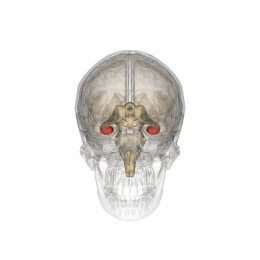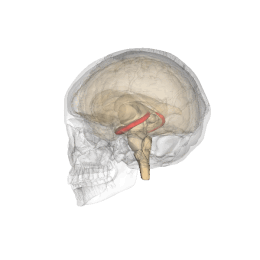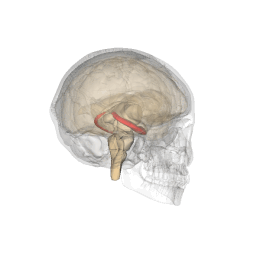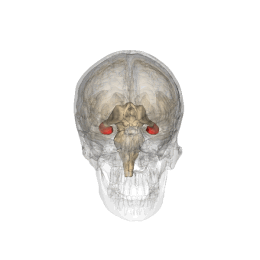
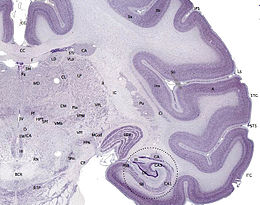
Hipocamp (anatomie)
| Hipocamp | |
|---|---|
 | |
 | |
| Nume latin | Hipocamp |
| Localizare anatomică | lobul temporal |
| Identificatori | |
| Plasă | A08.186.211.180.405 și A08.186.211.200.885.287.500.345 |
| TA | A14.1.09.321 |
| FMA | 275020 |
| ID NeuroLex | birnlex_721 |
Hipocampul face parte din creier , situat în regiunea interioară a lobului temporal . Face parte din formarea hipocampului , inserat în sistemul limbic și joacă un rol important în formarea amintirilor explicite (declarative și semantice), în transformarea memoriei pe termen scurt în memorie pe termen lung și în navigația spațială. . Hipocampul este situat sub cortexul cerebral în alocortex . [1] [2] iar la primate se află în lobul temporal medial. Conține două părți principale care se blochează: hipocampul propriu-zis (numit și cornul lui Ammon) [3] și girusul dentat .
Oamenii și alte mamifere au doi hipocampi, unul în fiecare emisferă a creierului. La rozătoare, animale la care hipocampul a fost studiat pe larg, hipocampul are aproximativ forma unei banane. La om, are o formă curbată și complicată, care a inspirat imaginea unui cal de mare primilor anatomiști. Numele, de fapt, derivă din greacă ( greacă : hipopotami = cal , kàmpe = omidă ).
În boala Alzheimer , hipocampul este una dintre primele regiuni ale creierului care suferă daune; afectarea memoriei și dezorientarea sunt primele simptome care apar. Leziunile hipocampice pot apărea și ca urmare a lipsei de oxigen ( anoxie ), a encefalitei sau a epilepsiei lobului temporal medial. Persoanele cu leziuni extinse ale țesutului hipocampic pot prezenta amnezie anterogradă , care este o incapacitate de a forma sau menține noi amintiri.
Deoarece diferite tipuri de celule neuronale sunt bine organizate în straturi în hipocampus, a fost adesea folosit ca sistem model pentru studierea neurofiziologiei. Plasticitatea neuronală cunoscută sub numele de potențare pe termen lung (LTP) a fost descoperită inițial în hipocamp și a fost adesea studiată în această structură. LTP este considerat a fi unul dintre mecanismele neuronale majore prin care amintirile sunt stocate în creier.
Istorie
Anatomistul Giulio Cesare Aranzi (în jurul anului 1564) a folosit prima dată termenul de hipocamp pentru a descrie acest organ cerebral datorită asemănării sale cu calul de mare. Acest organ a fost inițial legat de simțul mirosului și nu de funcțiile sale în formarea urmelor de memorie. În jurul anului 1900, rusul Vladimir Bechterev a remarcat rolul hipocampului în funcțiile de memorie, pe baza observării unui pacient cu insuficiență severă a memoriei. Cu toate acestea, timp de câțiva ani, rolul atribuit hipocampului a fost, ca și restul sistemului limbic , acela de a fi responsabil pentru emoții.
Importanța hipocampului în procesele de memorie a fost adusă în atenția cercetătorilor de către pacientul HM . HM a suferit de mai multe deficite de memorie anterograde și cronologice (astfel de tulburări au devenit subiectul filmelor) în urma îndepărtării bilaterale a mai multor structuri ale lobului temporal medial (inclusiv ablația bilaterală a hipocampilor) pentru a preveni convulsiile frecvente. Relevantă a fost observația că HM era încă capabil să învețe sarcini procedurale (asociate cu țesut striat ) și că IQ-ul său a rămas peste medie. Cazul HM a demonstrat disocierea dintre inteligență și memoria declarativă. Mărimea relativă a formațiunii hipocampice în raport cu volumul total al creierului este adesea conservată la majoritatea speciilor de mamifere, deși s-a găsit o anumită hipotrofie la cetacee.
Anatomie

Anatomic, hipocampul este o elaborare a marginii cortexului. Poate fi distins ca o zonă în sine, unde cortexul se subțiază într-un singur strat de neuroni dens organizați, care se pliază pentru a forma un S. foarte strâns. Structura care se aliniază cu marginea cortexului formează așa-numitul sistem limbic (din latinescul limbus = limită ): acesta include hipocampul, cortexul cingulat, cortexul olfactiv și amigdala. Paul MacLean a sugerat în teoria sa Triunbrain că structurile limbice formează baza neuronală a emoțiilor. Cu toate acestea, majoritatea neurologilor nu mai consideră valid conceptul de „sistem limbic unificat”.
Fiziologie
Hipocampul prezintă două „moduri” principale de activitate, fiecare asociat cu un model distinct al undelor EEG și al activității populației neuronale. Aceste moduri sunt numite după modelele EEG asociate acestora: theta și LIA (activitate neregulată mare). Iată câteva dintre caracteristicile lor majore la șobolan, cel mai studiat animal: [4]
Hipocampul pare să aibă o poziție importantă în învățarea spațială (în emisfera dreaptă) și în învățarea semantică (în emisfera stângă). La șobolan s-a observat prezența celulelor locului care, în ceea ce privește câmpurile receptive, par să se activeze după ocuparea unei regiuni spațiale specifice.

Hipocampul poate fi considerat ca un „spațiu de memorie” în care informațiile multisenzoriale legate de o memorie declarativă (episodică și semantică) s-ar integra pentru o perioadă scurtă de timp. Ulterior vor fi trimiși în regiuni parahippocampale care, prin disocierea lor, le vor face o amintire mai durabilă.
Funcțiile hipocampului
Poate că prima ipoteză a presupus hipocampul implicat în olfacție, o ipoteză sugerată în principal de localizarea sa în creier, lângă cortexul olfactiv. Există în continuare un anumit interes în implicarea hipocampului în funcțiile olfactive, dar funcția principală a hipocampului nu este simțul mirosului.
De-a lungul anilor au existat trei idei dominante despre funcția hipocampului: inhibiție, memorie și spațiu. Teoria inhibiției comportamentale (definită ironic de O'Keefe și Nadel ca „un picior pe frână”), populară până în anii 1960, a apărut din două observații: În primul rând, animalele al căror hipocamp a fost deteriorat au avut tendința de a fi hiperactive; în al doilea rând, că animalele cu leziuni ale țesutului hipocampic au avut deseori dificultăți de învățare pentru a inhiba răspunsurile pe care le-au fost predate anterior. Jeffrey Gray a dezvoltat această linie de gândire într-o teorie reală asupra rolului hipocampului în stările de anxietate [6] . Teoria inhibiției nu este larg considerată astăzi, deoarece această funcție și alte funcții sunt atribuite în prezent amigdalei , o structură anatomic apropiată de hipocamp.
A doua linie importantă de gândire asociază funcția hipocampului cu memoria. Cel mai important este cuprins în celebrul tratat de Scoville și Milner [7] privind consecințele distrugerii chirurgicale a hipocampului (în încercarea de a elimina convulsiile de epilepsie), observate la un pacient numit HM . Pacientul a prezentat amnezie severă și nu și-a amintit ce i s-a întâmplat după operație sau evenimentele petrecute anterior, chiar și pe o perioadă de ani. Acest lucru a stârnit un astfel de interes, încât HM este cel mai studiat caz medical din istorie. În anii următori, au fost studiați cu aceeași intensitate alți pacienți cu disfuncții de memorie similare legate de leziunile hipocampice (datorate accidentelor sau malformațiilor congenitale) și s-au efectuat mii de experimente privind fiziologia plasticității neuronale în hipocamp. Astăzi nu există nici o diferență cu privire la importanța hipocampului, considerat universal ca sediul memoriei. Cu toate acestea, rolul specific pe care îl joacă în raport cu această funcție psihică rămâne încă o chestiune de dezbatere. [8] [9] .
Rol în memoria generală
Psihologii și neurologii sunt de acord în general că hipocampul joacă un rol important în formarea de noi amintiri privind evenimentele trăite ( memoria episodică sau memoria autobiografică ) [9] [10] . Unii cercetători preferă să considere hipocampul ca parte a unui sistem mnemonic mai mare al lobului temporal medial , responsabil în general pentru memoria declarativă (amintiri care pot fi verbalizate explicit ca memorie semantică, precum și ca memorie episodică) [8] .
Unele dovezi susțin ipoteza că, deși unele forme de memorie pot persista pe tot parcursul vieții, hipocampul încetează să mai joace un rol crucial în păstrarea memoriei după o perioadă de consolidare [11] . Deteriorarea hipocampului duce în general la dificultăți severe în formarea de noi amintiri ( amnezie anterogradă ) și accesul la amintiri înainte de deteriorare este, de asemenea, afectat în mod normal ( amnezie retrogradă ). Deși efectul retrograd se poate extinde cu câțiva ani înainte de avarie, în unele cazuri rămân amintirile mai îndepărtate - această salvare a amintirilor mai vechi a condus la ideea că consolidarea în timp implică transferul amintirilor din hipocamp, în alte părți ale creierului. Cu toate acestea, experimentarea întâmpină mari dificultăți în măsurarea persistenței acestor amintiri mai vechi; în plus, în unele cazuri de amnezie retrogradă, această persistență pare să implice amintiri formate cu zeci de ani înainte de apariția leziunii hipocampice, astfel încât rolul său în menținerea acestor amintiri îndepărtate rămâne îndoielnic.
Deteriorarea hipocampului nu are niciun efect asupra unor aspecte ale memoriei, cum ar fi capacitatea de a dobândi noi abilități motorii (de exemplu, a cânta la un instrument muzical): acest lucru sugerează că aceste tipuri de abilități depind de un alt tip de memorie ( memoria procedurală ) și de diferite regiuni ale creierului. Există, de asemenea, dovezi că pacientul HM (ai cărui lobi temporali mediali au fost eliminați bilateral ca tratament pentru epilepsie [7] ) este capabil să formeze o nouă memorie semantică . [12]
Electroencefalografie
Hipocampul prezintă două „moduri” principale de activitate, fiecare asociat cu un model distinct de activitate a populației neuronale și valuri de activitate electrică măsurate printr-o electroencefalogramă (EEG). Aceste moduri sunt numite după modelele EEG asociate acestora: unde theta și activitate neregulată mare (LIA). Principalele caracteristici descrise mai jos sunt pentru șobolan, care este cel mai studiat animal. [13]

Modul Theta apare în timpul stărilor de comportament activ, alert (în special locomoție), precum și în timpul somnului REM (visare). [14] În modul teta, EEG este dominat de unde regulate mari cu o frecvență cuprinsă între 6 și 9 Hz, iar principalele grupuri de neuroni hipocampici (celule piramidale și granule) prezintă o activitate slabă a populației, ceea ce înseamnă că în orice pe o perioadă scurtă de timp, marea majoritate a celulelor sunt silențioase, în timp ce fracția mică rămasă trage la viteze relativ mari, până la 50 de vârfuri pe secundă pentru cele mai active. O celulă activă rămâne activă de obicei timp de o jumătate de secundă până la câteva secunde. Pe măsură ce șobolanul se mișcă, celulele active devin tăcute și noile celule devin active, dar procentul total de celule active rămâne mai mult sau mai puțin constant. În multe situații, activitatea celulară este în mare măsură determinată de localizarea spațială a animalului, dar alte variabile comportamentale îl influențează în mod clar.
Modul LIA apare în timpul somnului cu undă lentă (nu visează) și, de asemenea, în timpul stărilor de liniște de veghe, cum ar fi odihna sau mâncarea. [15] În modul LIA, EEG este dominat de unde acute, care sunt devieri mari temporizate ale semnalului EEG, care durează 25-50 de milisecunde. Undele ascuțite sunt adesea generate în serie, cu serii care conțin până la 5 sau mai multe unde individuale și durează până la 500 ms. Activitatea de spionaj a neuronilor din cadrul hipocampului este puternic corelată cu activitatea undelor acute. Majoritatea neuronilor scad rata de tragere între valurile acute; cu toate acestea, în timpul unui val puternic, există o creștere dramatică a ratei de tragere de până la 10% din populația hipocampală
Aceste două moduri de activitate hipocampală pot fi observate atât la primate, cât și la șobolani, cu excepția faptului că ritmicitatea teta robustă în hipocampul primat a fost dificil de văzut. Există, totuși, valuri acute calitativ similare și modificări ale valurilor dependente de stat în activitatea populației neuronale.
Fiziopatologie
Boala Alzheimer
Condițiile legate de vârstă, cum ar fi boala Alzheimer și alte forme de demență (pentru care afectarea hipocampului este unul dintre primele semne [16] ) au un impact sever asupra multor tipuri de cunoaștere, inclusiv memoria. Îmbătrânirea normală este, de asemenea, asociată cu o scădere treptată a anumitor tipuri de memorie, inclusiv memoria episodică și memoria de lucru (sau memoria pe termen scurt). Din moment ce se crede că hipocampul joacă un rol central în memorie, a existat un interes considerabil în posibilitatea ca declinul legat de vârstă să fie cauzat de deteriorarea hipocampului. [17] Unele studii timpurii au raportat pierderi neuronale substanțiale în hipocampus la vârstnici, dar studii ulterioare folosind tehnici mai precise au constatat doar diferențe minime. [17] În mod similar, unele studii RMN au raportat o îngustare a hipocampului la vârstnici, dar alte studii nu au reușit să reproducă această constatare. Există, totuși, o relație fiabilă între dimensiunea hipocampului și performanța memoriei; astfel încât acolo unde există o reducere legată de vârstă, performanța memoriei va fi compromisă. [18] Există, de asemenea, dovezi că activitățile de memorie tind să producă mai puțină activare a hipocampului la vârstnici decât la tineri. [18] În plus, un studiu de control randomizat publicat în 2011 a constatat că exercițiile aerobe ar putea crește dimensiunea hipocampusului la adulți cu vârste cuprinse între 55 și 80 de ani și, de asemenea, ar putea îmbunătăți memoria spațială.
În boala Alzheimer, cortexul asociativ: temporal, prefrontal și parietal, este locul depunerii anormale a β-amiloidului . Pe de altă parte, depunerea proteinelor Tau anormal de hiperfosforilate se găsește în extensiile neuronale. Acestea formează grupuri neurofibrilare găsite în cantități mari în regiunile interne ale lobului temporal și în special în hipocamp. Neuronii ale căror extensii au fost locul clusterelor neurofibrilare se pierd. Acest lucru duce la atrofierea regiunilor afectate.
Stres
Hipocampul conține niveluri ridicate de receptori glucocorticoizi , ceea ce îl face mai vulnerabil la stres pe termen lung decât majoritatea celorlalte zone ale creierului. [19] Există dovezi că oamenii care au suferit stres traumatic sever și de lungă durată prezintă atrofie a hipocampului mai mult decât alte părți ale creierului. Aceste efecte apar în PTSD [20] și pot contribui la atrofia hipocampală raportată în schizofrenie [21] și la depresie severă. [22] Un studiu recent a relevat atrofia cauzată de depresie, dar aceasta poate fi oprită cu antidepresive chiar dacă acestea nu sunt eficiente în ameliorarea altor simptome. [22]
Stresul cronic care determină niveluri crescute de glucocorticoizi, în special cortizolul, este considerat o cauză de atrofie neuronală în hipocamp. Această atrofie are ca rezultat un volum mai mic al hipocampului, care se observă și în sindromul Cushing. Nivelurile mai ridicate de cortizol din sindromul Cushing sunt de obicei rezultatul administrării medicamentelor pentru alte afecțiuni. [23] Pierderea neuronală apare și din cauza neurogenezei afectate. Un alt factor care contribuie la reducerea volumului hipocampului este cel al retracției dendritice în care dendritele sunt scurtate în lungime și reduse ca număr, ca răspuns la creșterea glucocorticoizilor. Această retragere dendritică este reversibilă. [24] După tratamentul cu medicamente pentru reducerea cortizolului în sindromul Cushing, volumul hipocampului este restabilit cu până la 10%. Această schimbare se datorează schimbării dendritelor. [24] Această restaurare dendritică poate apărea și atunci când stresul este eliminat. Există totuși dovezi derivate în principal din studii efectuate la șobolani că stresul care apare la scurt timp după naștere poate afecta funcția hipocampului în moduri care persistă pe tot parcursul vieții.
La șobolani, s-a dovedit că răspunsurile sexuale specifice la stres au un efect asupra hipocampului. Stresul cronic la șobolanul mascul a arătat retracție dendritică și pierdere celulară în regiunea CA3, dar acest lucru nu a fost demonstrat la femelă. Se credea că acest lucru se datorează hormonilor ovarieni neuroprotectori. [25] La șobolani, daunele ADN cresc în hipocamp în condiții de stres. [26]
Epilepsie

Hipocampul este una dintre puținele regiuni ale creierului în care sunt generați noi neuroni. Acest proces de neurogeneză este limitat la girusul dentat. [27] Producția de noi neuroni poate fi influențată pozitiv de exerciții fizice sau negativ de crize epileptice. [27]

Convulsiile în epilepsia lobului temporal pot afecta dezvoltarea normală a noilor neuroni și pot provoca leziuni tisulare. Scleroza hipocampală este cel mai frecvent tip de astfel de leziuni tisulare. [28] Cu toate acestea, este încă neclar dacă epilepsia este de obicei cauzată de anomalii ale hipocampului sau dacă hipocampul este afectat de efectele cumulative ale convulsiilor. [29] Cu toate acestea, în medii experimentale în care convulsiile sunt induse artificial la animale, deteriorarea hipocampului este un rezultat frecvent. Aceasta poate fi o consecință a concentrației receptorilor de glutamat excitabili din hipocamp. Hiperexcitabilitatea poate duce la citotoxicitate și moarte celulară. Ar putea avea, de asemenea, de-a face cu hipocampul, care este un loc în care neuronii continuă să fie creați de-a lungul vieții și anomalii în acest proces. [30]
Notă
- ^ (EN) John Martin, Neuroanatomy: Text and Atlas , McGraw Hill Professional, 2003. ISBN 978-0-07-138183-3 . Adus la 30 aprilie 2020 .
- ^ (EN) Pentru Andersen, Richard Morris și David Amaral, The Hippocampus Book , Oxford University Press, SUA, 2007, ISBN 978-0-19-510027-3 . Adus la 30 aprilie 2020 .
- ^ J PEARCE, cornul lui Ammon și hipocampul , în Jurnalul de neurologie, neurochirurgie și psihiatrie , vol. 71, nr. 3, 2001-9, p. 351, DOI : 10.1136 / jnnp.71.3.351 . Adus la 30 aprilie 2020 .
- ^ Vezi bibliografia: Buzsaki, 2006
- ^ Vezi bibliografia: Skaggs și colab., 1996
- ^ Vezi bibliografia: Gray și McNaughton, 2000
- ^ a b A se vedea bibliografia: Scoville și Milner, 1957
- ^ a b A se vedea bibliografia: Squire, 1992
- ^ a b A se vedea bibliografia: Eichenbaum și Cohen, 1993
- ^ Vezi bibliografia: Squire și Schacter, 2002
- ^ Vezi bibliografia: Squire și Schacter, 2002, Cap. 1
- ^ Vezi bibliografia: O'Kane și colab., 2004
- ^ Buzsáki, G., Rhythms of the brain , Oxford University Press, 2006, ISBN 978-0-19-804125-2 ,OCLC 71338388 . Adus la 30 aprilie 2020 .
- ^ (EN) György Buzsáki, S. Chen Lan și Fred H. Gage, Progresul în cercetarea creierului , în Înțelegerea creierului prin hipocampul hipocampului regiunea ca model pentru studierea structurii și funcției creierului, vol. 83, Elsevier, 1 ianuarie 1990, pp. 257-268. Adus la 30 aprilie 2020 .
- ^ (EN) György Buzsáki, Theta Oscillations in the Hippocampus , în Neuron, vol. 33, nr. 3, 31 ianuarie 2002, pp. 325-340, DOI : 10.1016 / S0896-6273 (02) 00586-X . Adus la 30 aprilie 2020 .
- ^ Harald Hampel, Katharina Bürger și Stefan J. Teipel, biomarkeri neurochimici și imagini candidați principali ai bolii Alzheimer , în Alzheimer și demență , vol. 4, nr. 1, 1 ianuarie 2008, pp. 38-48, DOI : 10.1016 / j.jalz.2007.08.006 . Adus la 30 aprilie 2020 .
- ^ a b Prull MW, Gabrieli JD, Bunge SA (2000). "Ch 2. Modificări legate de vârstă în memorie: O perspectivă a neuroștiinței cognitive". În Craik FI, Salthouse TA (eds.). Manualul îmbătrânirii și al cunoașterii. Erlbaum .
- ^ a b ( EN ) F. Rodrıйguez, JC López și JP Vargas, Memoria spațială și paliumul hipocampic prin evoluția vertebratelor: perspective de pe reptile și pești teleostici , în Brain Research Bulletin , vol. 57, nr. 3, 1 februarie 2002, pp. 499-503, DOI : 10.1016 / S0361-9230 (01) 00682-7 . Adus la 30 aprilie 2020 .
- ^ (EN) Marian Joels, Acțiuni funcționale ale corticosteroizilor din hipocamp , în European Journal of Pharmacology, vol. 583, nr. 2, 7 aprilie 2008, pp. 312-321, DOI : 10.1016 / j.ejphar.2007.11.064 . Adus la 30 aprilie 2020 .
- ^ (EN) Anke Karl, Michael Schaefer și Loretta S. Malta, O meta-analiză a anomaliilor structurale ale creierului în PTSD , în Neuroscience & Biobehavioral Reviews, vol. 30, n. 7, 1 ianuarie 2006, pp. 1004-1031, DOI :10.1016 / j.neubiorev.2006.03.004 . Adus la 30 aprilie 2020 .
- ^ Ian C. Wright, Sophia Rabe-Hesketh și Peter WR Woodruff, Meta-Analysis of Regional Brain Volumes in Schizophrenia , în American Journal of Psychiatry , vol. 157, nr. 1, 1 ianuarie 2000, pp. 16-25, DOI : 10.1176 / ajp.157.1.16 . Adus la 30 aprilie 2020 .
- ^ a b ( EN ) Matthew J. Kempton, Zainab Salvador și Marcus R. Munafò, Structural Neuroimaging Studies in Major Depressive Disorder: Meta-analysis and Comparation With Bipolar Disorder , in Archives of General Psychiatry , vol. 68, nr. 7, 4 iulie 2011, pp. 675-690, DOI : 10.1001 / arhigenpsihiatrie.2011.60 . Adus la 30 aprilie 2020 .
- ^ (EN) Forumul Institutului de Medicină (SUA) privind neuroștiința și tulburările sistemului nervos, Prezentare generală a sistemului glutamatergic , National Academies Press (SUA), 2011. Accesat la 30 aprilie 2020.
- ^ a b Garcia-Segura, Luis Miguel., Hormonii și plasticitatea creierului , Oxford University Press, 2009, ISBN 978-0-19-971682-1 ,OCLC 373185473 . Adus la 30 aprilie 2020 .
- ^ Cheryl D. Conrad, Vulnerabilitatea hipocampală indusă de stresul cronic: ipoteza vulnerabilității glucocorticoide , în Review in the neurosciences , vol. 19, nr. 6, 2008, pp. 395-411. Adus la 30 aprilie 2020 .
- ^ (RO) Consiliul AR, allp Ramos și JAP Henriques, leziuni cerebrale ale ADN-ului după stres la șobolani , în progres în Neuro-Psihofarmacologie și Psihiatrie Biologică, vol. 34, nr. 4, 30 mai 2010, pp. 652-656, DOI : 10.1016 / j.pnpbp.2010.03.004 . Adus la 30 aprilie 2020 .
- ^ a b Ramkumar Kuruba, Bharathi Hattiangady și Ashok K. Shetty, Hippocampal Neurogenesis and Neural Stem Cells in Temporal Lobe Epilepsy , în Epilepsy & behaviour: E&B , vol. 14, Suppl 1, 2009-1, pp. 65-73, DOI : 10.1016 / j.yebeh.2008.08.020 . Adus la 30 aprilie 2020 .
- ^ Bernard S. Chang și Daniel H. Lowenstein, Epilepsie , New England Journal of Medicine , vol. 349, n. 13, 25 septembrie 2003, pp. 1257-1266, DOI : 10.1056 / NEJMra022308 . Adus la 30 aprilie 2020 .
- ^ (EN) Robert S. Sloviter, The neurobiology of temporal lobe epilepsy: too much information, not enough knowledge , în Comptes Rendus Biologies, vol. 328, nr. 2, 1 februarie 2005, pp. 143-153, DOI : 10.1016 / j.crvi.2004.10.010 . Adus la 30 aprilie 2020 .
- ^ (EN) Ramkumar Kuruba, Hattiangady Bharathi și Ashok K. Shetty, neurogeneză hipocampală și celule stem neuronale în epilepsia lobului temporal , în Epilepsia și comportamentul, vol. 14, n. 1, 1 ianuarie 2009, pp. 65-73, DOI : 10.1016 / j.yebeh.2008.08.020 . Adus la 30 aprilie 2020 .
Bibliografie
- F Aboitiz, Morales D, Montiel J, Originea evolutivă a izocortexului de mamifere: Către o abordare integrată de dezvoltare și funcțională , în Behav. Brain Sciences , vol. 26, 2003, pp. 535-552, PMID 15179935 .
- D Amaral, Lavenex P, Ch 3. Neuroanatomia hipocampală , în Andersen P, Morris R, Amaral D, Bliss T, O'Keefe J (eds), The Hippocampus Book , Oxford University Press, 2006, ISBN 978-0- 19- 510027-3 .
- C Broglio, Gómez A, Durán E, Ocaña FM, Jiménez-Moya F, Rodríguez F, Salas C, Semne distinctive ale unui plan comun pentru vertebrate din creier: zone paliale specializate pentru memoria spațială, temporală și emoțională la pești actinopterigieni , în Brain Res. Bull . , vol. 57, 2002, pp. 397-399, PMID = 16144602.
- G Buzsáki, Oscilații Theta în hipocamp ( PDF ), în Neuron , vol. 33, 2002, pp. 325-340, PMID = 11832222 (arhivat din original la 25 iunie 2008) .
- G Buzsáki, Rhythms of the Brain , Oxford University Press, 2006, ISBN 0-19-530106-4 .
- JL Cantero, Atienza M, Stickgold R, Kahana MJ, Madsen JR, Kocsis B, oscilații theta dependente de somn în hipocampul uman și neocortex , în J. Neurosci. , vol. 23, 2003, pp. 10897-10903, PMID = 14645485.
- M Colombo, Broadbent N, este hipocampul aviar un omolog funcțional al hipocampului mamifer? , în Neurosci. Biobehav. Rev. , vol. 24, 2000, pp. 465-484, PMID 10817844 .
- H Eichnbaum, Cohen NJ, Memory, Amnesia, and the Hippocampal System , MIT Press, 1993.
- AD Ekstrom, Kahana MJ, Caplan JB, Fields TA, Isham EA, Newman EL, Fried I, Rețele celulare care stau la baza navigației spațiale umane ( PDF ), în Nature , vol. 425, 2003, pp. 184-188, PMID = 12968182. Adus la 19 aprilie 2008 (arhivat din original la 11 mai 2008) .
- JA Gray, McNaughton N,Neuropsihologia anxietății: o anchetă despre funcțiile sistemului Septo-Hipocampal , Oxford University Press, 2000.
- JD Green, Arduini AA, Activitatea electrică a hipocampului în excitare , în J. Neurofiziol. , vol. 17, 1954, pp. 533-557, PMID = 13212425.
- LF Jacobs, Gaulin SJ, Sherry DF, Hoffman GE, Evoluția cogniției spațiale: tiparele de comportament spațial specifice sexului prezic dimensiunea hipocampului , în PNAS , vol. 87, 1990, pp. 6349-6352, PMID 2201026 .
- LF Jacobs, Evoluția hărții cognitive , în Brain Behav. Evol. , vol. 62, 2003, pp. 128-139, DOI : 10.1159 / 000072443 , PMID 12937351 .
- IJ Kirk, McNaughton N, Tragerea celulelor supramamilare și activitatea lentă a ritmului hipocampic , în Neuroreport , vol. 11, 1991, pp. 723-725, PMID = 1810464.
- EA Maguire, Gadian DG, Johnsrude IS, Good CD, Ashburner J, Frackowiak RS, Frith CD, Schimbare structurală legată de navigație în hipocampii șoferilor de taxi , în PNAS , vol. 97, 2000, pp. 4398-4403, DOI : 10.1073 / pnas.070039597 , PMID = 10716738.
- BL McNaughton, Battaglia FP, Jensen O, Moser EI, Moser MB, Integrarea căii și baza neuronală a „hărții cognitive” , în Nat. Pr. Neurosci. , vol. 7, 2006, pp. 663-678, PMID = 16858394.
- EI Moser, Kropf E, Moser MB, Place Cells, Grid Cells, and the Brain's Spatial Representation System , in Ann. Rev. Neurosci. , vol. 31, 2008, DOI : 10.1146/annurev.neuro.31.061307.090723 .
- R Nieuwenhuys, An Overview of the Organization of the Brain of Actinopterygian Fishes , in Am. Zool. , vol. 22, 1982, pp. 287-310, DOI : 10.1093/icb/22.2.287 .
- G O'Kane, Kensinger EA, Corkin S, Evidence for semantic learning in profound amnesia: An investigation with patient HM [ collegamento interrotto ] , in Hippocampus , vol. 14, 2004, pp. 417-425, PMID =15224979.
- J O'Keefe, Nadel L, The Hippocampus as a Cognitive Map , Oxford University Press, 1978.
- M Portavella, Vargas JP, Torres B, Salas C, The effects of telencephalic pallial lesions on spatial, temporal, and emotional learning in goldfish , in Brain Res. Bull. , vol. 57, 2002, pp. 397-399, PMID =11922997.
- F Rodríguez, Lópeza JC, Vargasa JP, Broglioa C, Gómeza Y, Salas C, Spatial memory and hippocampal pallium through vertebrate evolution: insights from reptiles and teleost fish , in Brain Res. Bull. , vol. 57, 2002, pp. 499-503, PMID 11923018 .
- RS Sainsbury, Heynen A, Montoya CP, Behavioral correlates of hippocampal type 2 theta in the rat , in Physiol. Behav. , vol. 39, 1987, pp. 513-519, PMID =3575499.
- WB Scoville, Milner B, Loss of Recent Memory After Bilateral Hippocampal Lesions , in J. Neurol. Neurosurg. Psych. , vol. 20, 1957, pp. 11-21. URL consultato il 19 aprile 2008 (archiviato dall' url originale il 4 maggio 2008) .
- SJ Shettleworth, Memory and Hippocampal Specialization in Food-Storing Birds: Challenges for Research on Comparative Cognition , in Brain Behav. Evol. , vol. 62, 2003, pp. 108-116, PMID 12937349 .
- WE Skaggs, McNaughton BL, Wilson MA, Barnes CA, Theta phase precession in hippocampal neuronal populations and the compression of temporal sequences [ collegamento interrotto ] , in Hippocampus , vol. 6, 1996, pp. 149-176, PMID 8797016 .
- LR Squire, Memory and the hippocampus: a synthesis from findings with rats, monkeys, and humans , in Psych. Rev. , vol. 99, 1992, pp. 195-231.
- LR Squire, Schacter DL, The Neuropsychology of Memory , Guilford Press, 2002.
- CH Vanderwolf, Hippocampal electrical activity and voluntary movement in the rat , in EEG & Clin. Neurophysiol. , vol. 26, 1969, pp. 407-418, PMID =4183562.
- JP Vargas, Bingman VP, Portavella M, López JC, Telencephalon and geometric space in goldfish , in Eur. J. Neurosci. , vol. 24, 2006, pp. 2870-2878, PMID =17156211.
- MJ West, Stereological studies of the hippocampus: a comparison of the hippocampal subdivisions of diverse species including hedgehogs, laboratory rodents, wild mice and men. , in Prog. Brain Res. , vol. 83, 1990, pp. 13-36, PMID =2203095.
Galleria d'immagini

Dissezione superficiale della radice del cervello. Visione laterale.

Sezione del cervello.

Ventricolo sinistro.

Corno inferiore e posteriore, visti dall'alto.

Il fornice e il corpo calloso visti dal basso.

Sezione frontale (coronale) del cervello).
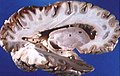
Cervello umano sezionato

Diagramma dell'ippocampo.

Aree dell'ippocampo

Forma caratteristica dell'ippocampo










Voci correlate
Altri progetti
-
 Wikiquote contiene citazioni sull' ippocampo
Wikiquote contiene citazioni sull' ippocampo -
 Wikizionario contiene il lemma di dizionario « ippocampo »
Wikizionario contiene il lemma di dizionario « ippocampo » -
 Wikimedia Commons contiene immagini o altri file sull' ippocampo
Wikimedia Commons contiene immagini o altri file sull' ippocampo
Collegamenti esterni
- ( EN ) Ippocampo , su Enciclopedia Britannica , Encyclopædia Britannica, Inc.
- Article in New Scientist , su newscientist.com .
- Scientific webpage of the project , su bmsr.usc.edu . URL consultato il 19 aprile 2008 (archiviato dall' url originale il 31 luglio 2007) .
- Diagram of network ( GIF ), su bris.ac.uk . URL consultato il 19 aprile 2008 (archiviato dall' url originale il 16 maggio 2006) .
- International Symposium “HIPPOCAMPUS AND MEMORY” , su hippo2006.psn.ru . URL consultato il 18 aprile 2019 (archiviato dall' url originale il 26 dicembre 2005) .
- Diagram of a Hippocampal Brain Slice , su stanford.edu .
- Temporal-lobe.com An interactive diagram of the rat parahippocampal-hippocampal region , su temporal-lobe.com .
{kind=link}
| Controllo di autorità | Thesaurus BNCF 51458 · LCCN ( EN ) sh85060982 · GND ( DE ) 4642127-0 · BNF ( FR ) cb11983242q (data) |
|---|