Metabolismul fierului
Metabolismul fierului uman este ansamblul reacțiilor chimice care mențin homeostazia fierului uman la nivel sistemic și celular. Fierul este atât necesar pentru corp, cât și potențial toxic, iar controlul nivelului de fier din organism este o parte importantă a multor aspecte ale sănătății și bolilor umane. Hematologii s-au interesat în mod special de metabolismul sistemic al fierului, deoarece fierul este esențial pentru celulele roșii din sânge, unde este conținută cea mai mare parte a fierului din corpul uman. Înțelegerea metabolismului fierului este, de asemenea, importantă pentru înțelegerea bolilor de supraîncărcare a fierului, cum ar fi hemocromatoza ereditară și carența de fier, cum ar fi anemia cu deficit de fier .
Funcțiile fierului în corpul uman

Fierul este un bioelement esențial pentru majoritatea formelor de viață, de la bacterii la mamifere. Importanța sa constă în capacitatea sa de a intermedia transferul de electroni. În starea feroasă, fierul acționează ca un donator de electroni, în timp ce în starea ferică acționează ca un acceptor. Prin urmare, fierul joacă un rol fundamental în cataliza reacțiilor enzimatice care implică transferul de electroni (reducere și oxidare, redox). Proteinele pot conține fier ca parte a mai multor cofactori, cum ar fi grupurile fier-sulf (Fe-S) și grupările hem, care sunt ambele asamblate în mitocondrii.
Respirație celulară
Celulele umane necesită fier pentru a obține energie ca ATP dintr-un proces în mai multe etape cunoscut sub numele de respirație celulară, mai precis din fosforilarea oxidativă a crestelor mitocondriale. Fierul este prezent în grupele fier-sulf și hem de proteine din lanțul de transport al electronilor care generează un gradient de protoni care permite ATP sintaza să sintetizeze ATP (chemosmoza).
Grupurile hemo fac parte din hemoglobină, o proteină care se găsește în celulele roșii din sânge și care servește la transportul oxigenului din plămâni către țesuturi. Grupurile hemo sunt, de asemenea, prezente în mioglobină pentru a stoca și difuza oxigenul în celulele musculare.
Transportul oxigenului
Corpul uman are nevoie de fier pentru a transporta oxigenul. Oxigenul (O 2 ) este necesar pentru funcționarea și supraviețuirea a aproape toate tipurile de celule. Oxigenul este transportat de la plămâni la restul corpului legat de grupul hem al hemoglobinei din eritrocite. În celulele musculare, fierul leagă mioglobina, care reglează eliberarea acestuia.
Toxicitate
Fierul este, de asemenea, potențial toxic. Capacitatea sa de a dona și accepta electroni înseamnă că poate cataliza conversia peroxidului de hidrogen în radicali liberi. Radicalii liberi pot provoca daune unei mari varietăți de structuri celulare și, în cele din urmă, pot ucide celula. [1]
Fierul legat de proteine sau cofactori precum hemul este sigur. Mai mult, nu există practic ioni de fier cu adevărat liberi în celulă, deoarece formează cu ușurință complexe cu molecule organice. Cu toate acestea, o parte din fierul intracelular este legat de complexele cu afinitate scăzută și este denumit fier labil sau „liber”. Fierul din astfel de complexe poate provoca daune așa cum este descris mai sus. [2]
Pentru a preveni acest tip de daune, toate formele de viață care utilizează fier leagă atomii de fier de proteine. Această legare permite celulelor să beneficieze de fier și, în același timp, să-și limiteze capacitatea de a face rău. [3] Concentrațiile tipice de fier labil intracelular în bacterii sunt de 10-20 μM, [4] deși pot fi de 10 ori mai mari în mediul anaerob, [5] unde radicalii liberi și speciile de oxigen reactiv sunt mai rare. În celulele de mamifere, concentrațiile de fier labil intracelular sunt de obicei mai mici de 1 μM, mai puțin de 5% din fierul celular total.
Protecție împotriva bacteriilor

Ca răspuns la o infecție bacteriană sistemică, sistemul imunitar inițiază un proces cunoscut sub numele de retenție de fier. Dacă bacteriile supraviețuiesc, trebuie să obțină fier din mediul lor. Bacteriile cauzatoare de boli fac acest lucru în mai multe moduri, inclusiv prin eliberarea de molecule care leagă fierul numite siderofori și apoi le reabsorbe pentru a recupera fierul sau prin eliminarea fierului din hemoglobină și transferină. Cu cât este mai greu să lucrezi pentru fier, cu atât este mai mare prețul metabolic pe care trebuie să îl plătească. Aceasta înseamnă că bacteriile fără fier se reproduc mai lent. Așadar, controlul nivelului de fier pare a fi o apărare importantă împotriva majorității infecțiilor bacteriene; Cu toate acestea, există câteva excepții. TBC-ul care cauzează bacteria poate locui în macrofage care sunt un mediu bogat în fier, iar Borrelia burgdorferi folosește mangan în loc de fier. Persoanele cu cantități mai mari de fier, cum ar fi persoanele cu hemocromatoză, sunt mai susceptibile la unele infecții bacteriene. [6]
În timp ce acest mecanism este un răspuns elegant la infecțiile bacteriene pe termen scurt, poate provoca probleme atunci când inflamația durează mai mult. Deoarece ficatul produce hepcidină ca răspuns la citokinele inflamatorii, nivelurile de hepcidină pot crește ca urmare a surselor neinflamatorii de inflamație, cum ar fi infecții virale, cancer, boli autoimune sau alte boli cronice. Când se întâmplă acest lucru, sechestrarea cu fier pare a fi principala cauză a sindromului de anemie a bolilor cronice, în care nu este disponibil suficient fier pentru a produce suficient globule roșii care conțin hemoglobină.
Zăcăminte de fier
Cei mai bine hrăniți oameni din țările industrializate au între 4 și 5 grame de fier în corp (~ 38 mg de fier / kg de greutate corporală pentru femei și ~ 50 mg de fier / kg de corp pentru bărbați) [7] . Dintre acestea, aproximativ 2,5 sunt conținute în hemoglobina necesară pentru a transporta oxigenul prin sânge, iar cea mai mare parte a restului (aproximativ 2 grame la bărbații adulți și puțin mai puțin la femeile aflate la vârsta fertilă) este conținută în complexele de feritină care sunt prezente în toate celulele, dar cele mai frecvente în măduva osoasă, ficat și splină. Depozitele de feritină hepatică sunt principala sursă fiziologică de fier de rezervă din organism. Rezervele de fier din țările industrializate tind să fie mai mici la copii și femei în vârstă fertilă decât la bărbați și vârstnici. Femeile care trebuie să își folosească rezervele pentru a compensa fierul pierdut în timpul menstruației, sarcinii sau alăptării au depozite corporale mai mici de non-hemoglobină, care pot fi de 500 miligrame sau mai puțin.
Din conținutul total de fier al corpului, aproximativ 400 mg este destinat proteinelor celulare care utilizează fierul pentru procesele celulare importante, cum ar fi stocarea oxigenului ( mioglobina ) sau efectuarea de reacții redox producătoare de energie ( citocromi ). O cantitate relativ mică (3-4 mg) circulă prin plasmă, legată de transferină. [8] Datorită toxicității sale, fierul solubil liber este menținut la o concentrație scăzută în organism.
Deficitul de fier afectează mai întâi fierul de stocare în organism, iar epuizarea acestor depozite este considerată relativ nesimptomatică, deși au fost asociate unele simptome vagi și nespecifice. Deoarece fierul este necesar în principal pentru hemoglobină, anemia cu deficit de fier este principala manifestare clinică a deficitului de fier. Persoanele cu deficit de fier vor suferi sau vor muri din cauza deteriorării organelor înainte ca celulele să rămână fără fier, necesare proceselor intracelulare, cum ar fi transportul electronilor.
Macrofagele sistemului reticuloendotelial stochează fierul ca parte a procesului de descompunere și procesare a hemoglobinei din celulele roșii din sânge. Fierul este, de asemenea, stocat ca un pigment numit hemosiderin care este un depozit slab definit de proteine și fier, creat de macrofage în care fierul în exces este prezent, local sau sistemic, de exemplu în rândul persoanelor cu supraîncărcare de fier din cauza distrugerii frecvente și a transfuziei celulelor sanguine. Dacă supraîncărcarea sistemică a fierului este corectată, hemosiderina este reabsorbită încet de macrofage în timp.
Mecanisme de reglare a fierului
Homeostazia fierului uman este reglementată pe două niveluri diferite. Nivelurile sistemice de fier sunt echilibrate prin absorbția controlată a fierului din dietă de către enterocite, celule care acoperă interiorul intestinului și pierderea necontrolată de fier din epitaxial, sudoare, răni și pierderea de sânge. Mai mult, fierul sistemic este reciclat continuu. Nivelurile de fier celulare sunt controlate diferit de diferite tipuri de celule datorită expresiei anumitor proteine reglatoare și de transport ale fierului.
Reglarea sistemică a fierului

Absorbția fierului din dietă
Absorbția fierului din dietă este un proces variabil și dinamic. Cantitatea de fier absorbită în raport cu cantitatea ingerată este în general redusă, dar poate varia de la 5% până la 35% în funcție de circumstanțe și de tipul de fier. Eficiența cu care fierul este absorbit variază în funcție de sursă. În general, cele mai bune forme de fier absorbite provin din produse de origine animală. Absorbția fierului din dietă sub formă de săruri de fier (ca în majoritatea suplimentelor) variază ușor în funcție de necesitatea organismului de fier și este de obicei între 10% și 20% din aportul de fier. Absorbția fierului din produsele de origine animală și unele produse vegetale se prezintă sub formă de fier hemic și este mai eficientă, permițând absorbția a 15% până la 35% din aport. Fierul hem la animale provine din sânge și proteine care conțin hem în carne și mitocondrii, în timp ce la plante, fierul hem este prezent în mitocondrii în toate celulele care utilizează oxigen pentru respirație.
La fel ca majoritatea nutrienților minerali, cea mai mare parte a fierului absorbit din alimentele sau suplimentele digerate este absorbit în duoden de către enterocitele căptușelii duodenale. Aceste celule au molecule speciale care le permit să deplaseze fierul în jurul corpului. Fierul din dietă poate fi absorbit fie ca parte a unei proteine, cum ar fi proteina hem, fie în forma sa feroasă Fe 2+ . Citocromul duodenal B (Dcytb), o enzimă ferică reductază situată în marginea periei suprafeței luminale a enterocitelor, reduce Fe 3+ (fier feric) la Fe 2+ (fier feros) [9] . O proteină numită transportor de metal divalent 1 ( DMT1 ), care poate transporta mai multe metale divalente peste membrana plasmatică, transportă apoi fierul prin membrana celulei enterocite în celulă.
Aceste celule de căptușeală intestinală pot stoca fierul ca feritină, care se obține din Fe 3+ prin legarea de apoferritină (caz în care fierul va părăsi corpul atunci când celula moare și se excretă în scaun), sau celula o poate elibera prin scaunul.corp prin singurul exportator de fier cunoscut la mamifere, feroportina . Hefestina , o feroxidază care poate oxida Fe 2+ la Fe 3+ și se găsește în principal în intestinul subțire, ajută feroportina să transfere fierul prin capătul basolateral al celulelor intestinale. În schimb, feroportina este reprimată după traducere de hepcidin, un hormon peptidic de 25 aminoacizi. Corpul reglează nivelurile de fier reglând fiecare dintre acești pași. De exemplu, enterocitele sintetizează mai multe Dcytb, DMT1 și feroportină ca răspuns la anemia cu deficit de fier. [10] Absorbția fierului din dietă este crescută în prezența vitaminei C și scăzută de excesul de calciu, zinc sau mangan. [11]
Rata de absorbție a fierului din corpul uman pare să răspundă la o varietate de factori interdependenți, inclusiv depozitele totale de fier, măsura în care măduva osoasă produce noi globule roșii, concentrația de hemoglobină din sânge și conținutul de oxigen din sânge. sânge. De asemenea, corpul absoarbe mai puțin fier în perioadele de inflamație pentru a priva bacteriile de fier. Descoperirile recente arată că reglarea feroportinei hepcidinei este responsabilă de sindromul anemiei bolii cronice.
Reciclarea și pierderea fierului
Majoritatea fierului din corp este stocat și reciclat de sistemul reticuloendotelial, care descompune celulele roșii din sânge. Spre deosebire de absorbția și reciclarea fierului, nu există un mecanism fiziologic de reglare pentru excreția acestuia. Oamenii pierd o cantitate mică, dar constantă de fier din cauza pierderii de sânge gastro-intestinal, transpirație și răsturnarea celulelor pielii și a mucoasei tractului gastro-intestinal. Cantitatea totală de pierderi pentru persoanele sănătoase din lumea dezvoltată se ridică la o medie estimată de 1 mg pe zi pentru bărbați și 1,5-2 mg pe zi pentru femeile cu perioade menstruale regulate. Persoanele cu infecții parazitare gastrointestinale, mai frecvent întâlnite în țările în curs de dezvoltare, pierd adesea mai mult. [1] Cei care nu reușesc să regleze absorbția suficient de bine suferă de tulburări de supraîncărcare a fierului. În aceste boli, toxicitatea fierului începe să copleșească capacitatea organismului de a-l lega și de a-l păstra. [12]
Reglarea celulară a fierului
Aport de fier
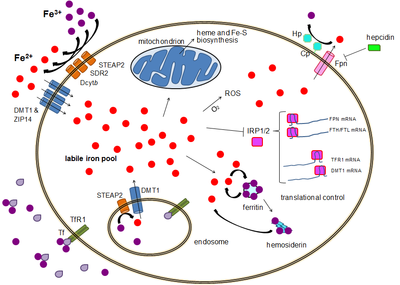
Majoritatea tipurilor de celule absorb fierul în principal prin endocitoza mediată de receptor prin receptorul transferinei 1 (TFR1), receptorul transferinei 2 (TFR2) și GAPDH. TFR1 are o afinitate pentru fier legat de transferină de 30 de ori mai mare decât TFR2 și, prin urmare, este principalul jucător în acest proces. [13] [14] Enzima glicolerală multifuncțională gliceraldehidă-3-fosfat dehidrogenază (GAPDH) de grad superior funcționează și ca receptor al transferinei. [15] [16] Fe 3+ legat de transferină este recunoscut de acești receptori de transferină, declanșând o schimbare conformațională care provoacă endocitoză. Fierul intră apoi în citoplasmă de la endosom prin intermediul importatorului DMT1 după ce a fost redus la starea feroasă de o familie de reductaze STEAP. [17]
Alternativ, fierul poate intra direct în celulă prin intermediul importatorilor de cationi de membrană plasmatică bivalentă, cum ar fi DMT1 și ZIP14 (proteina de tip Zrt-Irt 14). [18] Din nou, fierul intră în citoplasmă în stare feroasă după ce a fost redus în spațiul extracelular de o reductază precum STEAP2, STEAP3 (în eritrocite), Dcytb (în enterocite) și SDR2. [17]
Rezerva de fier Labile
În citoplasmă, fierul feros se află într-o stare solubilă și chelatabilă, care constituie rezerva de fier labil (~ 0,001 mM). [19] [20] Se crede că fierul din aceste rezerve este legat de compuși cu masă redusă, cum ar fi peptide, carboxilați și fosfați, deși unii pot fi sub formă liberă hidratată (ioni de apă). Alternativ, ionii de fier ar putea fi legați de proteine specializate cunoscute sub numele de metalochaperone. [21] În mod specific, proteinele de legare poli-r (C) par să treacă de la transferul liber de fier la feritină (pentru depozitare) și enzime non-hem (pentru utilizare în cataliză). [18] [22] Bazinul de fier labil este potențial toxic datorită capacității fierului de a genera specii reactive de oxigen. Fierul din acest bazin poate fi absorbit de mitocondrii prin mitoferrină pentru a sintetiza grupurile Fe-S și grupurile hem. [17]
Depozitare fier
Fierul poate fi depozitat în feritină ca fier feric datorită activității feritoxinei cu lanț greu de feritină. [23] Feritina disfuncțională se poate acumula sub formă de hemosiderină, ceea ce poate fi problematic în caz de supraîncărcare cu fier. [24] Rezerva de fier de stocare a feritinei este mult mai mare decât rezerva de fier labilă, cu o concentrație între 0,7 mM și 3,6 mM. [19]
Expulzarea fierului
Expulzarea fierului are loc într-o varietate de tipuri de celule, inclusiv neuroni, eritrocite, macrofage și enterocite. Ultimele două sunt deosebit de importante deoarece nivelurile sistemice de fier depind de ele. Există un singur exportator de fier cunoscut, feroportinul. [25] Transportul fierului feros din celulă, în general ajutat de ceruloplasmină și / sau efestin (în principal în enterocite), care oxidează fierul la starea sa ferică, astfel încât să se poată lega la transferină în mediul extracelular. [17] Hepcidina determină internalizarea feroportinei, scăzând exportul de fier. Mai mult, hepcidina pare să regleze atât TFR1 cât și DMT1 printr-un mecanism necunoscut. [26] Un alt jucător care ajută feroportina în efectuarea exportului de fier celular este GAPDH. [27] O izoformă GAPDH post-translațională modificată este recrutată pe suprafața celulelor încărcate cu fier unde recrutează apo-transferină în imediata apropiere a feroportinei pentru a chela rapid fierul extrudat. [28]
Expresia hepcidinei, care apare doar în anumite tipuri de celule, cum ar fi hepatocitele, este strict controlată la nivel transcripțional și reprezintă legătura dintre homeostazia celulară și sistemică a fierului datorită rolului hepcidinei ca „gatekeeper” eliberarea fierului din enterocite în restul corpului. Eritroblastele produc eritroferon, un hormon care inhibă hepcidina și astfel crește disponibilitatea fierului necesar pentru sinteza hemoglobinei. [29]
Controlul transportului celular al fierului
Deși există mai mult de un control la nivel transcripțional, reglarea nivelurilor de fier celular la nivel de translație este controlată fundamental de proteinele care leagă elementul reactiv de fier IRP1 și în special IRP2. [30] Atunci când nivelurile de fier sunt scăzute, aceste proteine sunt capabile să se lege de elementele sensibile la fier (IRE). IRE sunt prezente în regiunile netraduse (UTR) ale ARNm. [17]
Atât feritina, cât și feroportinul conțin un IRE în UTR-ul lor de 5 ', astfel încât, prin deficit de fier, translația lor este reprimată de IRP2, prevenind sinteza inutilă a proteinelor de stocare și exportul dăunător de fier. În schimb, TFR1 și unele variante DMT1 conțin 3 URE IRE, care leagă IRP2 sub deficit de fier, stabilizând ARNm, care asigură sinteza importatorilor de fier. [17]
Patologia legată de fier
Deficiență de fier

Deficiența de fier funcțională sau efectivă poate fi rezultatul diferiților factori care pot fi grupați după cum urmează:
- Cerere crescută de fier, pe care dieta nu o poate adapta.
- Pierderi crescute de fier (de obicei datorate pierderii de sânge).
- Deficiență nutrițională. Poate rezulta din lipsa fierului alimentar sau din consumul de alimente care inhibă absorbția fierului. Acestea includ calciu conținut în suplimente sau produse lactate, fitați de tărâțe [31] și taninuri conținute în ceai [32] .
- Administrarea de medicamente antiacide : Aceste medicamente, utilizate în general pentru tratarea gastritei , a bolii de reflux gastroesofagian și a ulcerelor, reduc absorbția fierului din dietă. In plus , inhibitori ai pompei de protoni , H 2 antihistaminice și antiacidele reduc metabolismul fierului [33] .
- Deteriorarea mucoasei intestinale cauzată de exemplu de o intervenție chirurgicală care implică duodenul sau boli precum boala Crohn sau boala celiacă care reduc drastic suprafața disponibilă pentru absorbție.
- Inflamație care determină restricționarea indusă de hepcidină a eliberării de fier din enterocite (vezi mai sus).
Suprasolicitare de fier
Corpul este capabil să reducă substanțial cantitatea de fier pe care o absoarbe prin membrana mucoasă. Nu pare capabil să întrerupă complet procesul de transport al fierului. De asemenea, în situațiile în care excesul de fier dăunează peretelui intestinal în sine (de exemplu, atunci când copiii mănâncă cantități mari de tablete de fier făcute pentru consumul adulților), chiar mai mult fier poate pătrunde în sânge și poate provoca un sindrom care pune viața în pericol. Cantități mari de fier liber în circulație vor provoca leziuni ale celulelor critice din ficat, inimă și alte organe metabolice active.
Toxicitatea fierului apare atunci când cantitatea de fier care circulă depășește cantitatea de transferină disponibilă pentru a-l lega, dar corpul este capabil să-și regleze energic absorbția fierului. Prin urmare, toxicitatea fierului ingerat se datorează, de obicei, unor circumstanțe extraordinare, cum ar fi consumul excesiv de suplimente de fier [34] , mai degrabă decât modificărilor alimentare. Tipul de toxicitate acută prin ingestia de fier provoacă diverse probleme, inclusiv afectarea severă a membranei mucoase a tractului gastro-intestinal.
Excesul de fier a fost legat de unele tipuri de cancer. Un studiu recent a arătat că pacienții cu cancer mamar cu niveluri scăzute de feroportină (care determină concentrații mai mari de fier intracelular) au în medie o speranță de viață mai mică. Pe de altă parte, nivelurile ridicate de feroportină duc în 90% din cazuri la o supraviețuire de 10 ani. [35]
Toxicitatea cronică a fierului este de obicei rezultatul mai multor sindroame cronice de supraîncărcare a fierului asociate cu boli genetice, transfuzii repetate sau alte cauze. În astfel de cazuri, depozitele de fier ale unui adult pot ajunge la 50 de grame (de 10 ori mai mult decât fierul normal al corpului) sau mai mult. Exemplele clasice de supraîncărcare genetică cu fier includ hemocromatoza ereditară și hemocromatoza juvenilă. Aceasta din urmă, mai severă decât prima, este cauzată de mutații fie în gena RGMc, aparținând uneia dintre familiile de molecule de ghidare repulsive , [36] cunoscută și sub numele de hemojuvelină, HJV sau HFE2, fie din gena HAMP pe care o codifică (care este o peptidă de reglare a fierului). Majoritatea tulburărilor genetice de supraîncărcare a fierului constau în diferite forme de hemocromatoză la adulți, dar mecanismele exacte nu sunt încă cunoscute. Prin urmare, cercetătorii au reușit să identifice mutațiile genetice care cauzează diferite variante de hemocromatoză la adulți și acum trebuie să înțeleagă funcția normală a acestor gene mutante.
Notă
- ^ a b Conrad ME, Umbreit JN, Tulburări ale metabolismului fierului , în The New England Journal of Medicine , vol. 342, n. 17, aprilie 2000, pp. 1293-4, DOI : 10.1056 / NEJM200004273421716 , PMID 10787338 .
- ^ Kakhlon O, Cabantchik ZI, The labile iron pool: caracterizare, măsurare și participare la procesele celulare , în Free Radical Biology and Medicine , vol. 33, nr. 8, 2002, pp. 1037-1046, DOI : 10.1016 / s0891-5849 (02) 01006-7 .
- ^ Andrews NC, Tulburări ale metabolismului fierului , în The New England Journal of Medicine , vol. 341, nr. 26, decembrie 1999, pp. 1986-95, DOI : 10.1056 / NEJM199912233412607 , PMID 10607817 .
- ^ Yan Y, Waite-Cusic JG, Kuppusamy P, Yousef AE, fier liber intracelular și rolul său potențial în inactivarea Escherichia coli indusă de presiune ultra-înaltă , în Microbiologie aplicată și de mediu , vol. 79, nr. 2, ianuarie 2013, pp. 722-724, DOI : 10.1128 / aem.02202-12 , PMID 23124235 .
- ^ Yamamoto Y, Fukui K, Koujin N, Ohya H, Kimura K, Kamio Y, Regularea bazinului de fier liber intracelular de către Dpr oferă toleranță la oxigen la Streptococcus mutans , în Journal of Bacteriology , vol. 186, nr. 18, 2004, pp. 5997-6002, DOI : 10.1128 / jb.186.18.5997-6002.2004 , PMID 15342568 .
- ^ Ganz T, Hepcidin, un regulator cheie al metabolismului fierului și mediator al anemiei inflamației , în Sânge , vol. 102, nr. 3, august 2003, pp. 783-8, DOI : 10.1182 / blood-2003-03-0672 , PMID 12663437 .
- ^ Sareen S. Gropper și Jack L. Smith, Advanced Nutrition and Human Metabolism , 6th, Belmont, CA, Wadsworth, 2013, p. 481, ISBN 978-1133104056 .
- ^ Camaschella C, Schrier SL, Regulamentul balanței de fier , uptodate.com , 7 noiembrie 2011. Accesat la 3 noiembrie 2011 .
- ^ AT McKie, o fericreductază feric-reglementată asociată cu absorbția fierului dietetic , în Știință , vol. 291, nr. 5509, 2 martie 2001, pp. 1755-1759, DOI : 10.1126 / science.1057206 . Adus la 16 martie 2019 .
- ^ (EN) Robert E. Fleming și Bruce R. Bacon, Orchestration of Iron Homeostasis în New England Journal of Medicine, vol. 352, nr. 17, 28 aprilie 2005, pp. 1741-1744, DOI : 10.1056 / NEJMp048363 . Adus la 16 martie 2019 .
- ^ Copie arhivată , la ohioline.osu.edu . Adus pe 9 octombrie 2018 (arhivat din original la 16 iunie 2012) .
- ^ uptodate.com , http://www.uptodate.com/contents/iron-overload-syndromes-other-than-hereditary-hemochromatosis .
- ^ (EN) Hiroshi Kawabata, Rasha S. Germain și Peter T. Vuong,Transferrin Receptor 2-α Sprijină creșterea celulară atât în celulele cultivate cu fier în chelat, cât și în Vivo , în Journal of Biological Chemistry, vol. 275, nr. 22, 2 iunie 2000, pp. 16618-16625, DOI : 10.1074 / jbc.M908846199 . Adus la 16 martie 2019 .
- ^ (EN) Anthony P. West, Melanie J. Bennett și Vera M. Sellers,Comparația interacțiunilor dintre receptorul transferinei și receptorul transferinei 2 hemocromatoza ereditară cu transferrina și proteina HFE , în Journal of Biological Chemistry, vol. 275, nr. 49, 8 decembrie 2000, pp. 38135-38138, DOI : 10.1074 / jbc.C000664200 . Adus la 16 martie 2019 .
- ^ (EN) Santosh Kumar, Navdeep Sheokand și Mayur Anant Mhadeshwar, Caracterizarea gliceraldehidei-3-fosfat dehidrogenazei ca un receptor nou al transferinei , în The International Journal of Biochemistry & Cell Biology, vol. 44, nr. 1, 2012-1, pp. 189-199, DOI : 10.1016 / j.biocel.2011.10.016 . Adus la 16 martie 2019 .
- ^ (EN) Navdeep Sheokand, Santosh Kumar și Himanshu Malhotra, Gliceraldehye secretat-3-fosfat dehidrogenază este un receptor de transferină celulară autocrină multifuncțional pentru achiziționarea fierului , în Biochimica și Biophysica Acta (BBA) - Subiecte generale, vol. 1830, nr. 6, 2013-6, pp. 3816-3827, DOI : 10.1016 / j.bbagen.2013.03.019 . Adus la 16 martie 2019 .
- ^ a b c d e f ( EN ) Matthias W. Hentze, Martina U. Muckenthaler și Bruno Galy, Two to Tango: Regulation of Mammalian Iron Metabolism , în Cell , vol. 142, nr. 1, 2010-7, pp. 24-38, DOI : 10.1016 / j.cell.2010.06.028 . Adus la 16 martie 2019 .
- ^ a b ( EN ) DJR Lane, AM Merlot și ML-H. Huang, absorbția, traficul și metabolismul fierului celular: molecule și mecanisme cheie și rolurile lor în boli , în Biochimica și Biophysica Acta (BBA) - Molecular Cell Research , vol. 1853, nr. 5, 2015-5, pp. 1130-1144, DOI : 10.1016 / j.bbamcr.2015.01.021 . Adus la 16 martie 2019 .
- ^ a b ( EN ) Deficitul și supraîncărcarea de fier: de la biologia de bază la medicina clinică , în nutriție și sănătate , Humana Press, 2010, ISBN 9781934115220 . Adus la 16 martie 2019 .
- ^ Kakhlon O, Cabantchik ZI. și Kakhlon O, Cabantchik ZI., Rezerva de fier labil: caracterizare, măsurare și participare la procesele celulare , în Free Radic Biol Med. 2002 15 octombrie; 33 (8): 1037-46. .
- ^ LA Finney, Transition Metal Speciation in the Cell: Insights from the Chemistry of Metal Ion Receptors , în Știință , vol. 300, n. 5621, 9 mai 2003, pp. 931-936, DOI : 10.1126 / science.1085049 . Adus la 16 martie 2019 .
- ^ Caroline C. Philpott e Moon-Suhn Ryu, Special delivery: distributing iron in the cytosol of mammalian cells , in Frontiers in Pharmacology , vol. 5, 22 luglio 2014, DOI : 10.3389/fphar.2014.00173 . URL consultato il 16 marzo 2019 .
- ^ ( EN ) Paolo Arosio e Sonia Levi, Ferritin, iron homeostasis, and oxidative damage1,2 1Guest Editor: Mario Comporti 2This article is part of a series of reviews on “Iron and Cellular Redox Status.” The full list of papers may be found on the homepage of the journal. , in Free Radical Biology and Medicine , vol. 33, n. 4, 2002-8, pp. 457-463, DOI : 10.1016/S0891-5849(02)00842-0 . URL consultato il 16 marzo 2019 .
- ^ ( EN ) Tracey A. Rouault e Sharon Cooperman, Brain Iron Metabolism , in Seminars in Pediatric Neurology , vol. 13, n. 3, 2006-9, pp. 142-148, DOI : 10.1016/j.spen.2006.08.002 . URL consultato il 16 marzo 2019 .
- ^ ( EN ) Tomas Ganz, Cellular iron: Ferroportin is the only way out , in Cell Metabolism , vol. 1, n. 3, 2005-3, pp. 155-157, DOI : 10.1016/j.cmet.2005.02.005 . URL consultato il 16 marzo 2019 .
- ^ ( EN ) Fang Du, Christopher Qian e Zhong Ming Qian, Hepcidin directly inhibits transferrin receptor 1 expression in astrocytes via a cyclic AMP-protein kinase a pathway , in Glia , vol. 59, n. 6, 2011-6, pp. 936-945, DOI : 10.1002/glia.21166 . URL consultato il 16 marzo 2019 .
- ^ ( EN ) Vishant Mahendra Boradia, Manoj Raje e Chaaya Iyengar Raje, Protein moonlighting in iron metabolism: glyceraldehyde-3-phosphate dehydrogenase (GAPDH) , in Biochemical Society Transactions , vol. 42, n. 6, 1º dicembre 2014, pp. 1796-1801, DOI : 10.1042/BST20140220 . URL consultato il 16 marzo 2019 .
- ^ ( EN ) N. Sheokand, H. Malhotra e S. Kumar,Moonlighting cell-surface GAPDH recruits apotransferrin to effect iron egress from mammalian cells , in Journal of Cell Science , vol. 127, n. 19, 1º ottobre 2014, pp. 4279-4291, DOI : 10.1242/jcs.154005 . URL consultato il 16 marzo 2019 .
- ^ ( EN ) Léon Kautz, Grace Jung e Erika V Valore, Identification of erythroferrone as an erythroid regulator of iron metabolism , in Nature Genetics , vol. 46, n. 7, 2014-7, pp. 678-684, DOI : 10.1038/ng.2996 . URL consultato il 16 marzo 2019 .
- ^ ( EN ) Martina U. Muckenthaler, Bruno Galy e Matthias W. Hentze, Systemic Iron Homeostasis and the Iron-Responsive Element/Iron-Regulatory Protein (IRE/IRP) Regulatory Network , in Annual Review of Nutrition , vol. 28, n. 1, 2008-8, pp. 197-213, DOI : 10.1146/annurev.nutr.28.061807.155521 . URL consultato il 16 marzo 2019 .
- ^ ( EN ) Leif Hallberg, Wheat fiber, phytates and iron absorption , in Scandinavian Journal of Gastroenterology , vol. 22, sup129, 1987-1, pp. 73-79, DOI : 10.3109/00365528709095855 . URL consultato il 16 marzo 2019 .
- ^ ( EN ) PB Disler, SR Lynch e RW Charlton, The effect of tea on iron absorption. , in Gut , vol. 16, n. 3, 1º marzo 1975, pp. 193-200, DOI : 10.1136/gut.16.3.193 . URL consultato il 16 marzo 2019 .
- ^ UpToDate , su www.uptodate.com . URL consultato il 16 marzo 2019 .
- ^ Rudolph, Colin David, 1953-, Rudolph's pediatrics , 21st ed, McGraw-Hill, Medical Pub. Division, 2003, ISBN 0838582850 , OCLC 45575698 . URL consultato il 16 marzo 2019 .
- ^ ( EN ) ZK Pinnix, LD Miller e W. Wang, Ferroportin and Iron Regulation in Breast Cancer Progression and Prognosis , in Science Translational Medicine , vol. 2, n. 43, 4 agosto 2010, pp. 43ra56–43ra56, DOI : 10.1126/scitranslmed.3001127 . URL consultato il 16 marzo 2019 .
- ^ ( EN ) Christopher J. Severyn, Ujwal Shinde e Peter Rotwein, Molecular biology, genetics and biochemistry of the repulsive guidance molecule family , in Biochemical Journal , vol. 422, n. 3, 15 settembre 2009, pp. 393-403, DOI : 10.1042/BJ20090978 . URL consultato il 16 marzo 2019 .
Voci correlate
Collegamenti esterni
- ( EN ) A comprehensive NIH factsheet on iron and nutrition , su ods.od.nih.gov .
- ( EN ) Iron Disorders Institute: A nonprofit group concerned with iron disorders; site has helpful links and information on iron-related medical disorders. , su irondisorders.org .
- ( EN ) An interactive medical learning portal on iron metabolism , su ironatlas.com .
- ( EN ) Information about iron outside the body , su webelements.com .
| Controllo di autorità | LCCN ( EN ) sh85068202 |
|---|