Dilophosaurus wetherilli
| Dilophosaurus | |
|---|---|
 | |
| Starea de conservare | |
Fosil | |
| Clasificare științifică | |
| Domeniu | Eukaryota |
| Regatul | Animalia |
| Phylum | Chordata |
| Clasă | Sauropsida |
| Superordine | Dinozauria |
| Ordin | Saurischia |
| Subordine | Theropoda |
| Familie | † Dilophosauridae |
| Tip | † Dilophosaurus Welles , 1970 |
| Nomenclatura binominala | |
| † Dilophosaurus wetherilli Welles, 1970 | |
| Sinonime | |
| |
Dilophosaurus (al cărui înseamnă „șopârlă două crested“ nume), numite în mod obișnuit Dilophosaurus în limba italiană, este un gen pe cale de disparitie de dilophosaurid theropod dinozaur care a trăit în Jurasic de Jos , în urmă cu aproximativ 193 milioane ani ( Sinemurian ), în ceea ce este acum Formațiunea Kayenta , Arizona , America de Nord . Dilophosaurus a fost unul dintre cei mai mari dinozauri prădători ai timpului său, cu o lungime totală de 7 metri și își datorează numele unei perechi de creste rotunjite paralele pe craniu.
Descriere
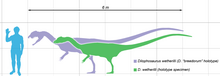
Dilophosaurus a fost unul dintre cei mai mari prădători ai timpului său. Un exemplar adult ar putea ajunge până la 7 metri (23 de picioare) în lungime, cântărind aproximativ 400 de kilograme (880 de lire sterline). [1]
Dinții lui Dilophosaurus erau foarte lungi, dar relativ subțiri și nu foarte largi la bază. [2] Dentiția animalului avea 12 dinți maxilari și 18 dinți mandibulari ; dinții de pe vârful maxilarului superior erau mai mici. Al doilea și al treilea dinți anteriori aveau indentări, absente în al patrulea. [3] O altă caracteristică neobișnuită a craniului a fost o depresiune profundă între maxilar și premaxilă , făcând craniul Dilophosaurus foarte similar cu cel al spinosauridelor . Această „depresie” există în virtutea unei legături slabe între oasele premaxilare și maxilare. [4] Craniul Dilophosaurus este bine cunoscut de paleontologi și este semnificativ, deoarece are o caracteristică a peretelui lateral superior care este absent în ceratozaurie . [3] Comparativ cu ceratozaurii, expansiunea scapulară distală în D. wetherilli este unic dreptunghiulară. [3] Osul coapsei ( femurul ) a fost mai lung decât tibia . [3]
Un studiu realizat de Robert J. Gay ( 2005 ), comparând diferite specimene, nu a găsit urme de dimorfism sexual între cele două sexe ale Dilophosaurus , întrucât atât bărbații, cât și femelele împărtășeau aceeași anatomie a scheletului. [5]
Conform lui Rauhut ( 2000 ), Dilophosaurus poate fi recunoscut pe baza următoarelor caracteristici: osul lacrimal are o îngroșare în partea dorsal-posterioară ; vertebrele cervicale au spini neuronali lungi cu o „capotă” ascuțită în procesele centrale și anterioare și posterioare; lama scapulară are expansiune distală; prezența crestelor nazolacrimale subțiri împerecheate vertical spre acoperișul craniului, fiecare cu o proiecție posterioară mai subțire (Carrano, 2012 ). [3] [6] [7]
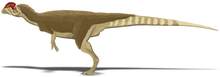
Creste craniene

Cea mai distinctivă trăsătură a Dilophosaurus este evident perechea de creste rotunjite paralele de pe craniu, constând dintr-o extensie a oaselor nazale și lacrimale . Aceste structuri par a fi mult prea delicate pentru orice scop, cu excepția vizualizării intraspecifice. [3] [8] Dodson (1997) a observat că Dilophosaurus a fost primul dinozaur care a posedat creste craniene, care ar fi mai târziu prezent în majoritatea teropodelor timpurii. [9]
Funcția crestelor craniului Dilophosaurus a fost un subiect de speculații în rândul oamenilor de știință de când au fost descoperite. În mod tradițional, aceste bizare structuri craniene (și nenumăratele structuri osoase din numeroase alte genuri) au fost întotdeauna interpretate ca instrumente pentru a atrage colegi și / sau a intimida rivalii și potențialii prădători ai altor specii. Padian, Horner și Dhaliwal (2004) susțin, pe baza analizei filogenetice histologice și a dovezilor funcționale, că aceste structuri bizare au fost probabil folosite pentru recunoașterea unui singur exemplar în cadrul speciei. [10] [11]
Clasificare
Clasificarea Dilophosaurus a fost examinată de mai multe ori de-a lungul anilor și genul a fost atribuit nu mai puțin de nouă grupuri diferite de teropode . Welles (1954) și majoritatea analizelor filogenetice ulterioare între 1980 și 1990 au clasificat acest gen ca un celofisoid mare în cadrul taxonilor Coelophysoidea . [8] [13] Cu toate acestea, majoritatea studiilor din secolul XXI până în prezent au arătat că Dilophosaurus și diferite alte dilofosauride sunt mai strâns legate de tetanidele teropode decât de adevăratele celofisoide. [14] [15]
Următoarea cladogramă arată analiza filogenetică a teropodelor și a Dilophosaurus , conform studiilor lui Hendrickx și colab. (2015): [16]
| Neotheropoda |
| |||||||||||||||||||||||||||||||||||||||||||||||||||
Istoria descoperirii

Primele exemplare de Dilophosaurus au fost descoperite de Sam Welles în vara anului 1942, în Formația Kayenta, Arizona . [17] Situl fusese găsit de un navajo , Jesse Williams, în 1940. Cei trei indivizi erau prezenți, dar unul dintre ei a fost expus unei săpături utile. Două probe au fost urmărite la Berkeley pentru curățarea și montarea oaselor, de la echipa Wann Langston, formând „ holotype UCMP 37302 (proba pe care se bazează numele), și un al doilea pe schelet metalic, paratype UCMP 37303. L“ animalul numit Megalosaurus wetherilli de Welles, în 1954. [18] Numele specific, wetherilli îi aduce un omagiu lui John Wetherill, care a explorat zona descoperirii. [19]
Revenind la aceeași formațiune, în 1964 , pentru a determina perioadei la care aparțineau oasele, Welles a găsit un nou specimen, UCMP 77270, nu departe de locul descoperirii anterioare. Pe baza crestelor duble craniene conservate în acest nou specimen pentru prima dată în noul schelet, Welles a realizat că specia era semnificativ diferită de Megalosaurus . În 1970, el a reclasificat animalul într - un nou gen, pe care el a numit Dilophosaurus, [2] [18] din greacă cuvintele δι care înseamnă „doi“, λόφος / lophos care înseamnă „creasta“ și σαυρος / sauros care înseamnă „șopârlă "; astfel numele complet înseamnă „șopârlă cu două creste”. [19] Dovezile disponibile sugerează cu tărie că noul exemplar trebuie să fi fost holotipul lui D. wetherilli, iar al doilea schelet să fie exemplare mai tinere. [3]
În 2001 , Robert Gay a raportat descoperirea a două sau, eventual, a trei noi exemplare de Dilophosaurus wetherilli în colecțiile Muzeului din Arizona de Nord. Exemplarele au fost găsite în 1978 în formațiunea Kayenta. Gay a comparat proba cu holotipul (UCMP 37302) dovedind că aparțin aceluiași gen. Gay a remarcat, de asemenea, că acest material este semnificativ, deoarece include părți ale bazinului care nu sunt conservate în holotipul sau specimenul „ D. breedorum ” UCMP 77270. [2]
Gay a mai raportat că unele articole din colecția Museum of Northern Arizona erau reprezentative pentru un specimen nou-născut (MNA P1.3181). Specimenul în cauză include un humerus parțial, o fibulă parțială și un fragment de dinte . Specimenul nou-născut este semnificativ din mai multe motive: în primul rând, arată ratele de creștere ale dinozaurilor timpurii și, în al doilea rând, oferă cercetătorilor posibilitatea de a compara rata de creștere a Dilophosaurus cu cele ale unei specii înrudite, cum ar fi Coelophysis. Care ar putea oferi informații în ratele de creștere ale teropodelor timpurii. Acesta este, de asemenea, primul exemplar tânăr din gen. [2]
Hu Shaojin (1993) a atribuit exemplarul KMV 8701 unei a doua specii de Dilophosaurus , pe care au numit-o Dilophosaurus sinensis . [20] Această specie a fost găsită în provincia Yunnan , China , în 1987, cu prosauropodul Yunnanosaurus huangi . [21] Comparativ cu D. wetherilli , această specie era mai mare și mai robustă. Lamanna și colab. (1998b) au examinat materialul atribuit lui D. sinensis dovedind că nu este altceva decât un sinonim pentru Sinosaurus triassicus . [22] Această constatare a fost confirmată în 2013. [23]
În 1999, a fost inventată o a treia specie presupusă, D. breedorum , nume inventat de Samuel Welles pentru proba UCMP 77270, colectată în 1964. Tykoski (2005) a efectuat o analiză ontogenetică în teza sa și a concluzionat că D. breedorum este doar un Dilophosaurus wetherilli adult. [24] Acesta a fost primul astfel de exemplar care a păstrat o creastă craniană aproape completă. [25] Welles a concluzionat că diferențele dintre exemplarele originale și UCMP 77270 justificau denumirea în specii separate. [3] [26] Această specie nu a fost considerată valabilă în alte recenzii ale genului, care o văd ca un nomen nudum și / sau ca un sinonim al lui D. wetherilli . [2] [27]
Paleobiologie


Dilophosaurus este considerat un biped obligatoriu datorită prezenței membrelor posterioare lungi orientate vertical sub pelvis, în timp ce membrele anterioare erau scurte și incapabile să susțină locomoția cvadrupedală. Osul coapsei (femurul) era mai lung decât cel al piciorului (tibia), sugerând că animalul era un alergător agil și rapid, așa cum s-ar fi așteptat de la un dinozaur prădător. [3]
Welles (1984) a propus că Dilophosaurus era un animal gregar , trăind în grupuri mici, pe baza faptului că majoritatea scheletelor au fost găsite în agregare. [3] Cu toate acestea, Gay (2001) a declarat că nu există dovezi ale unui astfel de comportament și a remarcat că inundațiile erau destul de frecvente în acel moment și că ar fi putut colecta material împrăștiat și izolat de la diferiți indivizi și depozitat împreună în aceeași zonă. [2] Cu toate acestea, dacă funcția crestelor ar fi fost doar pentru vizualizare, ar avea sens ca aceste animale să trăiască în grupuri sociale mici. [9]
Dietă
Prezența și distribuția suturilor ne-interdigitate în craniile unor grupuri de reptile, inclusiv Dilophosaurus , a fost interpretată ca o indicație a prezenței unui sistem de pârghii, ghidat de mușchii maxilarului , ca un ajutor pentru prădare. [28] Welles (1984) a respins această ipoteză și a interpretat mobilitatea potențială a craniului Dilophosaurus ca un semn de slăbiciune și a susținut că legătura liberă cu premaxilla nu ar fi permis capturarea și subjugarea unei pradă. [3] Acest lucru a condus la ipoteza inițială conform căreia Dilophosaurus era un scuturator , hrănindu-se cu cari , deoarece dinții lui erau prea slabi și subțiri pentru a doborî prada mare și crestele sale erau prea fragile pentru a fi implicate într-o singură. [4] Recent, unii cercetători au susținut că Dilophosaurus ar fi putut fi un piscivor. Dovada acestui comportament ar fi descoperirea amprentelor lui Eubrontes în timpul înotului, precum și asemănările anatomice ale craniului cu cele ale spinosauridelor . [29]
Creştere
Tkach (1996) a efectuat un studiu al structurii osoase a specimenului de tip și a concluzionat că Dilophosaurus wetherilli ar fi avut rate de creștere de aproape 35 de kilograme (77 lbs) în primii ani de viață, reflectând o rată de creștere foarte rapidă. [30]
Paleopatologie

Unul dintre exemplarele Dilophosaurus prezintă numeroase leziuni osoase cauzate de rănirea sau zdrobirea unei vertebre și un potențial abces pe un humerus. Un alt exemplar are un humerus stâng neobișnuit de mic decât cel drept, un posibil exemplu de „asimetrie plutitoare”. Asimetria fluctuantă apare din tulburările de dezvoltare și este mai frecventă la populațiile stresate și, prin urmare, poate oferi informații despre calitatea condițiilor în care a trăit dinozaurul.[31]
Un alt exemplu de Dilophosaurus , de la Muzeul de Paleontologie al Universității din California, etichetat ca UCMP 37302, prezintă semne ale a opt patologii diferite ale brâului umărului și ale membrelor anterioare, inclusiv trei tumori osoase, unele oase rupte și altele vindecate pe ambele. și incapacitatea de a utiliza unul dintre membre anterioare. Deși nu este sigur, se crede că majoritatea sau toate rănile acestui specimen au fost cauzate într-o coliziune cu un alt animal (posibil un alt Dilophosaurus ) sau într-un accident. Cu toate acestea, teropodul pare să fi supraviețuit suficient de mult pentru ca unele dintre rănile sale să se vindece. Patologiile acestui exemplar incredibil au fost documentate în jurnalul PLoS One . [32]
Într-un studiu din 2001 realizat de Bruce Rothschild și alți paleontologi, au fost examinate aproximativ 60 de oase de picioare, unele dintre ele din Dilophosaurus pentru semne de fracturi de stres, dar nu s-a găsit nicio urmă în oasele analizate. [33]
Paleoecologie

Rămășițele specimenului tip Dilophosaurus wetherilli (UCMP 37302) și scheletul parțial UCMP 37303, au fost recuperate în Silty Facies, membru al formației Kayenta, în nord-estul Arizona . Rămășițele au fost descoperite în 1942 și 1964 de la șisturi albastru și maro depuse în timpul Sinemurian - Pliensbachian faze ale Jurasic , în urmă cu aproximativ 196-183 milioane de ani. Alte două exemplare atribuite acestui gen au fost descoperite în aceeași formațiune; unul în 1982 (UCMP 130053) într-un canal de gresie și celălalt într-un bloc de gresie. [34]
Formația Kayenta face parte din grupul Glen Canyon, care include formațiuni nu numai din nordul Arizona, ci și părți din sud-estul Utah , vestul Colorado și nord-vestul New Mexico . Se compune în principal din două facies , unul dominat de depuneri de nămol și celălalt dominat de gresie . Faciesul cu lut se găsește în mare parte în Arizona, în timp ce faciesul de gresie se găsește în zone din nordul Arizona, sudul Utah, vestul Colorado și nord-vestul New Mexico. Prima formațiune care s-a așezat a fost faciesul noroios ca parte a unui sistem fluvial mai lent. Formația pare să se fi format atunci când dunele, care ar deveni gresie Navajo, au invadat zona. [35] O datare radiometrică definitivă a acestei formațiuni nu a fost încă realizată, iar corelația stratigrafică disponibilă s-a bazat pe o combinație de date radiometrice a fosilelor vertebrate, magnetostratigrafie și dovezi ale polenului. [34] Acesta a fost emis ipoteza că formarea Kayenta a fost depus între Sinemurian și Pliensbachian , din perioada Jurasicului, în urmă cu aproximativ 196-183 milioane de ani. [36]

Formația Kayenta a păstrat o colecție mică, dar în creștere, de organisme. Majoritatea fosilelor provin din faciesul limoasei. [37] Cele mai multe organisme cunoscute până acum sunt vertebrate Organismele non-vertebrate includ calcarele microbiene sau „alge”, [38] lemnul pietrificat, [39] amprentele plantelor, [40] bivalvi și melci de apă dulce, [35] ostracodele [41] și urmă de fosile de nevertebrate. [38]
Vertebratele sunt cunoscute atât din fosilele osoase, cât și din amprentele fosile. Vertebratele cunoscute din fosile osoase ale corpului includ: [37] rechini hybodonti , pești osoși nedeterminați, dipnoi [39] , salamandre, [42] broasca Prosalirus , Caecilia Eocaecilia , broasca țestoasă Kayentachelys , o reptilă sfenodonte , diverse șopârle [ 43] , mai mulți crocodylomorphi primitivi, inclusiv Calsoyasuchus , Eopneumatosuchus , Kayentasuchus și Protosuchus , pterosaurul Rhamphinion și unele mamifere, inclusiv Dinnetherium și un mamifer haramyid. Numeroși dinozauri au fost de asemenea prezenți în formație: au existat mai mulți teropode, inclusiv Dilophosaurus , Kayentavenator [44] , Coelophysis kayentakatae și teropodul "shake N Bake"; ierbivore a inclus primitiv sauropodomorpho Sarahsaurus [45] , un heterodontosaurid și dinozauri blindate scelidosaurus și scutellosaurus . Formarea de asemenea , a găzduit tritylodontid synapsids Dinnebiton , Kayentatherium și Oligokyphus . [43] Majoritatea acestor artefacte provin din vecinătatea Gold Spring, Arizona . [37] Urmele fosilizate ale vertebratelor includ, de asemenea, coprolite [38] și urme fosilizate ale teraselor . [46]
Explorările din anii 1970 și 1980 ale lui James M. Clark, Farish Jenkins și David E. Fastovsky, colecția și analiza lui William R. Downs au dovedit existența mai multor exemplare de vertebrate. Formația Kayenta a produs mai multe locuri de înmormântare în masă, inclusiv unul care conține rămășițele a trei taxoni celofisoizi de dimensiuni corporale diferite, reprezentând cea mai cunoscută colecție faunistică de ceratozauri . [47]
Alte rămășițe atribuite Dilophosaurus au fost recuperate în straturile superioare ale formațiunii Dharmaram, din Andhra Pradesh , India . Rămășițele au fost descoperite în noroiul întărit care a fost depus în timpul Sinemurianului jurasic , cu aproximativ 196-189 milioane de ani în urmă. Aceleași sedimente conțineau și fosile unui crocodil , un sauropodomorf și sauropodul Lamplughsaura . [48]
Icnologie

În formațiunea Kayenta din Arizona, aceeași formațiune care a dat probele originale ale acestui gen, s-au găsit numeroase urme de fosile clasificate ca Kayentapus hopii și Dilophosauripus williamsi , dar astăzi atribuite în mod normal Dilophosaurus , de Welles (1971). Aceste ichnotaxa prezintă o serie de amprente cu trei degete, care sunt în concordanță cu dimensiunea și forma așteptate a picioarelor pentru Dilophosaurus . [49] În 1991, Gerard Gierlinski, specialist în amprenta fosilă, a reexaminat urmele munților din Sfânta Cruce, Polonia și a redenumit urmele ca Kayentapus soltykovensis , concluzionând că forma „dilofosaurului” era cel mai potrivit candidat pentru atribuirea acestor icnotaxa. [50]
Alte amprente fosile datând de 200 de milioane de ani în roci sedimentare au fost atribuite Dilophosaurus și au fost găsite în formațiunea Höganäs din Vallåkra, Suedia în anii 1970. Amprentele par să arate că acești dinozauri au trăit în grupuri mici. [51] Amprentele fosile atribuite lui Dilophosaurus au fost descoperite și în Sala , tot în Suedia. Unele urme descoperite în formațiunea Höganäs au fost atribuite ichnogenus Grallator ( Eubrontes ) cf. giganteus , descoperit în straturile retice și Grallator ( Eubrontes ) soltykovensis , care au fost descoperite în straturile hetangiene . [52] Unele urme au fost aduse în muzee, dar cele mai multe au dispărut din păcate din cauza inundațiilor naturale. [53] În 1994, Gierlinski și Ahlberg au atribuit aceste piste de la formațiunea Hoganas din Suedia către Dilophosaurus . [54]

Gierlinski (1996) a observat câteva urme destul de neobișnuite asociate cu o pistă exemplară, din colecția de la Muzeul Pratt din Amherst , Massachusetts . Eșantionul AC 1/7 reprezintă urmele unui „dinozaur așezat”, realizat atunci când un dinozaur și-a așezat corpul pe pământ, lăsând impresia burticii între o pereche de urme. Unele urme asociate cu AC 1/7 au fost interpretate de Gierlinski ca amprente de pene , sugerând că Dilophosaurus era un dinozaur cu pene. [54] Cu toate acestea, o analiză suplimentară a arătat că liniile care arătau ca niște pene erau de fapt doar crăpături în noroiul în care s-a așezat animalul. Deși acest lucru nu exclude posibilitatea ca Dilophosaurus să fie parțial acoperit cu pene, nu există dovezi care să susțină acest lucru, ceea ce îl face doar speculații. [55]
Urmele fosile ale lui Eubrontes și Gigandipus , din Valea râului Connecticut , găsite atât în Connecticut, cât și în Massachusetts, au fost deseori atribuite lui Dilophosaurus , [56] [57] deși oasele fosile ale Dilophosaurus asociate cu urmele de picioare nu au fost niciodată găsite. Mărimea și forma sugerează că au fost făcute dintr-un teropod de aproximativ 6 metri lungime, similar cu dimensiunea Dilophosaurus , sugerând că amprentele aparțin lui Dilophosaurus sau unei rude apropiate. Două urme similare, numite Anchisauripus și Grallator , găsite și în Valea râului Connecticut, au fost deseori atribuite Dilophosaurus sau unei rude mai mici, precum Podokesaurus și Coelophysis . [56]
În cultura de masă
Dilophosaurus își datorează faima datorită apariției în romanul lui Michael Crichton Jurassic Park (1990) și adaptării sale cinematografice (1993). Cu toate acestea, Dilophosaurusul filmului și al romanului este deosebit de diferit: în carte, de fapt, are capacitatea de a scuipa otravă victimelor, orbindu-le (dacă este lovit în ochi) sau paralizându-le (foarte asemănătoare cu o scuipare) cobra ); în film, pe lângă faptul că este otrăvitor, are și un guler de piele retractabil în jurul gâtului (foarte asemănător cu cel al clamidosaurului ). El este responsabil pentru moartea lui Dennis Nedry , atacându-l în timp ce încerca să părăsească Isla Nublar. În film, Steven Spielberg a redus, de asemenea, dimensiunea Dilophosaurus la 3 picioare (0,91 metri) înălțime și 5 picioare (1,5 metri) în lungime, pentru a evita confuzia cu Velociraptorul filmului. [58] În ciuda succesului deosebit al acestei reconstrucții, nu există dovezi care să susțină prezența unui guler de piele sau a unui scuipat otrăvitor, [59] care au fost recunoscute de Crichton drept licență creativă. [60] Crichton însuși, în roman, explică faptul că nici biologii parcului nu au suspectat niciodată că Dilophosaurus este otrăvitor până când specimenele nu au fost observate urmărind șobolanii mușcându-i și așteptând ca veninul să intre în vigoare.
Cea mai recentă apariție în film a fost în Jurassic World , a patra tranșă a saga; apare pentru câteva secunde în centrul turistic și este reprodus ca o hologramă pentru a distrage atenția unui rapitor care îi urmărea pe protagoniști.
Succesul derivat din reconstrucția lui Spielberg, a însemnat că Dilophosaurus a apărut cu acest aspect în aproape toate aparițiile sale în filme , seriale TV și jocuri video , printre care putem include: Jurassic World , Jurassic Park: The Game , Ark: Survival Evolved , Primal Carnage , Ice Age 3 - Dawn of the Dinosaurs (joc video) și în Jurassic World Evolution .
Singurele apariții precise ale lui Dilophosaurus se află în documentarele Imperiul dinozaurilor și seria Preistorică .
Notă
- ^ Paul, GS, 2010, The Princeton Field Guide to Dinosaurs , Princeton University Press p. 75
- ^ a b c d e f Gay, Robert, New specimens of Dilophosaurus wetherilli (Dinosauria: Theropoda) from the Early Jurassic Kayenta Formation of northern Arizona , in Western Association of Vertebrate Paleontologists annual meeting volume Mesa, Arizona , vol. 1, 2001, p. 1.
- ^ a b c d e f g h i j k Welles, SP, Dilophosaurus wetherilli (Dinosauria, Theropoda), osteologie și comparații , în Paleontogr. Abt. A , vol. 185, 1984, pp. 85-180.
- ^ a b Norman, David , The Illustrated Encyclopedia of Dinosaurs , New York, Crescent Books, 1985, pp. 62-67, ISBN 0-517-46890-5 .
- ^ Gay, Robert, Dovezi pentru dimorfism sexual în dinozaurul teropod din Jurassic timpuriu, Dilophosaurus și o comparație cu alte forme conexe În: Carpenter, Ken, ed. The Carnivorous Dinosaurs , în The Carnivorous Dinosaurs , Indiana University Press, 2005, pp. 277 -283, ISBN 0-253-34539-1 .
- ^ Carrano, Benson și Sampson, 2012. Filogenia Tetanurei (Dinosauria: Theropoda). Jurnal de paleontologie sistematică. 10 (2), 211-300.
- ^ Rauhut, 2000. Relațiile și evoluția teropodelor bazale (Dinosauria, Saurischia). Teză de doctorat, Universitatea Bristol [Marea Britanie]. 440 pp.
- ^ a b Welles, SP, [591: NJDFTK 2.0.CO; 2 Nou dinozaur jurasic din formațiunea Kayenta din Arizona ], în Buletinul Societății Geologice din America , vol. 65, nr. 6, 1954, pp. 591-598, DOI : 10.1130 / 0016-7606 (1954) 65 [591: NJDFTK] 2.0.CO; 2 .
- ^ a b Dodson, P. (1997) Paleoecology In: Encyclopedia of Dinosaurs, editat de Currie, PJ și Padian, K., Academic Press.
- ^ Glut, DF, 2006, Dinosaurs, Encyclopedia, Supliment 4: McFarland & Company, Inc, 749pp.
- ^ Padian, K., Horner, JR și Dhaliwal, J. 2004. Recunoașterea speciilor ca fiind cauza principală a structurilor bizare la dinozauri. Journal of Vertebrate Paleontology 23 (3): 100A.
- ^ a b Milner ARC, Harris JD, Lockley MG, Kirkland JI, Matthews NA (2009) Anatomie, postură și comportament asemănător păsărilor, dezvăluit de o urmă de odihnă dinozaur cu un teropod jurassic timpuriu . PLoS ONE 4 (3) (2009): e4591.
- ^ MT Carrano, RBJ Benson și SD Sampson. 2012. Filogenia Tetanurei (Dinosauria: Theropoda). Jurnal de paleontologie sistematică 10 (2): 211-300 [M. Carrano / M. Carrano]
- ^ Yates, 2005. Un nou dinozaur teropod din Jurasicul timpuriu al Africii de Sud și implicațiile sale pentru evoluția timpurie a teropodelor. Paleontologia africană. 41, 105–122.
- ^ Smith, Makovicky, Hammer și Currie, 2007. Osteologia Cryolophosaurus ellioti (Dinosauria: Theropoda) din Jurasicul timpuriu al Antarcticii și implicații pentru evoluția timpurie a teropodului. Jurnalul Zoologic al Societății Linnean. 151, 377-421.
- ^ Hendrickx, C., Hartman, SA și Mateus, O. (2015). O prezentare generală a descoperirilor și clasificării teropodelor non-aviare. Jurnalul de paleontologie al vertebratelor PalArch , 12 (1): 1-73.
- ^ Welles, Sam, Dilophosaurus Discovered , la ucmp.berkeley.edu , Universitatea din California, Berkeley , 2007. Accesat la 17 noiembrie 2007 (arhivat din original la 8 noiembrie 2017) .
- ^ a b Welles, Sam, Dilophosaurus Details , ucmp.berkeley.edu , Universitatea din California, Berkeley , 2007. Accesat la 17 noiembrie 2007 .
- ^ a b Michael J. Benton, Prehistoric Life , Edinburgh, Scoția, Dorling Kindersley, 2012, p. 258 , ISBN 978-0-7566-9910-9 .
- ^ Irmis, Randall, First Report of Megapnosaurus from China ( PDF ), în PaleoBios , vol. 24, n. 3, 22 decembrie 2004, pp. 11-18 (arhivat din original la 17 decembrie 2008) .
- ^ Hu, Shaojin, A Short Report on the Occurrence of Dilophosaurus from Jinning County, Yunnan Province , in Vertebrata PalAsiatica , vol. 31, 1993, pp. 65-69.
- ^ Lamanna, MC, Holtz, TR Jr, and Dodson, P., 1998, A reassessment of the Chinese Theropod Dinosaur Dilophosaurus sinensis: Journal of Vertebrate Paleontology, Volume 18, Supplement to Number 3. Abstracts of papers. Fifty-eighth annual meeting, Society of Vertebrate Paleontology, Snowbird Ski and Summer Resort, Snowbird, Utah, September 30 – October 3, 1998, p. 57a.
- ^ L. Xing, PR Bell, BM Rothschild, H. Ran, J. Zhang, Z. Dong, W. Zhang e PJ Currie, Tooth loss and alveolar remodeling in Sinosaurus triassicus (Dinosauria: Theropoda) from the Lower Jurassic strata of the Lufeng Basin, China , in Chinese Science Bulletin , 2013, DOI : 10.1007/s11434-013-5765-7 .
- ^ Tykoski, 2005. Anatomy, ontogeny and phylogeny of coelophysoid theropods. PhD Dissertation. University of Texas at Austin. 553 pp.
- ^ Mickey Mortimer, Coelophysoidea , su archosaur.us , 2012 (archiviato dall' url originale il 4 maggio 2013) .
- ^ George Olshevsky, Dinosaur Genera List corrections #126 , su Dinosaur Mailing List Archives , Cleveland Museum of Natural History, 5 dicembre 1999. URL consultato il 25 giugno 2008 .
- ^ Tykoski, RS & Rowe, T. (2004). "Ceratosauria". In: Weishampel, DB, Dodson, P., & Osmolska, H. (Eds.) The Dinosauria (2nd edition). Berkeley: University of California Press. Pp. 47–70 ISBN 0-520-24209-2
- ^ Bradley, OC 1903. The muscles of mastication and movements of the skull in Lacertilia. Zoologische Jahrbiicher, Anatomie 18: 475–486.
- ^ A. Milner e J. Kirkland, The case for fishing dinosaurs at the St. George Dinosaur Discovery Site at Johnson Farm ( PDF ), in Survey Notes of the Utah Geological Survey , vol. 39, 2007, pp. 1-3.
- ^ Tkach, JS, 1996, Multi-element osteohistological analysis of Dilphosaurus wetherilli : Journal of Vertebrate Paleontology, v. 16, supplement to n. 3, Abstracts of Papers, Fifty-sixth Annual Meeting, Society of Vertebrate Paleontology, American Museum of Natural History, New York, New York, October 16–19.
- ^ Molnar, RE, 2001, Theropod paleopathology: a literature survey: In: Mesozoic Vertebrate Life, edited by Tanke, DH, and Carpenter, K., Indiana University Press, p. 337-363.
- ^ http://journals.plos.org/plosone/article?id=10.1371%2Fjournal.pone.0149140
- ^ Rothschild, B., Tanke, DH, and Ford, TL, 2001, Theropod stress fractures and tendon avulsions as a clue to activity: In: Mesozoic Vertebrate Life, edited by Tanke, DH, and Carpenter, K., Indiana University Press, p. 331-336.
- ^ a b JM Clark and DE Fastovsky. 1986. Vertebrate biostratigraphy of the Glen Canyon Group in northern Arizona. The Beginning of the Age of the Dinosaurs: Faunal change across the Triassic-Jurassic boundary, NC Fraser and H.-D. Sues (eds.), Cambridge University Press 285–301
- ^ a b JW Harshbarger, CA Repenning e JH Irwin, Stratigraphy of the uppermost Triassic and the Jurassic rocks of the Navajo country , Professional Paper, vol. 291, Washington, DC, US Geological Survey, 1957.
- ^ Padian, K (1997) Glen Canyon Group In: Encyclopedia of Dinosaurs, edited by Currie, PJ, and Padian, K., Academic Press.
- ^ a b c SG Lucas, AB Heckert e LH Tanner, Arizona's Jurassic fossil vertebrates and the age of the Glen Canyon Group , in Heckert, AB e Lucas, SG (a cura di), Vertebrate paleontology in Arizona , Bulletin, vol. 29, Albuquerque, NM, New Mexico Museum of Natural History and Science, 2005, pp. 95-104.
- ^ a b c Luttrell, PR, and Morales, M. 1993. Bridging the gap across Moenkopi Wash: a lithostratigraphic correlation. Aspects of Mesozoic geology and paleontology of the Colorado Plateau. Pages 111–127 in Morales, M., editor. Museum of Northern Arizona, Flagstaff, AZ. Bulletin 59.
- ^ a b Jenkins, FA, Jr. and Shubin, NH 1998. Prosalirus bitis and the anuran caudopelvic mechanism. Journal of Vertebrate Paleontology 18 (3):495–510.
- ^ Rigby, JK, Hamblin, WK, Matheny, R., and Welsh, SL 1971. Guidebook to the Colorado river: part 3, Moab to Hite, Utah through Canyonlands National Park. Brigham Young University Research Studies, Geology Series 18 (2).
- ^ Lucas, SG, and Tanner LH 2007. Tetrapod biostratigraphy and biochronology of the Triassic-Jurassic transition on the southern Colorado Plateau, USA. Palaeogeography, Palaeoclimatology, Palaeoecology 244 (1–4):242–256.
- ^ Curtis, K., and Padian, K. 1999. An Early Jurassic microvertebrate fauna from the Kayenta Formation of northeastern Arizona: microfaunal change across the Triassic-Jurassic boundary. PaleoBios 19 (2):19–37.
- ^ a b Jenkins, FA, Jr., Crompton, AW, and Downs, WR 1983. Mesozoic mammals from Arizona: new evidence in mammalian evolution. Science 222 (4629):1233–1235.
- ^ Gay, R. 2010. Kayentavenator elysiae , a new tetanuran from the early Jurassic of Arizona. Pages 27–43 in Gay, R. Notes on early Mesozoic theropods. Lulu Press (on-demand online press).
- ^ Rowe, TB, Sues, H.-D., and Reisz, RR 2011. Dispersal and diversity in the earliest North American sauropodomorph dinosaurs, with a description of a new taxon. Proceedings of the Royal Society B: Biological Sciences 278 (1708):1044–1053.
- ^ Hamblin, AH, and Foster, JR 2000. Ancient animal footprints and traces in the Grand Staircase-Escalante National Monument, south-central Utah. Pages 557–568 in Sprinkel, DA, Chidsey, TC, Jr., and Anderson, PB editors. Geology of Utah's parks and monuments. Utah Geological Association, Salt Lake City, UT. Publication 28.
- ^ Tykoski, RS, 1998, The Osteology of Syntarsus kayentakatae and its Implications for Ceratosaurid Phylogeny: Theses, The University of Texas, December 1998.
- ^ TS Kutty, S. Chatterjee, PM Galton e P. Upchurch, Basal Sauropodomorphs (Dinosauria: Saurischia) from the Lower Jurassic of India: Their Anatomy and Relationships , in Journal of Paleontology , vol. 81, n. 6, 2007, p. 1218, DOI : 10.1666/04-074.1 .
- ^ Welles, SP (1971). Dinosaur footprints from the Kayenta Formation of northern Arizona: Plateau, v. 44, pp. 27–38.
- ^ Gierlinski, G.(1991) New dinosaur ichnotaxa from the Early Jurassic of the Holy Cross Mountains, Poland. Palaeogeogr., Palaeoclimat.,Palaeoecol.,85(1–2): 137–148
- ^ Copia archiviata , su exempelbanken.se . URL consultato il 25 gennaio 2017 (archiviato dall' url originale il 20 ottobre 2013) .
- ^ G. Gierliński e A. Ahlberg, Late triassic and early jurassic dinosaur footprints in the höganäs formation of southern Sweden , in Ichnos , vol. 3, n. 2, 1994, p. 99, DOI : 10.1080/10420949409386377 .
- ^ Kent Lungren, Tankar i tiden: Kontakt med Trias och Jura , su kentlundgren.blogspot.se . URL consultato il 23 ottobre 2013 .
- ^ a b Glut, DF (1999). Dinosaurs, the Encyclopedia, Supplement 1: McFarland & Company, Inc., 442pp.
- ^ Martin, AJ & Rainforth, EC 2004. A theropod resting trace that is also a locomotion trace: case study of Hitchcock's specimen AC 1/7. Geological Society of America, Abstracts with Programs 36 (2), 96.
- ^ a b Dinosaur footprints of the Connecticut River Valley , su nashdinosaurtracks.com , Nash Dinosaur Track Site and Rock Shop. URL consultato il 25 gennaio 2017 (archiviato dall' url originale il 1º gennaio 2017) .
- ^ 10 Crested Facts About Dilophosaurus , su mentalfloss.com , Mental Floss.
- ^ The Making of Jurassic Park by Don Shay & Jody Duncan, Boxtree Ltd; 1st Edition. edition (30 Jun 1993), ISBN 1-85283-774-8
- ^ Bennington, J Bret, Errors in the Movie "Jurassic Park" , in American Paleontologist , vol. 4, n. 2, 1996, pp. 4-7.
- ^ Michael Crichton , Jurassic Park , Alfred A. Knopf , 1990, ISBN 0-394-58816-9 .
Bibliografia
- Welles, SP 1954. New Jurassic dinosaur from the Kayenta Formation of Arizona . Bulletin of the Geological Society of America 65:591-598.
- Welles, SP 1970. Dilophosaurus (Reptilia: Saurischia), a new name for a dinosaur . Journal of Paleontology 44:200.
Altri progetti
-
 Wikimedia Commons contiene immagini o altri file su Dilophosaurus wetherilli
Wikimedia Commons contiene immagini o altri file su Dilophosaurus wetherilli -
 Wikispecies contiene informazioni su Dilophosaurus wetherilli
Wikispecies contiene informazioni su Dilophosaurus wetherilli
Collegamenti esterni
- ( EN ) scheda sul Dilophosaurus , su internt.nhm.ac.uk . URL consultato il 19 luglio 2008 (archiviato dall' url originale il 10 settembre 2008) .
- ( EN ) sito dedicato completamente al Dilophosaurus , su ucmp.berkeley.edu . URL consultato il 19 luglio 2008 (archiviato dall' url originale il 14 aprile 2009) .
- BENVENUTI alla Dilophosauruspage , su web.tiscali.it .