagini o altri file su Canis dirus -
 Wikispecies contiene informazioni su Canis dirus
Wikispecies contiene informazioni su Canis dirus
Collegamenti esterni
- ( EN ) Aenocyon dirus , su Enciclopedia Britannica , Encyclopædia Britannica, Inc.
- ( EN ) Aenocyon dirus , su Fossilworks.org .
| Controllo di autorità | LCCN ( EN ) sh2002004650 |
|---|