Psittacosaurus
| Psittacosaurus | |
|---|---|
 | |
| Starea de conservare | |
Fosil | |
| Clasificare științifică | |
| Domeniu | Eukaryota |
| Regatul | Animalia |
| Phylum | Chordata |
| Superordine | Dinozauria |
| Ordin | † Ornitischia |
| Subordine | † Marginocefalia |
| Familie | † Psittacosauridae Osborn, 1923 |
| Tip | † Psittacosaurus Osborn, 1923 |
| Nomenclatura binominala | |
| † Psittacosaurus mongoliensis Osborn , 1923 | |
| Sinonime | |
| |
| Specii | |
| |
Psittacosaurus (al cărui nume înseamnă „ șopârlă de papagal ”) este un gen dispărut de dinozaur ceratopsid , care a trăit în perioada Cretacicului timpuriu între aproximativ 123,2 și 100 de milioane de ani , în ceea ce este astăzi Asia . Este unul dintre cei mai cunoscuți și mai cunoscuți dinozauri de către paleontologi datorită cantității excepționale de rămășițe fosile, care i-au permis acestora din urmă să clasifice 11 specii diferite, făcându-l unul dintre cele mai bogate genuri din lume, cu fosile din Mongolia , Siberia. , China și poate chiar Thailanda .
La vârsta adultă, aceste animale erau legate de bipede și craniul lor era împodobit cu pinteni de os și un cioc puternic și curbat. O particularitate a unor specii a fost prezența unor structuri filiforme asemănătoare penelor alungite, similare cu cele găsite în Tianyulong , lângă coadă și pe o parte a spatelui. Având în vedere proporția creierului cu corpul animalului, Psittacosaurusul trebuie să fi avut comportamente complexe, cel mai probabil era un animal care era activ atât ziua cât și noaptea și avea un simț al vederii și al mirosului foarte dezvoltat.
În ciuda aspectului său, Psittacosaurus este unul dintre cele mai vechi ceratopside , animale precum cunoscutul Triceratops , dintre care Psittacosaurus era o rudă mai apropiată, în comparație cu Yinlong . Acest animal împarte familia taxonomică cu Hongshanosaurus , (probabil sinonim cu Psittacosaurus ), împreună cu care este unul dintre primele ceratopside care au evoluat și din care au evoluat forme mai complexe. Animale similare au fost găsite mai ales în Asia, cu excepția Aquilops găsite în America de Nord . Specia P. lujiatunensis ar fi cea mai veche din care ar fi evoluat toate celelalte specii.
Cu peste 400 de exemplare fosile găsite, inclusiv nenumărate schelete complete, Psittacosaurus este unul dintre genurile de dinozauri cel mai bine cunoscute de paleontologi. Fosilele găsite aparțin tuturor claselor de vârstă ale animalului, de la faza de eclozare până la exemplarele adulte, ceea ce le-a permis paleontologilor să studieze creșterea , dezvoltarea și biologia animalului. Abundența de fosile ale acestui animal l-a făcut una dintre cele mai faimoase și apreciate fosile din Asia.
Descriere

Diferitele specii de Psittacosaurus diferă și variază între ele prin mărime și caracteristicile anatomice ale craniului și scheletului, chiar dacă împărtășesc forma generală a corpului. Cea mai cunoscută specie, P. mongoliensis , ar putea ajunge până la 2 metri (6,5 picioare) în lungime. [1] Greutatea maximă a unui animal adult a fost probabil de aproximativ 20 kg (44 lb). [2] Unele specii precum P. lujiatunensis, P. și P. neimongoliensis xinjiangensis [3] [4] [5] aveau dimensiuni similare, în timp ce altele precum P. sinensis și P. meileyingensis erau mai mici. [6] Specia P. ordosensis , în special, a fost cea mai mică dintre toate, cu aproximativ 30% mai puțin decât P. mongoliensis . [4] Cele mai mari specii au fost P. lujiatunensis și P. sibiricus . [7][8]

Craniul lui Psittacosaurus era foarte diferit de cel al altor dinozauri ornitischieni ai timpului său. Craniul era înalt și îndesat, cu un profil aproape rotund la unele specii. Doar porțiunea frontală a orbitei a reprezentat 40% din lungimea totală a craniului, mai scurtă decât orice alt ornitisc cunoscut. Mandibula animalului a fost caracterizată printr-o creastă bulboasă verticală, îndreptată în jos, în centrul fiecărui dinte. Vârful craniului era prevăzut cu un cioc mare și robust foarte pronunțat asemănător cu cel al papagalilor moderni (de unde și numele speciei). La nivelul taliei, ciocul animalului era acoperit cu cheratină , care forma o suprafață perfectă pentru tăierea și ruperea materialului vegetal . Craniul Psittacosaurus a împărtășit multe trăsături comune cu ciocul altor ceratopside, cum ar fi medula rostrală unică la vârful maxilarului superior și proeminențele osoase ale pomeților . Cu toate acestea, fiind încă destul de primitiv, nu există niciun semn nici al gulerului osos, nici al coarnelor prezente în cele mai evoluate forme. [9] Cu toate acestea, o schiță timpurie a coarnelor a fost găsită pe craniul speciei P. sibiricus , dar se crede că acesta este doar un exemplu de evoluție convergentă .[8]
Spre deosebire de craniu, scheletul Psittacosaurus era tipic pentru majoritatea dinozaurilor ornitischieni bipezi . La P. mongoliensis , în mod similar cu alte specii, membrele anterioare aveau doar aproximativ 58% din lungimea membrelor posterioare. Cu toate acestea, structura scheletică a mâinilor indică faptul că animalul nu le-ar putea roti pentru a merge pe patru picioare, așa că erau bipede obligați. [10] Spre deosebire de majoritatea ornitischienilor, mâinile lui Psittacosaurus aveau doar patru degete, comparativ cu cele cinci degete ale majorității celorlalți ornitischieni, în timp ce picioarele din spate aveau patru degete (inclusiv pinteni) ca toți ceilalți ornitischieni . [9]
Integumente și colorare

Structuri de tegument , în aparență asemănătoare penelor de gâscă, au fost adesea găsite printre nenumăratele fosile aparținând Psittacosaurusului . Primul eșantion pe care s-au găsit astfel de structuri a fost exportat ilegal din China în Germania, unde a fost descris în așteptarea repatrierii. Specimenul, expus la Senckenberg Naturmuseum , a prezentat semne de tegumente și structuri filamentoase pe numeroase părți ale corpului. Cu toate acestea, nu s-a stabilit încă cu certitudine cărei specii îi aparține, prin urmare este denumită P. sp. , de către savanții care l-au examinat, în 2002. Din analize a ieșit la iveală faptul că animalul era în principal acoperit cu solzi de formă vag hexagonală, asemănătoare cu impresiile de piele găsite în Chasmosaurus . [11] În partea superioară a cozii aceluiași exemplar, au fost descoperite structuri tegumentare lungi asemănătoare cu perii. Paleontologii au confirmat ulterior că nu este vorba de material vegetal. Părul s-a extins de pe piele, aproape de vertebre, lângă vârful cozii. Toate părurile erau o treime proximală a cozii și erau probabil de formă circulară sau tubulară la nivelul taliei. Sub lumina ultravioletă , plumele străluceau cu aceeași fluorescență ca solzii, sugerând posibilitatea ca acestea să fie keratinizate . În cele din urmă, paleontologii au afirmat că: „În prezent, nu există dovezi că aceste structuri ar putea fi similare sau diferite de filamentele cu pene ale dinozaurilor teropode găsite în formațiunea Yixian”. [11]

În 2008, a fost publicat un nou studiu asupra tegumentelor găsite pe alte două exemplare de Psittacosaurus sp. . Studiile au comparat o secțiune de piele cu cea a altor animale moderne, arătând că straturile dermice ale dinozaurilor au evoluat în paralel cu cele ale altor vertebrate mari. Fibrele de colagen ale țesutului dermic Psittacosaurus erau complexe, practic identice cu cele ale tuturor celorlalte vertebrate, dar aveau o grosime excepțională de aproximativ patruzeci de straturi. Eșantionul de piele a fost găsit în zona abdomenului, astfel încât pielea mai groasă din zona abdomenului ar putea servi drept factor de descurajare împotriva oricărui atac de prădători. [13]
Studiul unui specimen descris în 2016 arată prezența melanosomilor Psittacosaurus , indicând faptul că animalul avea pigmentare de lumină de fundal (burtă deschisă și spate întunecat), indicând faptul că animalul era un locuitor al pădurilor dense cu puțină lumină., Similar cu cerbii moderni și antilopa ; dungile și petele de pe membre pot reprezenta o colorare perturbatoare. Specimenul avea, de asemenea, grupuri dense de pigment pe umeri, față (poate pentru afișare) și în cloacă (care ar fi putut avea o funcție antimicrobiană ), precum și o clapă mare de piele care conecta tarsul la coadă, numit uropathagium . Ochii săi mari indică faptul că animalul avea probabil o vedere excelentă, utilă pentru găsirea hranei și evitarea prădătorilor. [14] [15] [16] Autorii nu au fost în măsură să determine ce specie de Psittacosaurus a aparținut exemplarul găsit în formațiunea Jehol, dar P. mongoliensis este exclus, pe baza caracteristicilor șoldului. [17]
Un alt studiu din 2016 a folosit un laser care induce fluorescența pentru a analiza structura internă a perilor animalului. Părul foarte cornificat este aranjat în grupuri mici de trei până la șase peri individuali, iar fiecare păr a fost umplut cu o pulpă. Autorii consideră ideea că firele au fost un omolog la Tianyulong pene, Beipiaosaurus EBFFs, sau , eventual , pene de păsări. Pere similare au fost găsite și la unele păsări existente, precum și la anhima cornuta ; aceste structuri diferă de pene prin faptul că nu au dezvoltat foliculul, dar ambele derivă din populații de celule discrete. Există, de asemenea, o masă mare de țesuturi moi foarte întunecate lângă coarnele jugale; aceasta reprezintă probabil o înveliș de keratină sau o clapă de piele. [18]
Caracteristicile speciei
Craniile speciei P. mongoliensis sunt plate deasupra, în special pe spatele craniului, cu o depresiune triunghiulară, fenestra antorbitală , prezentă pe suprafața exterioară a maxilarului . O flanșă este prezentă pe marginea inferioară a dentarului (osul pe care sunt ancorați dinții maxilarului inferior), deși nu este la fel de proeminent ca la P. meileyingensis sau P. major (= P. lujiatunensis ). Specia P. mongoliensis este printre cele mai cunoscute specii. Craniul exemplarului de tip (probabil un exemplar juvenil), [4] are 15,2 cm (6 in) lungime, iar femurul asociat are 16,2 cm (6,4 in) lungime. [19] Alte exemplare sunt mai mari, cu cel mai mare femur documentat măsurând aproximativ 21 de centimetri (8,25 țoli). [20]
P. sinensis se distinge ușor de toate celelalte specii prin numeroasele trăsături ale craniului. Craniile adulte sunt mai mici decât cele ale P. mongoliensis și au mai puțini dinți. În mod unic, premaxila contactează osul jugal (obraz) din exteriorul craniului. Creșterile jugalei se împrăștie lateral, formând „ coarne ” proporțional mai mari decât cele ale altor specii de Psittacosaurus , cu excepția P. sibiricus și P. lujiatunensis . Datorită obrajilor extinși, craniul este mai lat decât este lung. Există, de asemenea, un mic „corn” în spatele ochiului , în contact cu osul jugal și postorbital , caracteristică observată și la P. sibiricus . Mandibulei îi lipsesc deschiderile sau fenestra, așa cum se vede la alte specii, iar întreaga maxilară inferioară este înclinată spre exterior, dând gura animalului aspectul unei malocluzii . [21] [22] Craniul unui adult P. sinensis ar putea ajunge la 11,5 centimetri (4,5 inci) în lungime. [4]
P. sibiricus este cea mai mare specie cunoscută de Psittacosaurus . Craniul specimenului tip are o lungime de 20,7 cm (8,25 in), iar femurul are o lungime de 22,3 cm (8,75 in). Specia se distinge și prin gulerul osos mic din jurul gâtului, care este mai lung decât orice altă specie, la 15-18% din lungimea craniului. O caracteristică foarte izbitoare a P. sibiricus este numărul de „coarne” din jurul ochilor, cu trei puncte proeminente pe fiecare postorbital și unul în fața fiecărui ochi, pe oasele palpebrale . Coarnele similare cu cele găsite pe P. sinensis postorbital nu sunt la fel de pronunțate, dar pot fi omoloage . Jugala are „coarne” extrem de proeminente și ar putea contacta și premaxila, ambele caracteristici care se văd și în posibila corelație a P. sinensis . Există o flanșă pe dentarul maxilarului inferior, similară cu cea a P. mongoliensis , P. meileyingensis și P. sattayaraki . De asemenea, se poate distinge de alte specii de Psittacosaurus printr-o combinație de 32 de trăsături anatomice, dintre care șase sunt unice speciei. Cele mai multe dintre acestea sunt detalii ale craniului, dar o caracteristică neobișnuită este prezența a 23 de vertebre între craniu și pelvis, spre deosebire de 21 sau 22 din celelalte specii cunoscute pentru vertebre.[8]
Specia P. xinjiangensis se distinge printr-un „corn” jugal proeminent aplatizat în față, precum și prin unele caracteristici ale dinților. Ileonul , unul dintre cele trei oase pelvine, are un proces osos foarte lung în spatele cupei . [21] Un femur adult avea o lungime de aproximativ 16 centimetri (6,3 țoli). [5] Specia P. meileyingensis are un gât și un guler mai scurte decât oricare altă specie, făcând craniul aproape de profil circular. Orbita este aproximativ triunghiulară și are o flanșă proeminentă pe marginea inferioară a dentiției, caracteristică observată și la exemplarele P. lujiatunensis și, într-o măsură mai mică, la P. mongoliensis , P. sattayaraki și P. sibiricus .[8] [21] Craniul de tip complet, probabil al unui adult, are o lungime de 13,7 centimetri (5,5 țoli). [6] Dentariul lui P. sattayaraki are o flanșă similară cu cea a lui P. mongoliensis , P. sibiricus , P. lujiatunensis și P. meileyingensis , deși este mai puțin pronunțat la acele specii. Materialul pare a avea aproximativ aceeași dimensiune ca P. sinensis . [23] Osul frontal al P. neimongoliensis este distinct îngust în comparație cu cel al altor specii, rezultând un craniu mai complex. Ischiul bazinului este, de asemenea, mai lung decât femurul , care diferă de alte specii în care sunt cunoscute aceste oase. [21] Specimenul tip are un craniu lung de 13,2 cm și o lungime femurală de 13 cm, deși exemplarul nu era adult. Un P. neimongoliensis adult era probabil mai mic decât P. mongoliensis , cu un craniu și o coadă corespunzător mai lungi. [4] P. ordosensis se poate distinge prin numeroasele caracteristici ale jugalei, care au „coarne” foarte proeminente [21], precum și fiind cea mai mică specie cunoscută. Un craniu adult măsura doar 9,5 centimetri (3,75 in) în lungime. [4]
Craniul tip P. lujiatunensis măsoară 19 cm lungime, în timp ce cel mai cunoscut craniu are 20,5 cm lungime, deci această specie are dimensiuni similare cu P. mongoliensis și P. sibiricus . Această specie are o groapă în fața ochiului, ca la P. mongoliensis . Oasele jugale se extind în lățime, făcând craniul mai lat decât este lung, așa cum se vede în P. sinensis . Astfel de structuri se găsesc și la P. sibiricus . În general, se crede că această specie prezintă mai multe caracteristici primitive în comparație cu alte specii de Psittacosaurus , ceea ce este în concordanță cu vârsta sa geologică superioară. [7] Specia P. gobiensis a fost mai mică (1 metru) și diferă de alte specii de Psittacosaurus prin „detalii semnificative, dar structurale minore.” Acestea includ prezența unui corn piramidal pe postorbital, o depresiune pe postorbital-jugal contact și grosimea smalțului. Specia a fost contemporană cu P. mongoliensis . [24]
Clasificare

Psittacosaurus este genul tip al familiei Psittacosauridae , numit de Osborn în 1923. [25] [26] Psittacosauridele erau animale primitive comparativ cu orice ceratopsid cunoscut, cu excepția Yinlong și, eventual, Chaoyangsauridae . [9] [27] În timp ce Psittacosauridae a fost o ramură timpurie a arborelui genealogic Ceratopsia, Psittacosaurus în sine nu era probabil în mod direct ancestral niciunui alt grup de ceratopsie. Toți ceilalți ceratopsieni au păstrat al cincilea deget, o plesiomorfie sau un accident vascular cerebral primitiv, în timp ce toate speciile de Psittacosaurus aveau doar patru degete pe fiecare mână. Mai mult, fenestra anterorbitală , o deschidere în craniu între ochi și nară , a fost pierdută în timpul evoluției Psittacosauridae, dar este încă prezentă în majoritatea celorlalte ceratopsine și în majoritatea altor arosauri . [9]
În 2014, descriptorii unui nou taxon ceratopsian bazal au publicat o analiză filogenetică care a inclus Psittacosaurus . Cladograma care urmează provine din analizele lor, plasând genul ca fiind unul dintre cei mai primitivi ceratopsieni. Autorii (Farke și colab. ) Au remarcat faptul că toți taxonii din afara Leptoceratopsidae și Coronosauria , cu excepția noului lor gen Aquilops , provin din Asia, ceea ce înseamnă că probabil grupul își are originea acolo. [28]
| Marginocefalia |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

Deși au fost numite numeroase specii de Psittacosaurus , relațiile taxonomice dintre ele nu au fost încă explorate și nu există un consens științific pe această temă. [4] [22] [29] Au fost publicate mai multe analize filogenetice, dar cele mai detaliate sunt cele ale lui Alexander Averianov și colab. (2006)[8] , Hai-LU Lei și colab. (2008) [30] și Paul Sereno (2010). [31] Cladograma de mai jos, arată analiza de Hai-LU Lei și colab. (2008):
| Psittacosaurus |
| ||||||||||||||||||||||||||||||||||||||||||||||||
În 2005, Zhou și colegii săi au sugerat că P. lujiatunensis era cea mai primitivă specie dintre toate, ceea ce ar fi în concordanță cu apariția sa anterioară în dosarul fosil. [7]
Istoria descoperirii
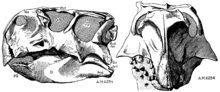
Psittacosaurus a fost descris pentru prima dată ca un gen în 1923 de Henry Fairfield Osborn . El a numit specia tip P. mongoliensis , datorită localizării sale în Mongolia , stabilind, de asemenea, noua familie a Psittacosauridae. Rămășițele acestui dinozaur au fost descoperite cu un an mai devreme, în timpul celei de-a treia expediții a Muzeului American de Istorie Naturală din deșertul Gobi , Mongolia, când unul dintre piloții expediției a găsit specimenul de tip (AMNH 6254), care păstrează un complet, precum și un schelet post-cranian lipsit de secțiuni ale membrelor. [25] Aceeași expediție a dezvăluit rămășițele multor alți dinozauri mongoli celebri, precum Protoceratops , Oviraptor și Velociraptor . [32] Expedițiile comune succesive mongole, ruse, chineze, americane, poloneze, japoneze și canadiene au recuperat mai multe exemplare din toată Mongolia și nordul Chinei. În aceste zone, fosilele P. mongoliensis au fost găsite în majoritatea straturilor sedimentare care datează din Aptian și Albian , din perioada Cretacic inferior , cu aproximativ 125-100 milioane de ani în urmă. În total, au fost recuperate fosile a peste 75 de indivizi, inclusiv aproape 20 de schelete complete de cranii. [9] Aceste fosile au provenit de la indivizi de vârste diferite, de la copii mici de până la 13 centimetri, la adulți și vârstnici care ating aproape 2 metri lungime. [33]

În descrierea Psittacosaurus mongoliensis în 1923, Osborn a atribuit numele Protiguanodon mongoliensis unui alt schelet găsit în apropiere, crezând că este strămoșul ornitopodului Iguanodon , în noua subfamilie Protiguanodontinae. Specimenul mongol Protiguanodon (AMNH 6523) a măsurat 1,35 metri (4,4 ft) și este cunoscut pentru o mare parte a scheletului, deși la momentul descrierii vertebrele din gât erau încă acoperite de matrice. Osborn și-a diagnosticat taxonii pe baza caracteristicilor dinților și botului. [25] Cu toate acestea, taxonomiștii moderni consideră că aceste caracteristici sunt nesemnificative, plasând în schimb Protiguanodon în Psittacosaurus mongoliensis . [25] [31] Când scheletul a fost pregătit în continuare, a devenit clar că era aproape identic cu Psittacosaurus mongoliensis . [19] În 1958, paleontologul chinez Yang Zhongjian (mai cunoscut sub numele de CC Young) a redenumit scheletul Psittacosaurus protiguanodonensis . [34] Astăzi specimenul este denumit în general specia Psittacosaurus mongoliensis și denumirile Protiguanodon mongoliense și Psittacosaurus protiguanodonensis sunt considerate sinonime ale Psittacosaurus mongoliensis , fiind inventate mai întâi. [9] [31]
În 1931, CC Young a numit o nouă specie de Psittacosaurus pe baza unui craniu parțial descoperit în Mongolia Interioară , China. [35] Craniul a fost numit P. osborni , în onoarea lui Henry Fairfield Osborn. Valabilitatea acestei specii este acum considerată echivocă. Sereno (1990) l-a considerat sinonim cu P. mongoliensis , găsit în straturile din apropiere de aceeași vârstă. [22] Dvs. și Dodson (2004) l-ați enumerat ca fiind valid într-un tabel, dar nu în textul lor. [9] Într-o revizuire din 2010, Sereno a considerat din nou P. osborni ca sinonim pentru P. mongoliensis , dar a menționat că a fost o mișcare riscantă din cauza prezenței mai multor specii valide de psittacosauride în Mongolia Interioară. [31] Young a descris, de asemenea, specia P. tingi în același raport din 1931 care conținea P. osborni . Această specie ar avea la bază mai multe fragmente ale craniului. [35] Ulterior, cele două specii au fost sinonime cu numele de P. osborni . [9] Tu și Dodson (2004) ați urmat acest lucru într-un tabel, [9] dar Sereno a considerat că ambele specii sunt sinonime ale P. mongoliensis ; [22] [31] un tabel din acesta din urmă a făcut totuși referire la P. tingi ca la un nomen dubium . [31] Jumătatea anterioară a unui craniu găsită în județul Guyang , Mongolia Interioară, a fost descrisă ca Psittacosaurus guyangensis , în 1983. Resturi postcraniene disjuncte reprezentând mai mulți indivizi au fost găsite în aceeași localitate și au fost atribuite aceleiași specii. [36] Deși diferă de specimenul de tip al P. mongoliensis , acesta se încadrează în sfera variației individuale observate la alte exemplare din specia respectivă și, prin urmare, nu mai este recunoscută ca specie valabilă. [22] Tu și Dodson (2004) ați inclus P. guyangensis într-un tabel de taxoni valabili, dar nu l-ați inclus ca atare în textul lor. [9]
Specii atribuite
De-a lungul anilor, până la șaptesprezece specii au fost numite în genul Psittacosaurus , deși astăzi doar nouă sau unsprezece sunt considerate valide, [3] [21] [31] [37] reprezentând cel mai mare număr de specii valide atribuite în prezent oricărui alt gen dinozaur (cu excepția păsărilor ). Dimpotrivă, de fapt, majoritatea celorlalte genuri de dinozauri sunt monospecifice, adică conțin doar o singură specie, deși această diferență se datorează probabil procesului de fosilizare. În timp ce Psittacosaurus este cunoscut pentru sute de exemplare fosile, majoritatea celorlalte specii de dinozauri sunt cunoscute pentru materiale mult mai fragmentare, iar multe sunt reprezentate de un singur exemplar. Cu atât de multe exemplare disponibile, diversitatea Psittacosaurus poate fi analizată mai cuprinzător decât cea a majorității dinozaurilor, rezultând în recunoașterea mai multor specii. Majoritatea genurilor de animale existente sunt reprezentate de mai multe specii, sugerând că acesta a fost și cazul genurilor de dinozauri, deși majoritatea acestor specii nu au fost păstrate sau nu au fost identificate. Mai mult, majoritatea dinozaurilor sunt cunoscuți exclusiv prin oase și pot fi evaluați doar din punct de vedere morfologic , în timp ce speciile existente au adesea o morfologie scheletică foarte similară, dar diferă în alte moduri care nu ar fi păstrate în fosilizare, cum ar fi comportamentul sau colorarea. Prin urmare, diversitatea eficientă a speciilor poate fi mult mai mare decât cea recunoscută în prezent în acest gen și în alte genuri de dinozauri. [38] Deoarece unele specii sunt cunoscute doar din materiale craniene, speciile de Psittacosaurus se disting în principal prin caracteristicile craniului și ale dinților. Mai multe specii pot fi recunoscute și după caracteristicile bazinului . [5]
În anii 1950, o nouă specie chineză de Psittacosaurus a fost găsită în formațiunea Qingshan din provincia Shandong , la sud-est de Beijing , datând din Aptian-Albian. Paleontologul CC Young a numit specia P. sinensis pentru a o diferenția de P. mongoliensis , găsită inițial în Mongolia. [34] În cadrul formațiunii au fost găsite fosile de peste douăzeci de indivizi, inclusiv câteva cranii și schelete complete, făcând această specie cea mai cunoscută după P. mongoliensis . [9] Paleontologul chinez Zhao Xijin a numit o nouă specie în onoarea mentorului său, CC Young, în 1962. [39] Cu toate acestea, specimenul tip de P. youngi (un schelet parțial și un craniu) a fost descoperit în aceleași roci unde P. sinensis a fost găsit și cei doi par a fi foarte asemănători, astfel încât P. youngi este considerat, în general, un sinonim junior al speciei P. sinensis . [22] [31] Ca și în cazul P. guyangensis și P. osborni , You și Dodson (2004) l-au enumerat ca fiind valid într-un tabel, dar nu și în textul lor. [9]
În 1988, Zhao și paleontologul american Paul Sereno au descris P. xinjiangensis , care își datorează numele regiunii autonome Xinjiang , unde a fost descoperită specia. [40] La începutul anilor 1970, mai mulți indivizi de vârste diferite au fost găsiți și descriși de Sereno și Zhao, deși holotipul și scheletul mai complet aparțineau unui tânăr. Ulterior, un schelet adult a fost descoperit într-o altă locație din Xinjiang. [5] Aceste exemplare provin din partea superioară a grupului Tugulu, care este considerat în mod normal ca datând din Aptian-Albian. [5] O a doua specie a fost descrisă în 1988 de Sereno și Zhao, împreună cu doi colegi chinezi, P. meileyingensis din formațiunea Jiufotang, lângă orașul Meileyingzi, provincia Liaoning , în nord-estul Chinei. Questa specie è conosciuta da quattro crani fossili, di cui uno associato a del materiale scheletrico, ritrovato nel 1973 da alcuni scienziati cinesi. [6] L'età dello Jiufotang a Liaoning è sconosciuta, ma nella vicina provincia della Mongolia Interna, è datata a circa 110 milioni di anni fa, nella fase Albiana del Cretaceo inferiore. [41]

Il paleontologo francese Eric Buffetaut e un collega tailandese, Varavudh Suteethorn, descrissero una mascella superiore e inferiore della Formazione Khok Kruat, della Thailandia nel 1992, dandogli il nome di P. sattayaraki . [23] Nel 2000, Sereno si interrogò sull'effettiva validità di questa specie, citando la sua natura erosa e frammentaria e notò l'assenza di caratteristiche del genere Psittacosaurus . [37] Tuttavia, nel 2002 gli autori originali hanno pubblicato nuove immagini del fossile che sembrano mostrare i denti nella mandibola inferiore che presentano la cresta verticale bulbosa caratteristica degli psittacosauri. [42] Altri autori hanno anche difeso la sua validità,[8] mentre altri continuano a considerarla una specie dubbia. [9] [21] [31] Sereno (2010) ha proposto che la migliore assegnazione per il tipo di materiale possa essere un incertae sedis in Ceratopsia . [31]
Due nuove specie di Psittacosaurus sono state descritte dal paleontologo canadese Dale Russell e Zhao, nel 1996. La prima è stata nominata P. neimongoliensis , ossia il nome cinese mandarino per la Mongolia Interna . La specie si basa su uno scheletro quasi completo, che comprende la maggior parte del cranio, ritrovato nella Formazione Ejinhoro, risalente al Cretaceo inferiore, con altri sette individui. [4] Russell e Zhao hanno anche nominato la specie P. ordosensis , nel 1996, chiamata così in onore alla prefettura di Ordos , della Regione Autonoma della Mongolia Interna. L'esemplare tipo è uno scheletro quasi completo, che include parte del cranio. Tuttavia, solo il cranio, la mascella inferiore ei piedi dell'esemplare sono stati descritti. Altri tre esemplari sono stati riferiti a questa specie ma rimangono ancora non-descritti. Come P. neimongoliensis , questa specie è stata scoperta nella Formazione Eijnhoro. [4] Sereno (2010) ha trovato la specie descritta come indistinguibile da P. sinensis , un'altra piccola specie, ma ha suggerito che uno studio supplementare di P. ordosensis potrebbe rivelare caratteristiche diagnostiche. Pertanto, ha provvisoriamente designato P. ordosensis come nomen dubium . [31]
Xu Xing , un altro paleontologo cinese, ha nominato una nuova specie di Psittacosaurus nel 1997, basata su un cranio completo con vertebre associate e un arto anteriore. Questo materiale è stato recuperato nella Provincia di Gansu , vicino al confine con la Mongolia Interna. Questa specie è chiamata P. mazongshanensis , in onore della vicina montagna chiamata Mazongshan (Mountain Horse Mane) ed è stata descritta in modo preliminare. [29] Purtroppo, il cranio è stato danneggiato durante la preparazione all'Istituto Cinese di Paleontologia dei Vertebrati e Paleoantropologia (IVPP), e diversi frammenti sono andati persi, compresi tutti i denti. [7] I resti sono stati ritrovati nella Formazione Xinminbao, la cui datazione è incerta, anche se ci sono prove che si sia depositata nel tardo Barremiano attraverso le fasi aptiane. [43] Nel 2000, Sereno suggerì che P. mazongshanensis fosse un nomen dubium , senza caratteristiche uniche che lo separavano da qualsiasi altra specie di Psittacosaurus . [37] Tuttavia, gli autori più recenti hanno osservato che può essere distinta per una protuberanza ossea, indirizzata verso l'esterno e il basso, sulla mascella superiore.[8] [7] [31] Altre caratteristiche inizialmente utilizzate per distinguere le specie sono state riconosciute come i risultati della deformazione del cranio dopo la fossilizzazione. [7] Tuttavia, Sereno (2010) non è convinto della sua validità. [31]
A partire dagli anni 50', i paleontologi russi hanno iniziato a ritrovare alcuni esemplari di Psittacosaurus in una località nei pressi del villaggio di Shestakovo, nello oblast' di Kemerovo , in Siberia . Negli anni 90' sono state esplorate altre due località vicine, una delle quali ha restituito diversi scheletri completi. Questa specie è stata nominata P. sibiricus nel 2000 in un documento scientifico scritto da cinque paleontologi russi, ma il merito di questo nome è ufficialmente dato a due di questi autori, Alexei Voronkevich e Alexander Averianov. [44] I resti non sono stati completamente descritti fino al 2006. Due scheletri quasi completi, articolati e una varietà di materiali disarticolati provenienti da altri individui di tutte le età sono conosciuti anche dalla Formazione Ilek, Siberia, che va dalle fasi Aptiane-Albiane del Cretaceo inferiore.[8]
La specie P. lujiatunensis , venne nominata nel 2006 dal paleontologo cinese Zhou Chang-Fu e tre colleghi cinesi, ed è una delle specie più antiche conosciute, basata su quattro teschi dagli strati inferiori della Formazione Yixian, nei pressi del villaggio di Lujiatun. [7] Sebbene questa formazione si stata datata in modo diverso da diversi autori, da 128 milioni di anni fa nella fase Barremiana, [45] a 125 milioni di anni fa nei primi anni dell'Aptiano [46] , i metodi di revisione datati hanno dimostrato che risalirebbe a circa 123 milioni di anni. [47] P. lujiatunensis fu contemporaneo di un'altra specie di psittacosauride , l' Hongshanosaurus , ritrovato nella stessa formazione. [48] È potenzialmente sinonimo di H. houi ; Sereno (2010), propose che Hongshanosaurus fosse un sinonimo di Psittacosaurus , optando per lasciare P. lujiatunensis e H. houi come due specie separate a causa delle inadeguatezze del campione di questo tipo. [31] Uno scheletro quasi completo di P. lujiatunensis dagli stessi strati della Formazione Yixian era stato precedentemente classificato nella propria specie, Psittacosaurus major , nominato per il grande cranio da Sereno, Zhao e due colleghi nel 2007. [3] You e colleghi descrissero un esemplare aggiuntivo concordando che fosse distinto da P. lujiatunensis . [30] P. major è stato originariamente caratterizzato da un cranio proporzionalmente grande, il 39% della lunghezza del corpo, rispetto al 30% di P. mongoliensis e altre caratteristiche. Tuttavia, uno studio del 2013, che utilizza l'analisi morfometrica ha mostrato che le supposte differenze tra P. lujiatunensis e P. major sono dovute a differenze di conservazione e schiacciamento. Lo studio ha concluso che entrambi rappresentano una singola specie. [47]

Una terza specie di psittacosauride dal Lujiatun, la prima ad essere stata nominata, è stata descritta come Hongshanosaurus houi , nel 2003. Il nome generico Hongshanosaurus deriva dalle parole cinese mandarino 紅 (hóng: "rosso") e 山 (shān: "collina") unito al suffisso del greco antico sauros ossia "lucertola". Questo nome si riferisce all'antica cultura Hongshan della Cina nordorientale, vissuta nella stessa area generale in cui è stato trovato il cranio fossile di Hongshanosaurus . Il nome specifico, houi , onora Hou Lianhai, professore presso l'IVPP di Pechino, che ha preparato il campione. Il genere e la specie sono stati nominati dai paleontologi cinesi Hailu, Xu Xing e Wang Xiaolin nel 2003. Sereno (2010) considerava le sue proporzioni distinte a causa della schiacciamento e della compressione dei crani di Hongshanosaurus , [31] pertanto considerò l'animale come un sinonimo junior di Psittacosaurus , e potenziale sinonimo di P. lujiatunensis . Tuttavia ciò non può essere dimostrato a causa delle cattive condizioni del cranio olotipo di H. houi . Inoltre, sono insorti dibattiti tassonomici sulla nuova combinazione del nome P. houi come nomen dubium all'interno di Psittacosaurus . [31] L'ipotesi di Sereno è stata sostenuta da uno studio morfometrico nel 2013, che ha trovato P. houi e P. lujiatunensis come sinonimi. Mentre P. houi è il nome più antico disponibile, i ricercatori hanno sostenuto che poiché l'esemplare tipo di P. lujiatunensis è meglio conservato, il nome corretto di questa specie dovrebbe essere P. lujiatunensis piuttosto che P. houi , che avrebbe normalmente priorità. [47]
La specie P. gobiensis è stata così chiamata per la regione in cui è stata ritrovata nel 2001, per poi essere descritta per la prima volta da Sereno, Zhao e Lin nel 2010. La specie è nota per un cranio e uno scheletro parziale articolato, contenente dei gastroliti . [24] Molti altri esemplari non possono essere determinati a appartenere ad alcuna specie particolare, o non sono ancora stati assegnati ad una specie in particolare. Questi esemplari sono generalmente definiti come Psittacosaurus sp. , anche se non si suppone che appartengano alla stessa specie. [9] Più di 200 esemplari di Psittacosaurus sono stati ritrovati nella Formazione Yixian, famosa per i suoi fossili di dinosauri piumati . La grande maggioranza di questi non è stata assegnata a nessuna specie conosciuta, anche se molti esemplari sono molto ben conservati e alcuni sono già stati parzialmente descritti. [49] [50] [51] Circa 100 scheletri di Psittacosaurus sono stati ritrovati in Mongolia durante le estati del 2005 e del 2006 da una squadra guidata dal paleontologo mongolo Bolortsetseg Minjin e dall'americano Jack Horner del Museo delle Montagne Rocciose del Montana . Benché solo la specie P. mongoliensis sia stata descritto dalla Mongolia finora, questi esemplari sono ancora in preparazione e non sono ancora stati assegnati a una specie in particolare. [52]
Paleobiologia
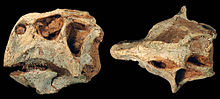
Il cervello della specie P. lujiatunensis è incredibilmente ben noto; difatti, uno studio sull'anatomia e la funzionalità del cervello dell'animale è stato condotto su tre esemplari, ed è stato pubblicato nel 2007. Fino alla pubblicazione dello studio, generalmente si pensava che il cervello dello Psittacosaurus fosse simile a quello di altri ceratopsiani , con bassi quozienti di encefalizzazione . Russell e Zhao (1996) credevano, infatti, che "le piccole dimensioni del cervello dello Psittacosaurus implicasse un repertorio comportamentale molto ristretto rispetto a quello dei mammiferi moderni di dimensioni corporee simili". Tuttavia, lo studio del 2007 ha smantellato questa ipotesi dimostrando che il cervello dell'animale era molto più avanzato di quello che si credeva. Vi è generalmente un'allometria negativa per la dimensione del cervello con lo sviluppo nei vertebrati, ma questo non è il caso di Psittacosaurus . Il punteggio EQ per P. lujiatunensis è di 0.31, significativamente superiore rispetto a generi come Triceratops . Un EQ più alto è correlato ad un comportamento più complesso, e diversi dinosauri hanno livelli elevati di EQ, simili a quelli degli uccelli, che vanno da 0,36 a 2,98. Pertanto, il comportamento dello Psittacosaurus potrebbe essere stato complesso quanto quello di un Tyrannosaurus , il cui EQ varia da 0,30 a 0,38. I comportamenti influenzati dagli alti livelli di EQ includono la costruzione di nidi, cure parentali, e un sonno simile a quello degli uccelli, comportamenti dimostrati anche da alcuni fossili di Psittacosaurus . [53]
Anche i sensi dello Psittacosaurus possono essere dedotti dall' endocasto . Il cranio dispone di grandi bulbi olfattivi , che indicano che l'animale aveva un acuto senso dell' olfatto . Le dimensioni di questi bulbi è paragonabile a quella dei grandi teropodi predatori, anche se quelli di Psittacosaurus si sono, probabilmente, evoluti per i evitare predatori anziché per cercare prede. Gli anelli sclerotici nei rettili mostrano direttamente la dimensione del bulbo oculare. Questi anelli non sono ben conservati in Psittacosaurus , con solo un individuo che li conserva probabilmente contratti, anche se si sono dimostrati essere molto simili a quelli di Protoceratops , pertanto lo Psittacosaurus doveva avere grandi occhi e una vista molto acuta. La curvatura dei canali semicircolari è legata all'agilità dei rettili, ei grandi canali curvi dello Psittacosaurus mostrano che questo animale era molto più agile rispetto ai successivi ceratopsiani. [53] Un confronto tra gli anelli sclerali di Psittacosaurus con quelli dei rettili e uccelli moderni suggerisce che l'animale era attivo durante il giorno ea brevi intervalli di notte. [54]
Ford e Martin (2010) hanno proposto che lo Psittacosaurus potesse essere un animale semi-acquatico, nuotando grazie alla propulsione della sua coda come un coccodrillo, mentre pagaiava con le zampe posteriori. Essi fondarono la loro interpretazione su alcune prove, tra cui: i depositi lacustrini (lago) dove sono stati ritrovati i fossili di molti esemplari; La posizione elevata delle narici e degli occhi; l'interpretazioni dei movimenti delle braccia e delle gambe; la coda dotata di lunghe chevron (con le setole della coda interpretate come un eventuale supporto per una pinna di pelle), fornendo una superficie propulsiva; e la presenza di gastroliti , interpretati come zavorra per le immersioni. Suggerirono inoltre che alcune specie di Psittacosaurus fossero più terrestri di altre. [55]
Dieta

Gli psittacosauri avevano denti autolivellanti utili per tagliare e sminuzzare il duro materiale vegetale. A differenza dei ceratopsiani più evoluti, gli psittacosauri non avevano i denti adatti a macinare oa masticare il cibo, per questo inerivano speciali pietre chiamati gastroliti, pietre ingerite dagli animali per sminuzzare il cibo all'interno del sistema digestivo dell'animale. All'interno di alcuni esemplari di Psittacosaurus sono state rinvenuto ben più di cinquanta di queste pietre, nelle rispettive cavità addominali di questi animali. È anche possibile che queste pietre venissero conservate nel ventriglio dell'animale, come negli uccelli moderni. [1]
A differenza di molti altri dinosauri, gli psittacosauri avevano crani akinetici: ossia la mascella e la mandibola si comportavano come una singola unità, senza giunzioni interne. L'unica giuntura era quella della mascella stessa, e gli psittacosauri potevano far scivolare la mandibola avanti e indietro sul giunto, permettendo un'azione di taglio. Diversamente dalla maggior parte dei ceratopsi, i loro becchi non formavano punte curve, ma erano invece arrotondati e appiattiti. Se le ganasce erano allineate, i becchi potevano essere utilizzati per ritagliare oggetti, ma se la mandibola inferiore fosse stata ritratta in modo che il becco inferiore fosse all'interno del becco superiore, le ganasce potrebbero aver invece avuto la funzione di "schiaccianoci". Una dieta ricca di noci o di semi sarebbe anche in accordo con la gran quantità di gastroliti rinvenuti nei vari esemplari. [24]
Funzione degli arti

Alcuni studi condotti da Phil Senter nel 2007 su P. neimongoliensis e P. mongoliensis hanno concluso che le braccia di questi taxa (e probabilmente quelli di altre specie di Psittacosaurus ) erano troppo brevi (solo il 58% degli arti posteriori) per raggiungere il terreno, e la loro gamma di moto indica che non potevano né essere pronati né generare una forza propulsiva per la locomozione, suggerendo che lo Psittacosaurus fosse un bipede obbligato. Le braccia erano troppo corte anche per essere usate per scavare o per portare il cibo alla bocca, pertanto Senter suggerì che se lo Psittacosaurus doveva scavare nel terreno, probabilmente utilizzava gli arti posteriori. Le braccia potrebbero quindi essere state usate per afferrare a due mani gli oggetti o per grattarsi il corpo corpo, ma a causa della loro limitata flessibilità e raggio di azione, potrebbero essere stati utilizzati solo per cogliere oggetti molto vicini alla pancia o ai lati dell'animale e potrebbero aver grattato solo la pancia, i fianchi e le ginocchia. Anche se le mani non riuscivano a raggiungere la bocca, lo Psittacosaurus avrebbe potuto ancora utilizzarle per trasportare materiali o cibo. [10]
Tuttavia, lo Psittacosaurus non poteva essere completamente bipede per tutta la vita. Prendendo le sezioni dalle ossa degli arti di 16 esemplari di Psittacosaurus , che vanno da un'età di meno di un anno all'età di dieci anni, Qi Zhao dell' Università di Bristol ha scoperto che lo Psittacosaurus era probabilmente un bipede facoltativo. Gli arti anteriori negli esemplari più giovani crescevano più velocemente degli arti posteriori. All'età di quattro o sei anni, la crescita delle braccia rallentava mentre la crescita delle gambe accelerava quando l'animale è diventava adulto. A questo punto, gli Psittacosaurus adulti passavano ad una posizione completamente bipede. Questi risultati rivelano inoltre che l'antenato di Psittacosaurus era probabilmente quadrupede e acquistò una posizione bipede a seguito della sua evoluzione, mentre gli esemplari più giovani erano ancora in grado di utilizzare una locomozione quadrupede. Questi risultati portano anche all'ipotesi che molte famiglie di dinosauri si siano evolute lungo questo cammino ad un certo punto della loro evoluzione. [56] [57]
Ontogenesi

Sono stati trovati diversi resti di psittacosauri giovanili. Il più piccolo è un neonato di P. mongoliensis conservato all' American Museum of Natural History (AMNH), lungo appena 11-13 centimetri, con un teschio di 2,8 centimetri. Un altro cranio presente all'AMNH è lungo solo 4,6 centimetri (1,8 pollici). Entrambi i campioni provengono dalla Mongolia. [33] I giovani scoperti nella Formazione Yixian hanno circa la stessa età del campione AMNH più grande. [49]
Un esame istologico di P. mongoliensis ha determinato il tasso di crescita di questi animali. I più piccoli esemplari dello studio sono stati stimati a tre anni d'età e meno di 1 kg (2,2 libbre), mentre i più grandi avevano nove anni e pesavano quasi 20 chilogrammi (44 libbre). Questo indica una crescita relativamente rapida rispetto alla maggior parte dei rettili e dei mammiferi marsupiali , ma più lenta degli uccelli moderni e dei mammiferi placentati . [2] Uno studio di determinazione dell'età sui residui fossili di P. mongoliensis utilizzando conteggi dell'anello di crescita suggerisce che la longevità dei ceratopsiani più primitivi era di 10-11 anni. [2]
Giovani gregari

La scoperta di un branco di sei individui di Psittacosaurus uccisi e sepolti da una colata di fango vulcanico indica la presenza di almeno due gruppi di età da due diverse frizioni riunite. Questa scoperta indicherebbe che questi animali si riunivano in gruppi di età differenti anche dopo lo svezzamento da parte dei genitori; questa rappresenta anche la prima prova di tale comportamento nei ceratopsiani. [58] Anche i denti degli esemplari più giovani erano già pronti all'uso, indicando che erano in grado di masticare il proprio cibo già in tenera età, quindi potrebbero essere stati precoci . [33] Un altro gruppo di esemplari di età diverse, dimostrerebbe che i giovani formavano frequentemente gruppi familiari e non, sia per protezione, sia per ricevere cibo, o come aiuti nelle cure parentali. [59] Tuttavia, non vi è alcuna prova di cure parentali da parte dei genitori. [60]

Nel 2004, un esemplare trovato nella Formazione Yixian ha fornito la prova di cure parentali nei dinosauri. L'esemplare DNHM D2156 è costituito da 34 scheletri articolati di giovani Psittacosaurus , strettamente associati al cranio di un adulto. I minori, approssimativamente tutti della stessa età, sono accoccolati in un gruppo sotto l'adulto, anche se tutti i 34 teschi sono posizionati sopra la massa dei corpi, come sarebbero stati in vita. Ciò suggerisce che gli animali erano vivi al momento della sepoltura, che sarebbe stata piuttosto rapida, forse dovuta al crollo della tana . [49] Tuttavia, un documento del 2013 ha rilevato che il campione adulto non appartiene al nido, e il suo cranio non aveva alcuna connessione sedimentaria alla lastra principale dove si trovavano i piccoli, che si erano fossilizzati con esso. Questa associazione artificiale ha portato alla conclusione che il cranio appartenesse ad un individuo, forse una "madre", che stava fornendo cura parentali ai 34 cuccioli, sebbene sia un'ipotesi priva di fondamenta fossili. Inoltre, un esame istologico ha dichiarato che il cranio apparteneva ad un esemplare di sei anni, mentre gli studi istologici hanno dimostrato che P. mongoliensis raggiungeva la maturità sessuale a dieci anni. È altrettanto improbabile che una sola femmina avrebbe avuto così tanti cuccioli in una sola volta. [60]
Una analisi del 2014 del medesimo esemplare sostiene l'associazione dei due fossili e ha concluso che la prossimità del campione di sei anni ai cuccioli potrebbe indicare una cooperazione post-svezzamento, rendendo il campione di sei anni un possibile custode della prole. [61] [62]
Paleopatologia
Tra le centinaia di esemplari di Psittacosaurus noti, solo uno è stato descritto in possesso di qualsiasi patologia . L'esemplare in questione, costituito da uno scheletro completo adulto e assegnato tentativamente a P. mongoliensis , è stato ritrovato negli strati inferiori della Formazione Yixian. Non ci sono segni di una frattura ossea , ma segni molto chiari di un' infezione vicino al centro del perone destro. L'osso presenta una grande fossa rotonda, prova di necrosi a causa della mancanza di approvvigionamento di sangue nella regione. La fossa è circondata da una massiccia quantità di gonfiore lungo il terzo inferiore dell'osso. Questa grande quantità di ossa depositata intorno alla lesione indica che l'animale è sopravvissuto per un bel po', nonostante la lesione e la conseguente infezione. Poiché gli psittacosauridi erano animali bipede, una lesione simile a un peso che portava l'osso nella gamba sarebbe stata probabilmente fatale. A differenza del femore e della tibia , il perone non è un osso di peso, quindi questo animale sarebbe stato ancora in grado di camminare in una certa misura. La fonte del danno non è ancora nota. [50]
Predazione

Un altro fossile proveniente dalla Formazione Yixian fornisce una prova diretta di come lo Psittacosaurus fosse una facile preda , soprattutto in tenera età. Uno scheletro di Repenomamus robustus , un grande mammifero triconodonte , è conservato con i resti di un giovane Psittacosaurus nella sua cavità addominale. Alcune delle ossa del cucciolo sono ancora articolate, indicando che il mammifero predatore ha inghiottito la sua preda a grossi pezzi. Questo esemplare è notevole in quanto è il primo esempio noto di un mammifero mesozoico in grado di predare dinosauri vivi. [63] La pesante predazione sui giovani Psittacosaurus da parte di Repenomamus e, probabilmente, da altri predatori della zona, può aver causato la Selezione R , ossia la produzione di prole più numerose per contrastare le perdite dovute ai predatori.[64]
Paleoecologia

Lo Psittacosaurus è conosciuto da centinaia di singoli esemplari, di cui oltre 75 sono stati assegnati alla specie tipo , P. mongoliensis . [9] [50] [52] Tutti i fossili di Psittacosaurus scoperti fino ad ora sono stati ritrovati nei sedimenti del Cretaceo inferiore in Asia , dalla Siberia meridionale fino alla Cina settentrionale e forse fino al sud della Thailandia . L'età più comune delle formazioni geologiche che portano i fossili di Psittacosaurus è quella del Barremiano superiore all'Albiano del Cretaceo inferiore, circa 126-101 milioni di anni fa. [46] Molte formazioni sedimentarie terrestri di questa età in Mongolia e nella Cina settentrionale hanno prodotto fossili di Psittacosaurus , portando alla definizione di questo periodo di tempo nella regione come lo " Psittacosaurus biochron". [65] [66]
La specie più antica conosciuta è P. lujiatunensis , ritrovata negli strati più bassi della Formazione Yixian. [7] Oltre 200 esemplari attribuiti a questo genere sono stati recuperati da questi e altri letti dello Yixian, la cui età esatta è oggetto di dibattito. [50] Anche se molti studi iniziali usavano la datazione radiometrica hanno messo Yixian nel Giurassico , ciò non coincide con la gamma temporale prevista per Psittacosaurus , la cui specie più antica risale al Cretaceo inferiore. Usando una radiazione argon-argon, un team di scienziati cinesi ha datato i letti più bassi della formazione a circa 128 milioni di anni fa, mentre i alti a circa 122 milioni di anni fa. [45] Uno studio cinese più recente, utilizzando l'apporto di uranio, suggerisce che gli strati inferiori siano più giovani, circa 123.2 milioni di anni fa, mentre acconsentono ad un'età di 122 milioni di anni fa per gli strati superiori. [46]
Note
- ^ a b PC Sereno, Psittacosauridae , in Philip J. Currie e Kevin P. Padian (a cura di), The Encyclopedia of Dinosaurs , Academic Press, 1997, pp. 611–613.
- ^ a b c GM† Erickson e TA Tumanova,Growth curve of Psittacosaurus mongoliensis Osborn (Ceratopsia: Psittacosauridae) inferred from long bone histology , in Zoological Journal of the Linnean Society , vol. 130, n. 4, 2000, pp. 551–566, DOI : 10.1111/j.1096-3642.2000.tb02201.x .
- ^ a b c PC Sereno, X. Zhao, L. Brown e L. Tan, New psittacosaurid highlights skull enlargement in horned dinosaurs ( PDF ), in Acta Palaeontologica Polonica , vol. 52, n. 2, 2007, pp. 275–284.
- ^ a b c d e f g h i DA Russell e X. Zhao, New psittacosaur occurrences in Inner Mongolia , in Canadian Journal of Earth Sciences , vol. 33, n. 4, 1996, pp. 637–648, DOI : 10.1139/e96-047 .
- ^ a b c d e DB Brinkman, DA Eberth, MJ Ryan e P. Chen, The occurrence of Psittacosaurus xinjiangensis Sereno and Chow, 1988 in the Urho area, Junggar basin, Xinjiang , in Canadian Journal of Earth Sciences , vol. 38, n. 12, 2001, pp. 1781–1786, DOI : 10.1139/e01-049 .
- ^ a b c Paul C. Sereno, Zhao Xijin, Chang Zhengwu e Rao Chenggang, Psittacosaurus meileyingensis (Ornithischia: Ceratopsia), a new psittacosaur from the Lower Cretaceous of northeastern China , in Journal of Vertebrate Paleontology , vol. 8, 1988, pp. 366–377, DOI : 10.1080/02724634.1988.10011725 .
- ^ a b c d e f g h Zhou Changfu, Gao Keqin, Richard C. Fox e Chen Shuihua, A new species of Psittacosaurus (Dinosauria: Ceratopsia) from the Early Cretaceous Yixian Formation, Liaoning, China , in Palaeoworld , vol. 15, 2006, pp. 100–114.
- ^ a b c d e f g h Alexander O. Averianov, Alexei V. Voronkevich, Sergei V. Leshchinskiy e Alexei V. Fayngertz, A ceratopsian dinosaur Psittacosaurus sibiricus from the Early Cretaceous of West Siberia, Russia and its phylogenetic relationships , in Journal of Systematic Paleontology , vol. 4, n. 4, 2006, pp. 359–395, DOI : 10.1017/s1477201906001933 .
- ^ a b c d e f g h i j k l m n o You Hailu & Dodson, Peter. (2004). Basal Ceratopsia. In: Weishampel, David B., Dodson, Peter, & Osmolska, Halszka (Eds.). The Dinosauria (2nd Edition). Berkeley: University of California Press. Pp. 478–493.
- ^ a b P Senter, Analysis of forelimb function in basal ceratopsians , in Journal of Zoology , vol. 273, 2007, pp. 305–314, DOI : 10.1111/j.1469-7998.2007.00329.x .
- ^ a b G. Mayr, SD Peters, G. Plodowski e O. Vogel, Bristle-like integumentary structures at the tail of the horned dinosaur Psittacosaurus ( PDF ), in Naturwissenschaften , vol. 89, 2002, pp. 361–365, DOI : 10.1007/s00114-002-0339-6 .
- ^ Jakob Vinther, Robert Nicholls, Stephan Lautenschlager, Michael Pittman, Thomas G. Kaye, Emily Rayfield, Gerald Mayr, Innes C. Cuthill. 3D Camouflage in an Ornithischian Dinosaur . Current Biology , 2016; DOI: 10.1016/j.cub.2016.06.065
- ^ T. Lingham-Soliar, A unique cross section through the skin of the dinosaur Psittacosaurus from China showing a complex fibre architecture , in Proceedings of the Royal Society B: Biological Sciences , vol. 275, n. 1636, 2008, pp. 775–780, DOI : 10.1098/rspb.2007.1342 .
- ^ Jakob Vinther, Robert Nicholls, Stephen Lautenschlager, Michael Pittman, Thomas G. Kaye, Emily Rayfield, Gerard Mayr e Innes C. Cuthill, 3D Camouflage in an Ornithischian Dinosaur ( PDF ) [ collegamento interrotto ] , in Current Biology , vol. 26, 2016, pp. 1-7, DOI : 10.1016/j.cub.2016.06.065 .
- ^ Scientists reveal most accurate depiction of a dinosaur ever created , su theguardian.com , 14 settembre 2016. URL consultato il 15 settembre 2016 .
- ^ This Dinosaur Wore Camouflage , su news.nationalgeographic.com , 14 settembre 2016. URL consultato il 15 settembre 2016 .
- ^ http://www.cell.com/cms/attachment/2063917292/2065805129/mmc1.pdf
- ^ Gerald Mayr, Michael Pittman, Evan Saitta, Thomas G. Kaye, Jakob Vinther e Roger Benson, Structure and homology of Psittacosaurus tail bristles , in Palaeontology , 2016, DOI : 10.1111/pala.12257 .
- ^ a b Henry F. Osborn, Psittacosaurus and Protiguanodon : Two Lower Cretaceous iguanodonts from Mongolia , in American Museum Novitates , vol. 127, 1924, pp. 1–16.
- ^ Gregory M. Erickson e Tatyana A. Tumanova,Growth curve of Psittacosaurus mongoliensis Osborn (Ceratopsia: Psittacosauridae) inferred from long bone histology , in Zoological Journal of the Linnean Society , vol. 130, n. 4, 2000, pp. 551–566, DOI : 10.1111/j.1096-3642.2000.tb02201.x .
- ^ a b c d e f g Xing Xu e Xijin Zhao, Psittacosaur fossils and their stratigraphical implications , in Wang, Y. e Deng, T. (a cura di), Proceedings of the Seventh Annual Meeting of the Chinese Society of Vertebrate Paleontology , Beijing, China Ocean Press, 1999, pp. 75–80.
- ^ a b c d e f Paul C. Sereno, New data on parrot-beaked dinosaurs ( Psittacosaurus ) , in Ken Carpenter e Philip J. Currie (a cura di), Dinosaur Systematics: Perspectives and Approaches , Cambridge, Cambridge University Press, 1990, pp. 203–210.
- ^ a b Eric Buffetaut e Varavudh Suteethorn, A new species of the ornithischian dinosaur Psittacosaurus from the Early Cretaceous of Thailand , in Palaeontology , vol. 35, 1992, pp. 801–812.
- ^ a b c Paul C. Sereno, Zhao Xijin e Tan Lin, A new psittacosaur from Inner Mongolia and the parrot-like structure and function of the psittacosaur skull ( PDF ), in Proceedings of the Royal Society B , vol. 277, n. 1679, 2010, pp. 199–209, DOI : 10.1098/rspb.2009.0691 , PMC 2842669 , PMID 19535376 .
- ^ a b c d Henry F. Osborn, Two Lower Cretaceous dinosaurs of Mongolia , in American Museum Novitates , vol. 95, n. 13, 1923, pp. 1–10.
- ^ Hailu You, Xing Xu e Xiaolin Wang,A new genus of Psittacosauridae (Dinosauria: Ornithopoda) and the origin and early evolution of marginocephalian dinosaurs , in Acta Geologica Sinica , vol. 77, n. 1, English, 2003, pp. 15–20, DOI : 10.1111/j.1755-6724.2003.tb00105.x .
- ^ Xu Xing, Catherine A. Forster, James M. Clark e Jinyou Mo,A basal ceratopsian with transitional features from the Late Jurassic of northwestern China , in Proceedings of the Royal Society B: Biological Sciences , vol. 273, n. 1598, 2006, pp. 2135–2140, DOI : 10.1098/rspb.2006.3566 , PMC 1635516 , PMID 16901832 .
- ^ AA Farke, WD Maxwell, RL Cifelli e MJ Wedel,A Ceratopsian Dinosaur from the Lower Cretaceous of Western North America, and the Biogeography of Neoceratopsia , in PLoS ONE , vol. 9, n. 12, 2014, pp. e112055, Bibcode : 2014PLoSO...9k2055F , DOI : 10.1371/journal.pone.0112055 , PMC 4262212 , PMID 25494182 .
- ^ a b Xing Xu, A new psittacosaur ( Psittacosaurus mazongshanensis sp. nov.) from Mazongshan area, Gansu Province, China , in Z. Dong (a cura di), Sino-Japanese Silk Road Dinosaur Expedition , Beijing, China Ocean Press, 1997, pp. 48–67.
- ^ a b Hai-Lu You, Kyo Tanoue e Peter Dodson, New data on cranial anatomy of the ceratopsian dinosaur Psittacosaurus major ( PDF ), in Acta Palaeontologica Polonica , vol. 53, n. 2, 2008, pp. 183–196, DOI : 10.4202/app.2008.0202 .
- ^ a b c d e f g h i j k l m n o p Paul C. Sereno, Taxonomy, cranial morphology, and relationships of parrot-beaked dinosaurs (Ceratopsia: Psittacosaurus ) , in Michael J. Ryan, Brenda J. Chinnery-Allgeier e David A. Eberth (a cura di), New Perspectives on Horned Dinosaurs: The Royal Tyrrell Museum Ceratopsian Symposium , Bloomington and Indianapolis, Indiana University Press, 2010, pp. 21–58.
- ^ Henry F Osborn, Three new Theropoda, Protoceratops zone, Central Mongolia , in American Museum Novitates , vol. 144, 1924, pp. 1–12.
- ^ a b c Walter P Coombs, Juvenile specimens of the ornithischian dinosaur Psittacosaurus , in Palaeontology , vol. 25, 1982, pp. 89–107.
- ^ a b CC Young, The dinosaur remains of Laiyang, Shantung , in Palaeontologia Sinica Series C , vol. 16, 1958, pp. 53–159.
- ^ a b CC Young, On some new dinosaurs from western Suiyuan, Inner Mongolia , in Bulletin of the Geological Society of China , vol. 11, n. 3, 1931, pp. 259–266, DOI : 10.1111/j.1755-6724.1932.mp11003005.x .
- ^ ( ZH ) Zhengwu Cheng, [Reptilia] , in [The Mesozoic Stratigraphy and Paleontology of the Guyang Coal-bearing Basin, Nei Monggol Autonomous Region, China] , Beijing, Geology Press, 1983, pp. 123–136.
- ^ a b c Paul C. Sereno, The fossil record, systematics and evolution of pachycephalosaurs and ceratopsians from Asia , in Michael J. Benton, Mikhail A. Shishkin, David M. Unwin e Evgeny N. Kurochkin (a cura di), The Age of Dinosaurs in Russia and Mongolia , Cambridge, Cambridge University Press, 2000, pp. 480–516.
- ^ J. David. Archibald, Species , in Philip J. Currie e Kevin Padian (a cura di), The Encyclopedia of Dinosaurs , San Diego, Academic Press, 1997, pp. 695–699.
- ^ ( ZH ) Xijin Zhao, [Concerning a new species of Psittacosaurus from Laiyang, Shantung] , in Vertebrata PalAsiatica , vol. 6, 1962, pp. 349–360.
- ^ Paul C. Sereno e Zhao Xijin, Psittacosaurus xinjiangensis (Ornithischia: Ceratopsia), a new psittacosaur from the Lower Cretaceous of northwestern China , in Journal of Vertebrate Paleontology , vol. 8, n. 4, 1988, pp. 353–365, DOI : 10.1080/02724634.1988.10011724 .
- ^ David A. Eberth, Dale A. Russell, DR Braman e AL Deino, The age of the dinosaur-bearing sediments at Tebch, Inner Mongolia, People's Republic of China , in Canadian Journal of Earth Sciences , vol. 30, n. 10, 1993, pp. 2101–2106, Bibcode : 1993CaJES..30.2101E , DOI : 10.1139/e93-182 .
- ^ E. Buffetaut e V. Suteethorn, Remarks on P. sattayaraki Buffetaut & Suteethorn, 1992, a ceratopsian dinosaur from the Lower Cretaceous of Thailand , in Oryctos , vol. 4, 2002, pp. 71–73.
- ^ F. Tang, Z. Luo, Z. Zhou, H. You, JA Georgi, Z. Tang e X. Wang, Biostratigraphy and palaeoenvironment of the dinosaur-bearing sediments in Lower Cretaceous of Mazongshan area, Gansu Province, China , in Cretaceous Research , vol. 22, 2001, pp. 115–129, DOI : 10.1006/cres.2000.0242 .
- ^ ( RU ) Sergei V. Leshchinskiy, Alexei V. Fayngertz, Alexei V. Voronkevich, EN Maschenko e Alexander O. Averianov, Preliminary results of the investigation of the Shestakovo localities of Early Cretaceous vertebrates , in AV Komarov (a cura di), Materials of the Regional Conference of the Geologists of Siberia, Far East and North East of Russia , Tomsk, GalaPress, 2000, pp. 363–366.
- ^ a b S Wang, H Hu, P Li e Y Wang, Further discussion on the geologic age of Sihetun vertebrate assemblage in western Liaoning, China: evidence from Ar-Ar dating , in Acta Petrologica Sinica , vol. 17, 2001, pp. 663–668.
- ^ a b c W. Yang, S. Li e B. Jiang, New evidence for Cretaceous age of the feathered dinosaurs of Liaoning: Zircon U-Pb SHRIMP dating of the Yixian Formation in Sihetun, northeast China , in Cretaceous Research , vol. 28, n. 2, 2007, pp. 177–182, DOI : 10.1016/j.cretres.2006.05.011 .
- ^ a b c BP Hedrick e P. Dodson,Lujiatun Psittacosaurids: Understanding Individual and Taphonomic Variation Using 3D Geometric Morphometrics , in Alistair Robert Evans (a cura di), PLoS ONE , vol. 8, n. 8, 2013, pp. e69265, DOI : 10.1371/journal.pone.0069265 , PMC 3739782 , PMID 23950887 .
- ^ Hailu You e Xing Xu,An adult specimen of Hongshanosaurus houi (Dinosauria: Psittacosauridae) from the Lower Cretaceous of Western Liaoning Province, China , in Acta Geologica Sinica , vol. 79, n. 2, English, 2005, pp. 198–173, DOI : 10.1111/j.1755-6724.2005.tb00879.x .
- ^ a b c Meng Qingjin, Liu Jinyuan, David J. Varrichio, Timothy Huang e Gao Chunling, Parental care in an ornithischian dinosaur , in Nature , vol. 431, n. 7005, 2004, pp. 145–146, Bibcode : 2004Natur.431..145M , DOI : 10.1038/431145a , PMID 15356619 .
- ^ a b c d Junchang Lü, Yoshitsugu Kobayashi, Yuong-Nam Lee e Qiang Ji, A new Psittacosaurus (Dinosauria: Ceratopsia) specimen from the Yixian Formation of western Liaoning, China: the first pathological psittacosaurid , in Cretaceous Research , vol. 28, n. 2, 2007, pp. 272–276, DOI : 10.1016/j.cretres.2006.08.005 .
- ^ Gerald Mayr, Plodowski Peters, D. Stephan, Gerhard e Olaf Vogel, Bristle-like integumentary structures at the tail of the horned dinosaur Psittacosaurus , in Naturwissenschaften , vol. 89, n. 8, 2002, pp. 361–365, Bibcode : 2002NW.....89..361M , DOI : 10.1007/s00114-002-0339-6 , PMID 12435037 .
- ^ a b Evelyn Boswell, MSU, Mongolian paleontologists find 67 dinosaurs in one week , su montana.edu , Montana State University News Service, 13 settembre 2006. URL consultato il 5 maggio 2007 .
- ^ a b CF Zhou, KQ Gao, RC Fox e XK Du, Endocranial morphology of psittacosaurs (Dinosauria: Ceratopsia) based on CT scans of new fossils from the Lower Cretaceous, China , in Palaeoworld , vol. 16, n. 4, 2007, pp. 285–293, DOI : 10.1016/j.palwor.2007.07.002 .
- ^ Schmitz, L. e Motani, R., Nocturnality in Dinosaurs Inferred from Scleral Ring and Orbit Morphology , in Science , vol. 332, n. 6030, 2011, pp. 705–8, Bibcode : 2011Sci...332..705S , DOI : 10.1126/science.1200043 , PMID 21493820 .
- ^ Tracy L. Ford e Martin, Larry D., A semi-aquatic life habit for Psittacosaurus , in Ryan, Michael J.; Chinnery-Allgeier, Brenda J.; and Eberth, David A. (editors.) (a cura di), New Perspectives on Horned Dinosaurs: The Royal Tyrrell Museum Ceratopsian Symposium , Bloomington and Indianapolis, Indiana University Press, 2010, pp. 328–339, ISBN 978-0-253-35358-0 .
- ^ University of Bristol, How 'parrot dinosaur' switched from four feet to two as it grew , su sciencedaily.com , ScienceDaily, 28 giugno 2013.
- ^ Qi Zhao, Michael J. Benton, Corwin Sullivan, P. Martin Sander e Xing Xu, Histology and postural change during the growth of the ceratopsian dinosaur Psittacosaurus lujiatunensis , in Nature Communications , vol. 4, 2013, p. 2079, Bibcode : 2013NatCo...4E2079Z , DOI : 10.1038/ncomms3079 , PMID 23811819 .
- ^ Q. Zhao, PM Barrett e DA Eberth, Social behaviour and mass mortality in the basal ceratopsian dinosaur Psittacosaurus (Early Cretaceous, People's Republic of China)". , in Palaeontology , vol. 50, n. 5, 2007, pp. 1023–1029, DOI : 10.1111/j.1475-4983.2007.00709.x .
- ^ RD Alexander, The evolution of social behaviour , in Annual Review of Ecology and Systematics , vol. 5, 1974, pp. 325–383, DOI : 10.1146/annurev.es.05.110174.001545 .
- ^ a b Q. Zhao, Juvenile-only clusters and behaviour of the Early Cretaceous dinosaur Psittacosaurus , in Acta Palaeontologica Polonica , 2013, DOI : 10.4202/app.2012.0128 .
- ^ Paleontologists describe a possible dinosaur nest and young 'babysitter' , su phys.org . URL consultato il 27 agosto 2014 .
- ^ BP Hedrick, Gao Chunling, GI Omar, Zhang Fengjiao, Shen Caizhi e P. Dodson, The osteology and taphonomy of a Psittacosaurus bonebed assemblage of the Yixian Formation (Lower Cretaceous), Liaoning, China , in Cretaceous Research , vol. 51, 2014, pp. 321–340, DOI : 10.1016/j.cretres.2014.06.015 .
- ^ Hu Yaoming, Meng Jin, Wang Yuanqing e Li Chuankui, Large Mesozoic mammals fed on dinosaurs , in Nature , vol. 433, n. 7022, 2005, pp. 149–152, Bibcode : 2005Natur.433..149H , DOI : 10.1038/nature03102 , PMID 15650737 .
- ^ Q Zhao, Benton, MJ, Xu, X. e Sander, MJ, Juvenile-only clusters and the behaviour of the Early Cretaceous dinosaur Psittacosaurus ( PDF ), in Acta Palaeontologica Polonica , vol. 59, n. 4, 2014, pp. 827–833.
- ^ Spencer G. Lucas, The Psittacosaurus biochron, Early Cretaceous of Asia , in Cretaceous Research , vol. 27, n. 2, 2006, pp. 189–198, DOI : 10.1016/j.cretres.2005.11.011 .
- ^ Spencer G. Lucas, Chinese Fossil Vertebrates , 2001, pp. 168 –170.
Altri progetti
-
 Wikimedia Commons contiene immagini o altri file su Psittacosaurus
Wikimedia Commons contiene immagini o altri file su Psittacosaurus -
 Wikispecies contiene informazioni su Psittacosaurus
Wikispecies contiene informazioni su Psittacosaurus
Collegamenti esterni
- ( EN ) Psittacosaurus , su Enciclopedia Britannica , Encyclopædia Britannica, Inc.
- ( EN ) Psittacosaurus , su Fossilworks.org .