Dimetrodon
| Dimetrodon | |
|---|---|
 | |
| Gama geologică | |
| Starea de conservare | |
Fosil | |
| Clasificare științifică | |
| Domeniu | Eukaryota |
| Regatul | Animalia |
| Phylum | Chordata |
| Superclasă | Tetrapoda |
| Clasă | Synapsida |
| Ordin | † Pelicozaurie |
| Familie | † Sphenacodontidae |
| Subfamilie | † Sphenacodontinae |
| Tip | † Dimetrodon Cope , 1878 |
| Nomenclatura binominala | |
| † Dimetrodon limbatus Cope, 1878 (inițial Clepsydrops limbatus ) | |
| Sinonime | |
| |
| Specii | |
Dimetrodon (al cărui nume înseamnă „dinți de două dimensiuni”) este un gen dispărut de sinapsă sfenacodontidă care a trăit în perioada permiană , epoca Cisuralianului , în urmă cu aproximativ 295-272 milioane de ani ( Sakmarian - Kungurian ). [1] [2] [3] Este un membru al familiei sphenacodontidae . Cea mai faimoasă caracteristică a Dimetrodonului este, fără îndoială, vela mare dorsală, formată din procese spinale alungite vertical conectate între ele printr-un strat de piele. Animalul era patruped și avea un craniu înalt, curbat, cu dinți mari de diferite dimensiuni așezate de-a lungul maxilarelor. Majoritatea fosilelor au fost găsite în sud-vestul Statelor Unite , iar majoritatea provin dintr-un depozit geologic numit Red Beds, în Texas și Oklahoma . Mai recent, fosile de Dimetrodon au fost găsite și în Germania . De la descrierea sa în 1878, au fost descrise peste o duzină de specii atribuite genului.
Deși în cultura populară este adesea confundat cu un dinozaur sau contemporanul lor, Dimetrodon a dispărut cu aproximativ 40 de milioane de ani înainte de apariția dinozaurilor. Deși aspectul său și fiziologia îl fac să semene cu reptilele , Dimetrodonul este mai strâns legat de mamifere , chiar dacă nu este un strămoș direct. [2] Dimetrodonul a fost catalogat ca „proto-mamifer” ( stem-mamifer ). [2] Împreună cu mamiferele, acest grup, care conține Dimetrodon , alcătuiește clada (grupul evolutiv) numit Synapsida , în timp ce dinozaurii , reptilele și păsările fac parte din clada separată, Sauropsida . Deschiderile unice ale craniului din spatele fiecărui ochi, cunoscute sub numele de fenestre temporale , și alte caracteristici ale craniului disting Dimetrodon și mamifere de majoritatea sauropsidelor timpurii.
Dimetrodonul a fost probabil unul dintre super prădătorii ecosistemelor cisuraliene, vânând și hrănindu-se cu pești și tetrapode , inclusiv reptile și amfibieni , în timp ce speciile mai mici de Dimetrodon ar fi putut avea roluri ecologice diferite. S-au avansat numeroase ipoteze cu privire la funcția velei Dimetrodon : cea mai acreditată ipoteză de la descoperirea sa este că această structură a servit pentru termoreglarea corpului și a servit la preluarea și dispersarea căldurii mai repede. Cu toate acestea, descoperirea endotermiei în sfenacodontide [4] a infirmat această ipoteză și acum se crede în principal că pânza avea doar funcția de atracție sexuală, precum și la alte animale precum Sphenacodon și Edaphosaurus .
Descriere
Dimetrodonul era o sinapsă cvadrupedală , ale cărei specii variau ca mărime, variind în lungime de la 1,7 până la 4,6 metri (5,6 până la 15,1 picioare) și se estimează că greutatea sa era între 28 și 250 de kilograme (62 și 551 lbs) . [5] Cea mai mare specie cunoscută de Dimetrodon este D. angelensis la 4,6 metri (15 picioare), în timp ce cea mai mică specie este D. teutonis la doar 60 de centimetri (24 inci). [6] [7] Cea mai mare specie a lui Dimetrodon trebuie să fi fost cei mai mari prădători ai Permianului inferior, deși Tappenosaurusul înrudit, cunoscut doar din fragmentele scheletice găsite în roci ceva mai tinere, ar fi putut fi chiar mai mare cu o lungime de aproximativ 5,5 metri ( 18 picioare). [8] [9] Cu toate acestea, unele specii de Dimetrodon sunt cunoscute doar de la exemplare tinere, astfel încât animalele adulte ar fi putut fi mult mai mari. [10]
Craniu
Dimetrodon poseda o singură deschidere mare de ambele părți ale spatelui craniului, care îl conectează cu mamifere și îl deosebește de majoritatea sauropsidelor timpurii, care fie nu aveau, fie aveau două deschideri complet . Caracteristici precum crestele din interiorul cavității nazale și o creastă din spatele mandibulei sunt considerate a face parte dintr-o evoluție evolutivă a tetrapodelor timpurii (vertebrate cu patru picioare) către mamifere.
Craniul lui Dimetrodon este înalt, curbat și comprimat lateral . Prizele pentru ochi sunt poziționate sus și în spatele fiecărei prize pentru ochi există o singură gaură, numită fereastră temporală . O gaură suplimentară în craniu, fenestra supratemporală , poate fi văzută când este privită de sus. Partea din spate a craniului (regiunea occipitală ) este orientată la un unghi ușor în sus, caracteristică pe care o împarte cu toate celelalte sinapide timpurii. [11] Marginea superioară a craniului se înclină într-un arc convex până la vârful botului. Vârful maxilarului superior, format de premaxilla , este ridicat deasupra părții maxilarului formată de osul maxilar , fiind împărțit printr-un fel de canelură care împarte premaxilla de maxilarul propriu-zis. Această canelură se numește diastemă sau decalaj în rândul dinților. Craniul lui Dimetrodon este, de asemenea, mai robust decât al unui dinozaur , datorită absenței cavităților.
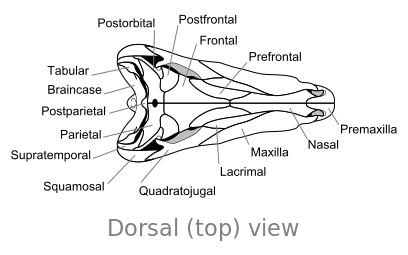
Oasele craniului lui Dimetrodon , în vedere dorsală
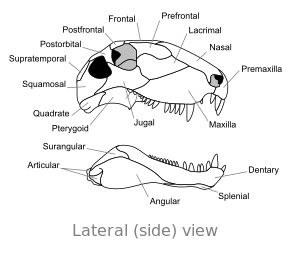
Oasele craniului Dimetron , în vedere laterală
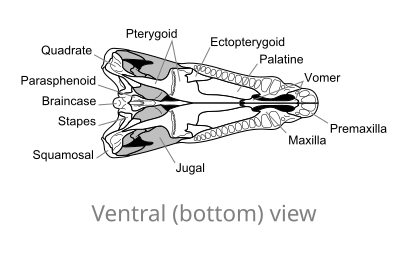
Oasele craniului lui Dimetrodon , în vedere ventrală
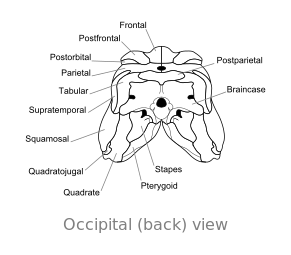
Oasele craniului lui Dimetrodon , în viziunea occipitală


Dinții

Mărimea dinților variază în funcție de lungimea maxilarelor, dând astfel numele lui Dimetrodon , ceea ce înseamnă, de fapt, „dinți de două dimensiuni” în raport cu grupurile de dinți mici și mari. [12] Una sau două perechi de dinți canini (mari canine- ca și dinți ) se extind de la maxilei. Există, de asemenea, incisivi caniniformi mari prezenți la extremitatea maxilarului superior și inferior, înrădăcinați în premaxilă și, de asemenea, în osul dentar . Dinții mai mici erau prezenți de-a lungul maxilarului și în spatele dinților canini, devenind mai mici pe măsură ce se retrăgeau în maxilar. [13]
Mulți dinți sunt mai largi în secțiunea lor și se micșorează în apropierea maxilarelor, dându-le aspectul unei lacrimi. Forma „lacrimă” a dinților este unică pentru Dimetrodon și alte sfenacodontide strâns înrudite și adesea îi ajută pe paleontologi să le distingă de alte sinapide primitive. [7] La fel ca în multe alte sinapside primitive, dinții majorității speciilor de Dimetrodon sunt zimțate la margini. [7] Indentările dentare ale Dimetrodonului erau atât de fine încât păreau mici fisuri. [14] Dinozaurul Albertosaurus a avut și indentări similare, dar la baza fiecărei indentări este un cerc gol , care ar fi servit pentru a distribui forța pe o suprafață mare și pentru a preveni stresul și a-l difuza prin dinte. Spre deosebire de Albertosaurus , totuși, dinților lui Dimetrodon le lipseau adaptările care împiedicau formarea fisurilor în canelurile lor. [14] Dinții speciei D. teutonis nu aveau zimțări , dar aveau totuși margini ascuțite. [14]
Un studiu din 2014 arată că dinții genului Dimetrodon au fost adaptați la dimensiunea prăzii sale. Specii mai mici, cum ar fi D.milleri , nu au avut indentări , deoarece au mâncat pradă mică. Cu cât prada este mai mare, cu atât dinții diferitelor specii de Dimetrodon au dezvoltat indentări și o creștere progresivă a dimensiunii. Specia D. limbatus avea indentări de smalț care ajuta la mărunțirea cărnii. Aceleași indentări pot fi găsite și la Secodontosaurus . A doua cea mai mare specie a lui Dimetrodon , D. grandis , avea indentări similare cu cele ale rechinilor și dinozaurilor teropode , făcându-și dinții și mai specializați pentru tăierea cărnii. Acest studiu nu numai că arată evoluția lui Dimetrodon de-a lungul a milioane de ani, dar demonstrează, de asemenea, că Dimetrodon se afla într-o cursă înarmării în desfășurare împotriva prăzii sale. Pe măsură ce prada a devenit mai mare, cu atât Dimetrodonul a contracarat creșterea în dimensiune și dezvoltarea unor dinți mai ascuțiți. [15]
Cavitatea nazală
Pe suprafața internă a secțiunii nazale a craniului erau mici structuri numite creste nazoturbinale, utilizate probabil pentru a susține structurile cartilajului, care au mărit suprafața epiteliului olfactiv , stratul de țesut care detectează mirosurile. Aceste creste sunt mult mai mici decât cele ale sinapsidelor mai evoluate, la sfârșitul Permianului și Triasicului, ale căror nazoturbinale erau dovezi ale endotermiei , deoarece susțineau membranele mucoase care încălzeau și umidificau aerul de intrare. Astfel, cavitatea nazală a lui Dimetrodon pare a fi o tranziție între cea a vertebratelor terestre timpurii și a mamiferelor. [16]
Articulația mandibulară și urechea
O altă caracteristică de tranziție a Dimetrodonului este o creastă din spatele mandibulei numită lamina reflexă. Lamina reflexă se găsește pe articulațiile osului, care se conectează la osul pătrat al craniului pentru a forma mandibula. Strămoșii mamiferelor de mai târziu au o articulație și un os pătrat separat de mandibulă în timp ce articulația se dezvoltă în ciocanul urechii medii . Lamina reflexă face parte dintr-un inel numit inelul timpanic care susține timpanul la toate mamiferele vii. [17]
Coadă

Coada Dimetrodonului reprezintă o mare parte din lungimea totală a corpului și include aproximativ 50 de vertebre caudale . Scheletele timpurii ale lui Dimetrodon erau incomplete, iar coada lipsea; singurul indiciu al prezenței sale a fost descoperirea a unsprezece vertebre caudale găsite lângă o parte . Deoarece aceste vertebre caudale timpurii s-au îngustat rapid pe măsură ce s-au îndepărtat de flanc, mulți paleontologi de la sfârșitul secolului al XIX-lea și începutul secolului al XX-lea au crezut că Dimetrodon are o coadă foarte scurtă. Abia după 1927, o coadă în mare parte completă a Dimetrodonului a fost găsită și descrisă în cele din urmă. [18]
Naviga

Vela lui Dimetrodon este formată dintr-o alungire a coloanelor neuronale care ies din vertebre . Fiecare coloană neurală variază în secțiune și formă de la baza sa până la vârful său în ceea ce este cunoscut sub numele de diferențiere "dimetrodon". [19] În apropierea corpului vertebrei, secțiunea transversală a coloanei vertebrale este comprimată lateral într-o formă dreptunghiulară și, mai aproape de vârf, își asumă o cifră opt ca o canelură care se întinde de-a lungul ambelor părți ale coloanei vertebrale. Se crede că cifra de opt a servit la întărirea coloanei vertebrale, prevenind flexia și fracturile. [20] O secțiune transversală a coloanei vertebrale a unui specimen de Dimetrodon giganhomogenes are formă dreptunghiulară, dar păstrează forma a opt inele în formă de lângă centrul său, indicând faptul că forma spinilor s-ar putea schimba de la individ la individ sau în funcție de vârstă. [21] Anatomia microscopică a fiecărei coloane vertebrale variază de la bază la vârf, indicând unde au ancorat mușchii spatelui și unde a fost expusă o parte a pânzei. Porțiunea inferioară sau proximală a coloanei vertebrale prezintă o suprafață aspră care ar fi servit ca punct de ancorare pentru mușchii epaxiali ai spatelui și are, de asemenea, o rețea de țesuturi conjunctive numite fibre Sharpey care indică, care a fost încorporată în corp . Mai sus pe partea distală (exterioară) a coloanei vertebrale, suprafața osoasă este netedă. Periostul , un strat de țesut care înconjoară osul, este acoperit cu mici caneluri care probabil au susținut vasele de sânge care au furnizat baldachinul. [22]
Șanțurile mari care parcurgeau întreaga lungime a coloanei vertebrale au fost considerate odată ca fiind un canal pentru vasele de sânge , dar, din moment ce osul nu conține canale vasculare, coronamentul nu trebuia să fie la fel de vascularizat pe cât se credea anterior. Unele exemplare de Dimetrodon rețin zone deformate ale coloanei vertebrale neuronale care par să se fi vindecat după fractură. Osul cortical care crește în aceste zone deformate este foarte vascularizat, sugerând că țesutul moale trebuie să fi fost prezent pe baldachin pentru a alimenta locul cu vase de sânge. [20] Osul lamelar constituie majoritatea secțiunii transversale a coloanei vertebrale neuronale și conține linii de oprire a creșterii care pot fi utilizate pentru a determina vârsta fiecărui individ la moarte. [23] La multe exemplare de D. gigashomogenes porțiunile distale ale spinilor sunt îndoite brusc, indicând faptul că pânza avea un profil neregulat când animalul era în viață. Flexibilitatea lor ar putea sugera că țesutul moale care leagă coloanele vertebrale neurale poate să nu fie prezent până la vârfurile coloanei vertebrale, ceea ce înseamnă că întinderea velei pielii nu era atât de largă pe cât se credea anterior. [19]
Piele
Nu s-au găsit niciodată dovezi fosile pe pielea lui Dimetrodon . Cu toate acestea, sa constatat impresia pielii unui animal legat de Dimetrodon, anii Estemmenosuchus , care avea o piele încrețită și uscată și echipate cu glande, similară cu cea a elefanților [24] cu structuri mici , cum ar fi solzi pe abdomen si pe partea inferioara o parte a cozii, ca și alte sinapside. [25] [26]
Descoperire și clasificare
Primele descrieri ale lui Cope

Fosilele atribuite acum lui Dimetrodon , au fost studiate de paleontologul american Edward Drinker Cope , în 1870. Cope a obținut fosilele împreună cu cele ale multor altor tetrapode permiene (vertebrate cu patru picioare), trimise lui de mai mulți colecționari care explorau un grup de roci din Texas , numit Pat Roșu. Printre acești colecționari s-au numărat naturalistul elvețian Jacob Boll, geologul din Texas WF Cummins și paleontologul amator Charles Hazelius Sternberg. [27] Majoritatea exemplarelor lui Cope au fost trimise la Muzeul American de Istorie Naturală și / sau Muzeul Walker al Universității din Chicago (cea mai mare parte a colecției de fosile ale lui Walker este acum găzduită în Field Museum ).
Sternberg i-a trimis câteva dintre probele sale paleontologului german Ferdinand Broili , la Universitatea din München , deși Broili nu a fost atât de grozav când a fost vorba de descrierea probelor. Rivalul lui Cope, Othniel Charles Marsh a găsit și câteva oase de Dimetrodon , pe care le-a trimis ulterior la Muzeul Walker. [28] Numele Dimetrodon a fost folosit pentru prima dată în descrierea lui Cope (1878), când Cope a numit primele trei specii, și anume Dimetrodon incisivus , D. rectiformis și D. gigas , în revista științifică Proceedings of the American Philosophical Society . [29]
Cu toate acestea, prima descriere a fosilelor Dimetrodon a venit cu un an mai devreme, când Cope a numit specia Clepsydrops limbatus , din Patul Roșu din Texas. [30] Mai târziu, specia a fost schimbată din C. limbatus într-o specie Dimetrodon , devenind D. limbatus .
Cu descrierea lui Natalis Clepsydrops, Cope a fost primul care a descris - o Crested sinapsă , cu toate că el a descris vela Dimetrodon e ca o aripioară și comparat cu creasta dorsală a moderne Baziliscul (gen. Basilicus). Vele nu au fost conservate în exemplarele D. incisivus și D. gigas , așa cum a fost descris de Cope (1878), dar spinii lungi alungi au fost prezenți în fosilele D. rectiformis . [13] Cope a comentat, de asemenea, scopul pânzei în 1886, afirmând că „utilitatea [pânzei] este greu de imaginat. Cu excepția cazului în care animalul avea obiceiuri acvatice și nu înota pe spate, creastă sau înotătoare. au un scop motor ... membrele nu erau suficient de lungi, iar ghearele nu erau suficient de ascuțite pentru a demonstra obiceiurile copacilor, ca în genul existent Basilicus , unde există de fapt o creastă similară. " [13]
Primele descrieri ale secolului XX
În primele decenii ale secolului al XX-lea, paleontologul american Ermine Cowles Case, autor al multor studii asupra Dimetrodonului, a descris mai multe specii noi din gen. Case a primit mai multe finanțări de la Carnegie Institution pentru studiul său asupra numeroaselor exemplare de Dimetrodon din colecțiile Muzeului American de Istorie Naturală și din multe alte muzee. [28] Multe dintre aceste fosile fuseseră colectate de Cope, dar nu fuseseră încă descrise cu exactitate, deoarece se știa că Cope erigea specii noi chiar și pe baza câtorva fragmente osoase.
Începând cu sfârșitul anilor 1920, paleontologul Alfred Romer a re-studiat multe exemplare de Dimetrodon și a numit mai multe specii noi. În 1940, Romer a fost coautor al studiului amplu alături de Llewellyn Ivor Price, supranumit „Revista Pelycosauria ”, în care speciile Dimetrodon au fost reevaluate de Case. Majoritatea denumirilor de specii considerate valabile de Romer și Price sunt încă în uz astăzi. [22]
Specimene noi
În următoarele decenii: numeroase specii noi de Dimetrodon au fost adăugate la monografia lui Romer și Price, provenind din localități din afara Texasului și Oklahoma. Primul a fost găsit în Four Corners , Utah , în 1966 [31] în timp ce altul a fost găsit în Arizona, în 1969. [32] În 1975, Olson a raportat material nou găsit în Ohio, la Dimetrodon . [33] O nouă specie de Dimetrodon , numită D. occidentalis (al cărei nume înseamnă „dimetrodon occidental”) a fost numită în 1977, din New Mexico . [34] Exemplarele găsite în Utah și Arizona aparțin probabil speciei D. occidentalis . [35]
Inainte de aceste descoperiri, a existat ca în teoria Permian inferior, o mare interioară care separă ceea ce astăzi sunt Texas și Oklahoma din ținuturile vestice, izolarea Dimetrodon într - o regiune mică a Americii de Nord, în timp ce sphenacodontide mai mici sphenacodon , a dominat zona de vest. Deși prezența acestei mări interioare este foarte probabilă, descoperirea fosilelor în afara Texasului și Oklahoma arată că întinderea sa era limitată și că nu era o barieră eficientă pentru distribuția Dimetrodonului . [34] [36]
În 2001, o nouă specie de Dimetrodon , numită D. teutonis, a fost descrisă dintr-o locație permiană timpurie din pădurea Turingia , Germania , extinzând pentru prima dată aria geografică a Dimetrodon dincolo de America de Nord. [5]
Specii
| Specii | Autor / i | Închiriere | Stat | Sinonime | Imagine |
|---|---|---|---|---|---|
| Dimetrodon angelensis | Olson, 1962 | Texas | Valabil |  | |
| Dimetrodon boreal | Leidy, 1854 | Insula Prințul Eduard | Valabil | Cunoscut inițial sub numele de Bathygnathus borealis |  |
| Dimetrodon booneorum | Romer, 1937 | Texas | Valabil | ||
| Dimetrodon dollovianus | Case, 1907 | Texas | Valabil | Embolophorus dollovianus Cope, 1888 | |
| Dimetrodon gigashomogenes | Case, 1907 | Texas | Valabil |  | |
| Dimetrodon grandis | Romer & Price, 1940 | Oklahoma Texas | Valabil | Clepsydrops gigas Cope, 1878 Dimetrodon gigas Cope, 1878 Theropleura grandis Case, 1907 Cazul Bathyglyptus theodori , 1911 Dimetrodon maximus Romer 1936 |  |
| Dimetrodon kempae | Romer, 1937 | Texas | Nomen dubium posibil | ||
| Dimetrodon limbatus | Romer & Price, 1940 | Oklahoma Texas | Valabil | Clepsydrops limbatus Cope, 1877 Dimetrodon incisivus Cope, 1878 Dimetrodon rectiformis Cope, 1878 Dimetrodon semiradicatus Cope, 1881 | |
| Dimetrodon loomisi | Romer, 1937 | Texas Oklahoma | Valabil |  | |
| Dimetrodon macrospondylus | Case, 1907 | Texas | Valabil | Clepsydrops macrospondylus Cope, 1884 Cazul Dimetrodon platycentrus , 1907 | |
| Dimetrodon milleri | Romer, 1937 | Texas | Valabil |  | |
| Dimetrodon natalis | Romer, 1936 | Texas | Valabil | Clepsydrops natalis Cope, 1878 |  |
| Dimetrodon occidentalis | Berman, 1977 | Arizona New Mexico Utah | Valabil | ||
| Dimetrodon teutonis | Berman și colab. , 2001 | Germania | Valabil |
Filogenetica
Dimetrodonul este unul dintre primii membri ai unui grup numit sinapsi , care include mamifere și multe dintre rudele lor dispărute, deși Dimetrodonul nu este strămoșul tuturor mamiferelor (care au apărut milioane de ani mai târziu [37] ). Dimetrodonul este adesea confundat cu un dinozaur în cultura populară, deși a dispărut cu aproximativ 40 de milioane de ani înainte ca primii dinozauri să apară în Triasic . La fel ca toate sinapsidele, Dimetrodon este mai strâns legat de mamifere decât de dinozauri sau orice altă reptilă vie. Din 1900, majoritatea paleontologilor s-au referit la Dimetrodon drept reptilă, în conformitate cu taxonomia lineană, care a clasificat Reptilia ca o clasă și Dimetrodon ca un gen în această clasă. Mamiferele au fost alocate unei clase separate, iar Dimetrodon a fost descris ca o „reptilă asemănătoare unui mamifer”. Paleontologii au teoretizat că mamiferele au evoluat din acest grup, într-o tranziție reptilian-mamifer.
Taxonomia sinapsidelor

Sub sistematica filogenetică , descendenții ultimului strămoș comun al lui Dimetrodon și toate reptilele vii includ toate mamiferele, deoarece Dimetrodon este mai strâns legat de mamifere decât orice altă reptilă vie. Astfel, dacă se dorește evitarea cladei care conține atât mamifere, cât și reptile vii, Dimetrodon nu trebuie inclus în acea cladă, nici în nicio altă cladă „reptilă asemănătoare unui mamifer”. Descendenții ultimului strămoș comun al mamiferelor și reptilelor (care au apărut acum aproximativ 310 milioane de ani, în Carboniferul superior ) sunt, prin urmare, împărțiți în două clade: Synapsida , care include Dimetrodon și mamifere, și Sauropsida , care include reptilele vii și toate dispărute. reptile mai strâns legate între ele decât de mamifere. [2]
În cadrul clade Synapsida, Dimetrodon face parte din Sphenacodontia clade, care a fost propusă ca un grup de synapsids la începutul anilor 1940 de paleontologii Alfred Romer și Llewellyn Ivor Price, împreună cu Ophiacodontia și grupul Edaphosauria . [38] Toate cele trei grupuri sunt cunoscute de la Carboniferul superior până la Permianul inferior . Romer și Price le disting în principal prin trăsături postcraniene, cum ar fi formele membrelor și vertebrelor. Ophiacodontia este considerată cel mai primitiv grup, deoarece membrii săi sunt cei mai înrudiți cu reptilele, în timp ce Sphenacodontia a fost cel mai avansat grup, deoarece membrii săi au fost cei mai înrudiți cu Therapsida , care a inclus cele mai apropiate rude ale mamiferelor. Romer și Preț plasat un alt grup printre synapsids primitive, numite varanopids in cadrul Sphenacodontia, considerându - le mai primitivi decât alte sphenacodonts, cum ar fi Dimetrodon. [39] În trecut , se credea că varanopids au fost strâns legate de Dimetrodon- ca sphenacodonts, ca ambele grupuri au fost carnivore, deși varanopids sunt mult mai mici și nu mai au vele.
Viziunea modernă a relațiilor dintre sinapside a fost propusă de paleontologul Robert R. Reisz , în 1986, al cărui studiu include trăsături găsite mai ales în craniu, mai degrabă decât în scheletul postcranian. [40] Dimetrodon este încă considerat un sfenacodont sub această filogenie, dar varanodontidele sunt acum considerate sinapside mai bazale, neconforme cu clada Sphenacodontia. În cadrul Sphenacodontia se află grupul Sphenacodontoidea, care la rândul său conține Sphenacodontidae și Therapsida. Sphenacodontidae este grupul care conține Dimetrodon și alte câteva sinapside cu navigație, cum ar fi Sphenacodon și Secodontosaurus , în timp ce Therapsida include mamifere și rudele lor din perioadele permian și triasic.
Următoarea este cladograma cladei Synapsida , care urmează filogenia sinapsidei modificată din analiza lui Benson (2012): [39]
| Amniota |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Următoarea cladogramă arată relațiile unor specii de Dimetrodon , de Brink și colab. , (2015): [41]
| Sphenacodontidae |
| ||||||||||||||||||||||||||||||||||||
Paleobiologie
Funcția Sail
Paleontologii au propus mai multe ipoteze cu privire la funcția efectivă a pânzei în viață. Una dintre primele ipoteze propuse a fost utilizarea lor în camuflajul animalului, care s-ar putea camufla astfel între stuf , în timp ce Dimetrodonul își aștepta prada. O altă ipoteză destul de imaginativă este că a funcționat ca o adevărată pânză , pentru mișcarea animalului în apă, captând vântul. [42] O altă ipoteză este că spinii neuronali lungi aveau funcția de stabilizare a trunchiului prin limitarea mișcării verticale, ceea ce ar fi permis o mișcare laterală mai eficientă. [20]
Termoreglare
În 1940, Alfred Romer și Llewellyn Ivor Price au propus ca pânza să aibă o funcție de termoreglare , permițând Dimetrodonilor să-și încălzească corpul mai eficient, expunându-i la căldura soarelui. În anii următori, au fost create numeroase modele pentru a evalua eficacitatea termoreglării Dimetrodon . De exemplu, într-un articol din 1973 din revista Nature , paleontologii CD Bramwell și PB Fellgett au estimat că un individ de 200 kg (440 lb) ar trebui să rămână la soare aproximativ o oră și jumătate pentru a-și crește temperatura corpului . 26 până la 32 ° C (79 până la 90 ° F). [43] În 1986, Steven C. Haack a concluzionat că încălzirea animalului trebuie să fie mult mai lentă decât se credea anterior și că întregul proces ar dura probabil cel puțin patru ore. Folosind un model bazat pe o varietate de factori de mediu și a emis ipoteza unor aspecte fiziologice ale Dimetrodonului , Haack a descoperit că planorul îi va permite Dimetrodonului să se încălzească mai repede dimineața și să atingă o temperatură corporală puțin mai mare în timpul zilei, dar planorul a fost ineficient. la eliberarea excesului de căldură și nu a permis Dimetrodonului să mențină o temperatură corporală mai mare în timpul nopții. [44] Nel 1999, un gruppo di ingegneri meccanici hanno creato un modello al computer per analizzare le capacità della vela nella regolazione della temperatura corporea durante le diverse stagioni, e hanno concluso che la vela era uno strumento benefico sia per l'assorbimento sia per il rilascio di calore, in ogni momento durante l'anno. [45]

La maggior parte di questi studi danno due ruoli di termoregolazione per la vela di Dimetrodon : uno come un mezzo veloce di riscaldamento al mattino, e un altro come un mezzo di raffreddamento quando la temperatura corporea diventava troppo alta. Originariamente, il Dimetrodon , e tutti gli altri vertebrati terrestri del Permiano, venivano indicati come animali a sangue freddo o pecilotermi , dipendendo dal calore del sole per mantenere costante la temperatura corporea elevata, come i rettili attuali. A causa delle sue grandi dimensioni, il Dimetrodon aveva un'elevata inerzia termica , il che significa che i cambiamenti nella temperatura corporea si verificavano più lentamente rispetto ad animali più piccoli. All'innalzamento della temperatura al mattino, le prede del Dimetrodon dai corpi più piccoli, si scaldavano molto più rapidamente di quanto potesse fare invece il più grande Dimetrodon . Molti paleontologi tra cui Haack hanno proposto che la vela del Dimetrodon potrebbe aver permesso di scaldarsi più rapidamente al mattino, al fine di tenere il passo con le sue prede. [44] La grande superficie della vela implicava anche che il calore si dissipava più rapidamente nell'ambiente circostante, utile se l'animale aveva bisogno di rilasciare il calore in eccesso prodotto dal metabolismo o assorbito dal sole. Il Dimetrodon potrebbe anche aver angolato la vela in base alla posizione del sole per rinfrescarsi o per limitare il flusso di sangue alla vela e mantenere il calore durante la notte. [42]
Nel 1986, J. Scott Turner e Richard C. Tracy hanno proposto che l'evoluzione della vela in Dimetrodon era legata all'evoluzione del sangue caldo negli antenati dei mammiferi . Turner e Tracy hanno pensato che la vela del Dimetrodon avrebbe permesso all'animale di essere omeotermo , mantenendo un costante, seppur bassa, la temperatura corporea. Anche i mammiferi sono omeotermi, anche se differiscono da Dimetrodon nell'essere endotermici, controllare la loro temperatura corporea internamente attraverso il metabolismo accresciuto. Turner e Tracy notarono che i primi terapsidi , un gruppo più avanzato di sinapsidi strettamente legato ai mammiferi, hanno lunghi arti che possono rilasciare calore in un modo simile a quello della vela di Dimetrodon . L'omeotermia sviluppatosi in animali come Dimetrodon può essere riportato ai terapsidi attraverso una modifica della forma del corpo, che si sarebbe poi sviluppato nel sangue caldo dei mammiferi. [46]

Recenti studi sulla vela di Dimetrodon e di altri sphenacodontidi , supportano la tesi di Haack (1986) che affermava che la vela era scarsamente adatta a rilasciare calore e mantenere una temperatura corporea stabile. La presenza della vela nelle specie più piccole di Dimetrodon , come D. milleri e D. teutonis non si adattano all'idea che lo scopo della vela era di termoregolazione perché le vele più piccole sono insufficienti per trasferire il calore e perché i piccoli corpi possono assorbire e rilasciare calore facilmente da soli. Inoltre, i parenti più stretti di Dimetrodon , come lo Sphenacodon hanno creste molto basse che sarebbero state inutili come dispositivi per la termoregolazione. [22] Si pensa che la grande vela del Dimetrodon si sia evoluta proprio da creste piccole e basse come quelle dello Sphenacodon , il che significa che su gran parte della storia evolutiva della vela, la termoregolazione non sarebbe servita a molto. [47]
Gli esemplari più grandi di Dimetrodon hanno vele più grandi rispetto alle loro dimensioni, un esempio di allometria positiva. L'allometria positiva può beneficiare della termoregolazione perché significa che, come gli individui diventavano più grandi, più la superficie aumenta. Gli animali più grandi generano una grande quantità di calore attraverso il metabolismo e la quantità di calore che deve essere dissipato dalla superficie del corpo è significativamente maggiore di quello che doveva essere dissipata da animali più piccoli. La dissipazione del calore poteva essere più efficace da un singolo rapporto tra massa e superficie. Tuttavia, uno studio del 2010 di allometria in Dimetrodon ha provato un diverso rapporto tra la vela e la massa corporea: l'esponente di scala reale della vela era molto più grande dell'esponente previsto in un animale atto alla dissipazione del calore. I ricercatori hanno concluso che la vela del Dimetrodon cresceva ad un ritmo molto più veloce di quanto era necessario per la termoregolazione, e ha suggerito che la selezione sessuale è stata la ragione principale per la sua evoluzione. [47]
Selezione sessuale
L'esponente allometrica per l'altezza della vela è simile in grandezza alla scala di lunghezza interspecifica nelle corna all'altezza delle spalle nei cervidi . Inoltre, come Bakker (1970) ha osservata nel contesto di Dimetrodon , molte specie di lucertola sollevano una cresta di pelle dorsale durante il display di minaccia e corteggiamento, e positivamente allometriche, soprattutto nelle specie che presentano un forte dimorfismo sessuale (Echelle et al. , 1978; Christian et al. , 1995). Anche in Dimetrodon vi è un forte dimorfismo sessuale, soprattutto nella robustezza dello scheletro e l'altezza relativa delle spine di D. limbatus (Romer & Price, 1940). [47]
Dimorfismo sessuale
Anche il Dimetrodon potrebbe aver avuto un forte dimorfismo sessuale , il che significa che i maschi e le femmine hanno dimensioni leggermente diverse. Alcuni esemplari di Dimetrodon sono stati ipotizzati come maschi, perché hanno ossa più spesse, vele più grandi, teschi più lunghi e mascellari più profondi. Sulla base di queste differenze, gli scheletri montati al Museo Americano di Storia Naturale (AMNH 4636) e al Field Museum sarebbero in prevalenza maschi, mentre gli scheletri del Denver Museum of Natural History (MCZ 1347) e dell'University of Michigan Museum of Natural History sarebbero femmine. [38]

Scheletro di D. incisivus all'University of Michigan Museum of Natural History, che potrebbe rappresentare una femmina .


Paleoecologia

I fossili di Dimetrodon sono stati ritrovati negli Stati Uniti ( Texas , Oklahoma , Nuovo Messico , Arizona , Utah e Ohio ) e in Germania , aree che all'epoca facevano parte del supercontinente Euramerica , durante il Permiano inferiore. Negli Stati Uniti, quasi tutto il materiale attribuito a Dimetrodon viene da tre gruppi geologici nel centro-nord del Texas e dell'Oklahoma centro-meridionale: il Gruppo Clear Fork, il Gruppo di Wichita e il Gruppo River Pearce. [48] La maggior parte dei reperti fossili fanno parte degli ecosistemi di pianura, che, durante il Permiano, sarebbero state vaste zone umide . In particolare, i Red Bed del Texas, una zona di grande diversità di fossili di tetrapodi. Oltre al Dimetrodon , i tetrapodi più comuni nel Red Bed e in tutti i depositi del Permiano presto nel sud-ovest degli Stati Uniti, sono gli anfibi Archeria , Diplocaulus , Eryops e Trimerorhachis , il reptiliomorpha Seymouria , il rettile Captorhinus ei sinapsidi Ophiacodon ed Edaphosaurus . Questi tetrapodi costituiscono un gruppo di animali che il paleontologo Everett C. Olson ha chiamato la "chronofauna del Permo-Carbonifero", una fauna che ha dominato l'ecosistema dell'Euramerica per diversi milioni di anni. [49] Sulla base della geologia dei depositi come i Red Beds, questa fauna abitava in una pianura rigogliosa e piena di vegetazione in un ecosistema deltizio . [50]
Olson ha fatto molte congetture sulla paleoecologia dei Red Beds del Texas e il ruolo di Dimetrodon all'interno del suo ecosistema. Olson ha proposto diversi tipi di ecosistema in cui i primi tetrapodi avrebbero potuto vivere. Il Dimetrodon apparteneva all'ecosistema più primitivo, che si è sviluppato da una catena alimentare acquatica. In esso, le piante acquatiche sono state i produttori primari e fornivano l'alimentazione principale per pesci e invertebrati acquatici. La maggior parte dei vertebrati terrestri si nutrivano principalmente di prede acquatiche. Il Dimetrodon era probabilmente il predatore alpha dell'ecosistema, nutrendosi di una grande varietà di organismi, come il grande squalo Xenacanthus , gli anfibi acquatici Trimerorhachis e Diplocaulus , ei tetrapodi terrestri Seymouria e Trematops . All'epoca erano presenti anche numerose specie di insetti che con buona probabilità entravano anch'essi, in una certa misura, nella catena alimentare di Dimetrodon , alimentando piccoli rettili come Captorhinus . L'assemblaggio dei Red Beds includono anche alcune dei primi grandi erbivori terrestri come l' Edaphosaurus e il Diadectes , che si nutrivano principalmente di prede terrestri, anziché acquatiche. Secondo Olson, il miglior analogico moderno per l'ecosistema del Dimetrodon , sono le Everglades . [50] Lo stile di vita esatto di Dimetrodon (anfibio o terrestre) è stato a lungo dibattuto, ma la microanatomia delle ossa supporta uno stile di vita terrestre, [51] il che implica che avrebbe cacciato principalmente sulla terraferma, sulle rive o in acque molto basse. Esistono anche prove di predazione di Dimetrodon ai danni di Diplocaulus , durante i periodi di siccità, grazie al ritrovamento di tre giovani Diplocaulus parzialmente mangiati in una tana e al ritrovamento di otto segni di denti recanti da un Dimetrodon , che li estrasse dal terreno e li mangiò. [52]
L'unica specie di Dimetrodon ritrovata al di fuori del sud-ovest degli Stati Uniti è il D. teutonis , dalla Germania . I suoi resti sono stati ritrovati nella Formazione Tambach in un sito chiamato Località Bromacker. L'assemblaggio dei tetrapodi del Bromacker, nel Permiano inferiore è alquanto insolito, in quanto ci sono pochi grandi sinapsidi che servono il ruolo di grandi predatori. Le dimensioni di D. teutonis è stimata a soli 1,7 metri (5.6 ft) di lunghezza, troppo piccole per predare grandi diadectidi erbivori , abbondanti nell'assemblaggio del Bromacker. È più probabile che questa specie si nutrisse di piccoli vertebrati e insetti. Sono stati ritrovati solo tre fossili che potrebbero essere attribuiti a grandi predatori, e si ritiene che siano per lo più grandi o piccoli sphenacodonti varanopidi , entrambi i quali potevano potenzialmente predare D. teutonis . In contrasto con le pianura deltizie dei Red Beds del Texas, i depositi del Bromacker si pensa fossero rappresentati da un ambiente montano con nessuna specie acquatica. È possibile che le grandi specie di Dimetrodon non erano parte del concatenamento del Bromacker, perché erano dipendenti da grandi prede acquatiche. [5]
Nella cultura di massa
- Nonostante non sia un dinosauro, come animale preistorico il dimetrodonte è sorprendentemente ben conosciuto presso il grande pubblico. Basti pensare che è presente in un famoso film del 1959 con James Mason , Viaggio al centro della Terra , tratto da un romanzo di Verne , e poi è anche presente nel documentario della BBC L'impero dei mostri . Spesso, poi, modellini di dimetrodonte vengono inclusi nelle confezioni di giocattoli per bambini. Nonostante la sua fama, il dimetrodonte è spesso confuso con il dinosauro Spinosaurus , a causa del fatto che entrambi questi animali presentavano una vela dorsale.
- Nel Parco della Preistoria di Rivolta d'Adda sono presenti 3 ricostruzioni di dimetrodonti. Altre ricostruzioni del dimetrodonte sono presenti al Parco Preistorico di Peccioli ( PI ), al Mondo della Preistoria di Simbario ( VV ) e al Parco dei Dinosauri di Castellana Grotte ( BA ).
- Era apparso anche nel film di Walt Disney Fantasia nel capitolo La sagra della primavera e compare molto brevemente anche nel primo film della serie Alla ricerca della Valle Incantata .
- Ranu, uno dei protagonisti del cartone animato I Saurini ei viaggi del meteorite nero è un dimetrodonte.
- Uno degli zord dei Power Rangers della serie Power Rangers Dino Thunder è un dimetrodonte celeste la cui "vela" è in realtà la lama di una sega circolare.
- Un giocattolo con le fattezze del dimetrodonte appare brevemente nel film d'animazione Toy Story 3 - La grande fuga prodotto dalla Pixar Animation Studios e nel corto Toy Story: Tutto un altro mondo .
Note
- ^ Dimetrodon , su Paleobiology Database . URL consultato il 23 agosto 2012 .
- ^ a b c d KD Angielczyk, Dimetrodon is Not a Dinosaur: Using Tree Thinking to Understand the Ancient Relatives of Mammals and their Evolution , in Evolution: Education and Outreach , vol. 2, n. 2, 2009, pp. 257-271, DOI : 10.1007/s12052-009-0117-4 .
- ^ Huttenlocker, AK, and E. Rega. 2012. The Paleobiology and Bone Microstructure of Pelycosauriangrade Synapsids. Pp. 90–119 in A. Chinsamy (ed.) Forerunners of Mammals: Radiation, Histology, Biology. Indiana University Press.
- ^ Ancestry of mammalian 'warm-bloodedness' revealed , su www.sciencedaily.com , Society of Vertebrate Paleontology, 29 ottobre 2015. URL consultato il 29 ottobre 2015 .
- ^ a b c DS Berman, Reisz, RR, Martens, T. e Henrici, AC, A new species of Dimetrodon (Synapsida: Sphenacodontidae) from the Lower Permian of Germany records first occurrence of genus outside of North America ( PDF ), in Canadian Journal of Earth Sciences , vol. 38, n. 5, 2001, pp. 803-812, DOI : 10.1139/cjes-38-5-803 .
- ^ The Kungurian Age , su Palaeos . URL consultato il 29 settembre 2013 (archiviato dall' url originale il 12 novembre 2013) .
- ^ a b c J. Fröbisch, Schoch, RR, Müller, J., Schindler, T. e Schweiss, D., A new basal sphenacodontid synapsid from the Late Carboniferous of the Saar-Nahe Basin, Germany ( PDF ), in Acta Palaeontologica Polonica , vol. 56, n. 1, 2011, pp. 113-120, DOI : 10.4202/app.2010.0039 .
- ^ EC Olson e Beerbower, JR, The San Angelo Formation, Permian of Texas, and its vertebrates , in The Journal of Geology , vol. 61, n. 5, 1953, pp. 389-423, Bibcode : 1953JG.....61..389O , DOI : 10.1086/626109 .
- ^ EC Olson, Parallelism in the evolution of the Permian reptilian faunas of the Old and New Worlds , in Fieldiana , vol. 37, n. 13, 1955, pp. 385-401.
- ^ CW Sternberg, The skeleton of an immature pelycosaur, Dimetrodon cf. grandis , from the Permian of Texas , in Journal of Paleontology , vol. 16, n. 4, 1942, pp. 485-486, JSTOR 1298848 .
- ^ M. Laurin e Reisz, RR, Synapsida: mammals and their extinct relatives , su Tree of Life Web Project , 2012. URL consultato il 24 agosto 2012 .
- ^ Exhibit Specimens: Dimetrodon , su American Museum of Natural History . URL consultato il 2 luglio 2012 (archiviato dall' url originale il 4 luglio 2012) .
- ^ a b c G. Baur e Case, EC, The history of the Pelycosauria, with a description of the genus Dimetrodon , Cope , in Transactions of the American Philosophical Society , vol. 20, n. 1, 1899, pp. 5-62, JSTOR 1005488 .
- ^ a b c Abler, WL 2001. A kerf-and-drill model of tyrannosaur tooth serrations. p. 84-89. In: Mesozoic Vertebrate Life . Ed.s Tanke, DH, Carpenter, K., Skrepnick, MW Indiana University Press.
- ^ http://phenomena.nationalgeographic.com/2014/02/07/sail-backed-dimetrodon-had-a-nasty-bite/
- ^ TS Kemp, The origin and early radiation of the therapsid mammal-like reptiles: a palaeobiological hypothesis , in Journal of Evolutionary Biology , vol. 19, n. 4, 2006, pp. 1231-1247, DOI : 10.1111/j.1420-9101.2005.01076.x , PMID 16780524 .
- ^ M. Laurin e Reisz, RR, Autapomorphies of the main clades of synapsids , su Tree of Life Web Project , 1997. URL consultato il 24 agosto 2012 .
- ^ AS Romer, Notes on the Permo-Carboniferous reptile Dimetrodon , in The Journal of Geology , vol. 35, n. 8, 1927, pp. 673-689, Bibcode : 1927JG.....35..673R , DOI : 10.1086/623462 , JSTOR 30060393 .
- ^ a b EA Rega, K. Noriega, SS Sumida, A. Huttenlocker, A. Lee e B. Kennedy, Healed Fractures in the Neural Spines of an Associated Skeleton of Dimetrodon: Implications for Dorsal Sail Morphology and Function , in Fieldiana Life and Earth Sciences , vol. 5, 2012, pp. 104-111, DOI : 10.3158/2158-5520-5.1.104 .
- ^ a b c E. Rega, Sumida, S., Noriega, K., Pell, C. e Lee, A., Evidence-based paleopathology I: Ontogenetic and functional implications of dorsal sails in Dimetrodon , in Journal of Vertebrate Paleontology , vol. 25, S3, 2005, pp. 103A, DOI : 10.1080/02724634.2005.10009942 .
- ^ S. Sumida, Rega, E. e Noriega, K., Evidence-based paleopathology II: Impact on phylogenetic analysis of the genus Dimetrodon , in Journal of Vertebrate Paleontology , vol. 25, S3, 2005, pp. 120A, DOI : 10.1080/02724634.2005.10009942 .
- ^ a b c AK Huttenlocker, Rega, E. e Sumida, SS, Comparative anatomy and osteohistology of hyperelongate neural spines in the sphenacodontids Sphenacodon and Dimetrodon (Amniota: Synapsida) , in Journal of Morphology , vol. 271, n. 12, 2010, pp. 1407-1421, DOI : 10.1002/jmor.10876 , PMID 20886514 .
- ^ EA Rega, Noriega, K., Sumida, S. e Lee, A., Histological analysis of traumatic injury to multiple neural spines of an associated skeleton of Dimetrodon : Implications for healing response, dorsal sail morphology and age-at-death in a Lower Permian synapsid , in Integrated Comparative Biology , vol. 44, 2004, p. 628.
- ^ Petr Konstantinovich Tchudinov, New Facts about the Fauna of the Upper Permian of the USSR , in Journal of Geology , vol. 73, 1965, pp. 117-30.
- ^ Botha-Brink, J.; Modesto, SP (2007). "A mixed-age classed 'pelycosaur' aggregation from South Africa: earliest evidence of parental care in amniotes?". Proceedings of the Royal Society B 274 (1627): 2829–2834. doi:10.1098/rspb.2007.0803. PMC 2288685. PMID 17848370 .
- ^ Niedźwiedzki, G.; Bojanowski, M. (2012). "A Supposed Eupelycosaur Body Impression from the Early Permian of the Intra-Sudetic Basin, Poland". Ichnos 19 (3): 150–155. doi:10.1080/10420940.2012.702549
- ^ WD Matthew, Review of Case's 'Revision of the Pelycosauria of North America' , in Science , vol. 27, n. 699, 1908, pp. 816-818, DOI : 10.1126/science.27.699.816 .
- ^ a b EC Case, Revision of the Pelycosauria of North America , Washington, DC, Carnegie Institution of Washington, 1907, pp. 1-176.
- ^ ED Cope, Descriptions of extinct Batrachia and Reptilia from the Permian formation of Texas , in Proceedings of the American Philosophical Society , vol. 17, n. 101, 1878, pp. 505 –530, JSTOR 982652 .
- ^ ED Cope, Descriptions of extinct vertebrata from the Permian and Triassic Formations of the United States , in Proceedings of the American Philosophical Society , vol. 17, n. 100, 1877, pp. 182 –193, JSTOR 982295 .
- ^ PP Vaughn, Comparison of the Early Permian vertebrate faunas of the Four Corners region and north-central Texas ( PDF ), Contributions in Science, vol. 105, Los Angeles County Museum of Natural History, 1966, pp. 1-13. URL consultato il 22 marzo 2017 (archiviato dall' url originale il 10 gennaio 2017) .
- ^ PP Vaughn, Early Permian vertebrates from southern New Mexico and their paleozoogeographic significance ( PDF ), Contributions in Science, vol. 166, Los Angeles County Museum of Natural History, 1969, pp. 1-22. URL consultato il 22 marzo 2017 (archiviato dall' url originale il 10 gennaio 2017) .
- ^ EC Olson, Permo-Carboniferous paleoecology and morphotypic series , in American Zoologist , vol. 15, n. 2, 1975, pp. 371-389, DOI : 10.1093/icb/15.2.371 .
- ^ a b DS Berman, A new species of Dimetrodon (Reptilia, Pelycosauria) from a non-deltaic facies in the Lower Permian of north-central New Mexico , in Journal of Paleontology , vol. 51, n. 1, 1977, pp. 108-115, JSTOR 1303466 .
- ^ K. Madalena, Sumida, S., Zeigler, K. e Rega, E., A new record of the Early Permian pelycosaurian-grade synapsid Dimetrodon (Eupelycosauria: Sphenacodontidae) from the Lower Cutler Group (Early Permian) of Jemez Pueblo, north-central New Mexico , in Journal of Vertebrate Paleontology , vol. 27, 3, Suppl., 2007, pp. 110A, DOI : 10.1080/02724634.2007.10010458 .
- ^ PP Vaughn, Vertebrates from the Cutler Group of Monument Valley and vicinity ( PDF ), New Mexico Geological Society Guidebook, vol. 24, New Mexico Geological Society, 1973, pp. 99-105.
- ^ The mammals appear
- ^ a b AS Romer e Price, LI, Review of the Pelycosauria , in Geological Society of America Special Paper , Geological Society of America Special Papers, vol. 28, 1940, pp. 1-538, DOI : 10.1130/spe28-p1 .
- ^ a b RJ Benson, Interrelationships of basal synapsids: cranial and postcranial morphological partitions suggest different topologies , in Journal of Systematic Paleontology , vol. 10, n. 2, 2012, pp. 601-624, DOI : 10.1080/14772019.2011.631042 .
- ^ RR Reisz, Pelycosauria , in Sues, H.-D. (ed.) (a cura di), Handbuch der Paläoherpetologie , 17A, Gustav Fischer Verlag, 1986, pp. 1-102, ISBN 978-3-89937-032-4 , OL 12985656M .
- ^ Kirstin S. Brink, Hillary C. Maddin, David C. Evans, Robert R. Reisz e Hans-Dieter Sues, Re-evaluation of the historic Canadian fossil Bathygnathus borealisfrom the Early Permian of Prince Edward Island , in Canadian Journal of Earth Sciences , vol. 52, n. 12, 2015, pp. 1109-1120, DOI : 10.1139/cjes-2015-0100 .
- ^ a b GA Florides, Kalogirou, SA, Tassou, SA e Wrobel, L., Natural environment and thermal behaviour of Dimetrodon limbatus , in Journal of Thermal Biology , vol. 26, n. 1, 2001, pp. 15-20, DOI : 10.1016/S0306-4565(00)00019-X , PMID 11070340 .
- ^ CD Bramwell e Fellgett, PB, Thermal regulation in sail lizards ( PDF ), in Nature , vol. 242, n. 5394, 1973, pp. 203-205, DOI : 10.1038/242203a0 .
- ^ a b SC Haack, A thermal model of the sailback pelycosaur , in Paleobiology , vol. 12, n. 4, 1986, pp. 450-458.
- ^ GA Florides, Wrobel, LC, Kalogirou, SA e Tassou, SA, A thermal model for reptiles and pelycosaurs , in Journal of Thermal Biology , vol. 24, n. 1, 1999, pp. 1-13, DOI : 10.1016/S0306-4565(98)00032-1 .
- ^ JS Turner e Tracy, CR, Body size, homeothermy and the control of heat exchange in mammal-like reptiles ( PDF ), in Hotton, N. III.; MacLean, PD; Roth, JJ; and Roth, EC (eds.) (a cura di), The Ecology and Biology of Mammal-Like Reptiles , Washington, DC, Smithsonian Institution Press, 1986, pp. 185-194.
- ^ a b c JL Tomkins, LeBas, NR, Witton, MP, Martill, DM e Humphries, S., Positive allometry and the prehistory of sexual selection ( PDF ), in The American Naturalist , vol. 176, n. 2, 2010, pp. 141-148, DOI : 10.1086/653001 , PMID 20565262 (archiviato dall' url originale il 12 aprile 2016) .
- ^ SG Lucas, Spielmann, JA, Rinehart, LF e Martens, T., Dimetrodon (Amniota: Synapsida: Sphenacodontidae) from the Lower Permian Abo Formation, Socorro County, New Mexico ( PDF ), New Mexico Geological Society Guidebook, vol. 60, New Mexico Geological Society, 2009, pp. 281-284.
- ^ C. Sullivan, Reisz, RR e May, WJ, [0456:LDSEFT 2.0.CO;2 Large dissorophoid skeletal elements from the Lower Permian Richards Spur fissures, Oklahoma, and their paleoecological implications ], in Journal of Vertebrate Paleontology , vol. 20, n. 3, 2000, pp. 456-461, DOI : 10.1671/0272-4634(2000)020[0456:LDSEFT]2.0.CO;2 , JSTOR 4524117 .
- ^ a b EC Olson, Community evolution and the origin of mammals , in Ecology , vol. 47, n. 2, 1966, pp. 291-302, DOI : 10.2307/1933776 , JSTOR 1933776 .
- ^ A. Kriloff, Germain, D., Canoville, A., Vincent, P., Sache, M. e Laurin, M., Evolution of bone microanatomy of the tetrapod tibia and its use in palaeobiological inference , in Journal of Evolutionary Biology , vol. 21, n. 3, 2008, pp. 807-826, DOI : 10.1111/j.1420-9101.2008.01512.x , PMID 18312321 .
- ^ http://www.livescience.com/40842-finned-monster-chomped-heads-off-ancient-amphibians.html
Bibliografia
- Bramwell, CD and Fellgett, PP, 1973, Termoregolazione nelle "lucertole" munite di vela . Nature, v. 242, p. 203-205
Altri progetti
-
 Wikimedia Commons contiene immagini o altri file su Dimetrodon
Wikimedia Commons contiene immagini o altri file su Dimetrodon -
 Wikispecies contiene informazioni su Dimetrodon
Wikispecies contiene informazioni su Dimetrodon
Collegamenti esterni
- Dimetrodon , su sapere.it , De Agostini .
- ( EN ) Dimetrodon , su Enciclopedia Britannica , Encyclopædia Britannica, Inc.
- ( EN ) Dimetrodon , su Fossilworks.org .