Alcelaphus buselaphus
| Alcelafo | |
|---|---|
 | |
| Starea de conservare | |
 Risc minim [1] | |
| Clasificare științifică | |
| Domeniu | Eukaryota |
| Regatul | Animalia |
| Phylum | Chordata |
| Clasă | Mammalia |
| Ordin | Artiodactila |
| Subordine | Ruminantia |
| Familie | Bovidae |
| Subfamilie | Alcelaphinae |
| Tip | Alcelaphus de Blainville , 1816 |
| Specii | A. buselaphus |
| Nomenclatura binominala | |
| Alcelaphus buselaphus ( Pallas , 1776 ) | |
| Sinonime | |
Bubalis buselaphus | |
| Subspecii [2] | |
| |
Alcelafo ( Alcelaphus buselaphus Pallas , 1766 ) este o specie de antilopă de prerie africană, descrisă pentru prima dată de Peter Simon Pallas în 1766. Adulții măsoară puțin peste 1 m la greabăn. Masculii cântăresc 125-218 kg, iar femelele sunt puțin mai mici. Culoarea hainei variază în funcție de subspecii , de la cea de nisip a arlequinului vestic la cea aproape neagră a arlequinului Swayne . Coarnele sunt prezente la ambele sexe; masoara 45-70 cm lungime, iar forma lor variaza foarte mult de la o subspecie la alta. Alcelafi pot trăi 11-20 de ani în sălbăticie și până la 19 ani în captivitate.
Alcelafi sunt animale sociale care formează turme de 20-300 de indivizi. În general, liniștiți în dispoziție, Hareleps pot deveni agresivi atunci când sunt provocați. Dieta lor constă în principal din iarbă, la care se adaugă, în orice moment al anului, cantități mici de părți verzi și păstăi de plante din genul Hyparrhenia . Timpul reproducerii variază în funcție de anotimpuri și depinde atât de subspecii, cât și de populație. Alcelafii ajung la maturitate sexuală la vârsta de unu sau doi ani. După o perioadă de gestație de opt luni, se naște un singur copil. Harelonul trăiește în savane , zone împădurite și întinderi deschise.
Fiecare dintre cele opt subspecii ale șoimului are o stare de conservare diferită. Bubalusul a fost declarat dispărut de Uniunea Internațională pentru Conservarea Naturii (IUCN) în 1994. În trecut, tocul era prezent în mare parte din Africa, dar diferitele populații au suferit un declin drastic din cauza distrugerii. Habitat, vânătoare , extinderea așezărilor umane și concurența cu vitele domestice pentru hrană. Alcelafo este dispărut în Algeria, Egipt, Lesotho, Libia, Maroc, Somalia și Tunisia. A fost introdus în Swaziland și Zimbabwe. Este o pradă foarte populară pentru vânători datorită cărnii sale foarte apreciate.
Etimologie
„Alcelafo” este italianizarea termenului latin Alcelaphus , la rândul său o formă compusă derivată din grecescul ἄλκη, „moose” și ἔλαϕος, „cerb”. Denumirea în engleză a speciei, hartebeest , derivă în schimb din afrikaans-ul hertebeest [3] . Acest nume i-a fost atribuit de boeri , care observaseră asemănarea cu cerbul [4] . În olandeză , cuvântul hert înseamnă „căprioară”, iar beest înseamnă „fiară” [4] . Termenul a fost folosit pentru prima dată în literatura sud-africană în jurnalul Daghregister al administratorului colonial olandez Jan van Riebeeck în 1660. El a scris: „ Meester Pieter ein hart-beest geschooten hadde (Mr. Pieter [van Meerhoff] a tăiat un copac) " [5] .
Evoluţie
Genul Alcelaphus a apărut în urmă cu aproximativ 4,4 milioane de ani, într-o cladă care include și genurile Damalops , Numidocapra , Rabaticeras , Megalotragus , Oreonagor și Connochaetes [6] . O analiză efectuată pe baza aspectelor filogeografice a sugerat posibila apariție a speciei în Africa de Est. De acolo, se crede că s-a răspândit ulterior în restul continentului. Analizele filogenetice au arătat că prima și cea mai veche diversificare genetică a avut loc în populațiile sudice și nordice ale alcelafo-ului. Linia evolutivă nordică s-a împărțit la rândul său în alte două descendențe, estice și vestice, probabil ca urmare a extinderii centurii pădurii tropicale din Africa centrală și a contracției consecvente a habitatelor savanei în timpul unei perioade de încălzire globală. Aceste evenimente importante care au avut loc în timpul evoluției arlequinului sunt strâns legate de factorii climatici, care ar fi putut juca un rol de primă importanță în istoria evoluției speciei [7] . Au fost găsite rămășițe fosile de aleleafo roșu în Elandsfontein, Cornelia și Florisbad în Africa de Sud, precum și în Kabwe în Zambia [8] .
Alcelafo este bine reprezentat în situri care datează din epoca natufiană și neolitică , precum și din epoca bronzului și a fierului . În Israel, rămășițele harelamusului expuse pe teren deschis au fost găsite în Negev și în câmpiile Shephelah și Sharon . Cele mai recente fosile au fost găsite în Tel Lachish . În vremurile istorice, specia era limitată la întinderile deschise din cele mai sudice regiuni din sudul Levantului . După toate probabilitățile, arlequinul a fost vânat în Egipt și acest lucru ar fi putut duce la o scădere a numărului Levantului, separându-i de principala populație africană [9] .
Taxonomie
Descris pentru prima dată de zoologul și botanistul german Peter Simon Pallas în 1766, harelapterul este încă menționat prin numele său științific original, Alcelaphus buselaphus . Este singura specie din genul Alcelaphus [2] . Specia poate fi împărțită în trei grupe principale bazate pe structura craniului: grupa A. b. buselaphus (incluzând și A. b. major ), grupa A. b. tora (inclusiv A. b. swaynei , A. b. cokii și A. b. lelwel ) și grupul A. b. lichtensteinii (incluzând și A. b. caama ). Analize genetice mai detaliate indică o afinitate mai mare între grupurile A. B. buselaphus și A. b. tora [2] .
Poziția taxonomică a arborelui lui Lichtenstein a fost dezbătută de ceva timp. Zoologii Jonathan Kingdon și Theodor Haltenorth au considerat-o o subspecie a lui A. buselaphus [10] . În 1979, paleontologul Elisabeth Vrba a instituit un nou gen, Sigmoceros , special pentru acest alcelafo [11] , pe baza afinității sale mai mari cu genul Connochaetes ; mai târziu, în 1997, același savant, după studii suplimentare, a lăsat deoparte noul gen [12] . Analiza ADN-ului mitocondrial nu a găsit dovezi care să justifice înființarea unui gen separat. În cadrul aceleiași analize s-a arătat că subfamilia Alcelafini este monofiletică și s-a descoperit o afinitate strânsă între genurile Alcelaphus și Damaliscus - ambele din punct de vedere genetic și morfologic [13] .
Subspecii
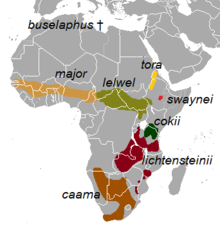
Mulți taxoni au fost considerați sintotipuri ale acestei specii și, din acest motiv, a fost necesar să se stabilească un lectotip . Cele șase specii de hawkfish recunoscute de vechii autori au fost ulterior considerate subspecii, când hibridizarea dintre unele dintre ele s-a dovedit posibilă [2] . Subspeciile recunoscute în prezent sunt următoarele [1] [2] :
- A. b. buselaphus † (Pallas, 1766) , bubala sau harelopa nordică [14] , a fost răspândită în trecut în nordul Africii. Ultimul exemplar a fost doborât în Algeria și a fost declarat dispărut în 1996 de IUCN [15] [16] .
- A. b. caama ( É. Geoffroy Saint-Hilaire , 1803) , hamul roșu sau Cape, este foarte popular printre pasionații de jocuri mari [17] . Este răspândit și comun în Africa, iar numărul exemplarelor este în creștere [18] . Uneori este considerată o specie separată, Alcelaphus caama [2] .
- A. b. cokii Günther, 1884, „ Coca-Cola sau congonii” este originară din Kenya și Tanzania [19] .
- A. b. lelwel ( Heuglin , 1877) , șoimul lelwel , este răspândit în Republica Centrafricană, sudul Ciadului, nord-estul Republicii Democrate Congo, sud-vestul Etiopiei, Kenya, Sudanul de Sud, extremitățile nord-vestice ale Tanzaniei și nordul Ugandei și vestul [ 20] . Scăderea drastică a populației a închis majoritatea indivizilor în ariile protejate [21] .
- A. b. lichtensteinii (Peters, 1849) , stăpânul Lichtenstein , locuiește în zonele împădurite de miombo din estul și sudul Africii. De asemenea, a fost considerată o specie separată [22] . Este originar din Angola, Republica Democrată Congo, Malawi, Mozambic, Africa de Sud, Tanzania, Zambia și Zimbabwe [23] .
- A. b. major (Blyth, 1869) , arlequinul vestic , este răspândit pe o fâșie de teritoriu între Senegal și nordul Camerunului [24] .
- A. b. swaynei (PL Sclater, 1892) , arlequinul lui Swayne , este adesea confundat cu arlequinul torei datorită aspectului său fizic foarte asemănător [25] . Este răspândit doar în Etiopia, iar numărul exemplarelor scade [26] .
- A. b. tora Grey, 1873 , alcelafo torà , este răspândit în nord-vestul Etiopiei și Eritreea [27] .

A. b. buselaphus .

A. b. caama .

A. b. cokii .

A. b. lichtensteinii .

A. b. swaynei .




Harelaphe-ul lui Jackson, un alt tip de harelope, nu are o poziție taxonomică bine definită. Primul exemplar cunoscut al acestui șoim a fost cel adăpostit la Grădina Zoologică din Bronx (SUA) în 1913 [28] . Este considerat un hibrid între lelwel și Coca-Cola. Cercetătorii IUCN / SSC Antelope Specialist Group (ASG) raportează că antilopele născute din încrucișările dintre un harelope lelwel kenyan și ugandez sunt identice ca aspect cu harelope Jackson [28] . Baza de date African Antelope (1998) consideră căprioara lui Jackson ca fiind un sinonim pentru lelwel-ul de copac [29] . Acest șoim este prezent în zonele în care zonele hamsterului lelwel și cocs se suprapun - în vestul Keniei și în districtul Karamoja (nord-vestul Ugandei) [28] . La vest de Nil este înlocuit de lelwel aleaf [30] .
Genetica și hibrizii
S-a găsit un grad ridicat de variabilitate genetică atât în populațiile de șoimi roșii, cât și în cele de Swayne din Sanctuarul Naturii Senkele și din Parcul Național Nechisar . Chiar și printre populațiile lui Swayne's Hawkbuck, cei de la Senkele Nature Sanctuary au prezentat o diversitate genetică mai mare decât cei de la Parcul Național Nechisar. Multe haplotipuri mitocondriale și alele microsatelite prezente cu frecvență ridicată printre exemplarele Senkele nu au fost prezente la indivizii Nechisar. Din acest motiv, programele de conservare și reproducere trebuie să țină seama de acest lucru pentru a menține pură diversitatea genetică a acestor populații [31] .
Numărul diploid de cromozomi din corpul ceresc este de 40. A fost posibil să se nască un mascul steril rezultat din hibridizarea dintre un roșu și un blesbok ( Damaliscus pygargus ), al cărui număr diploid de cromozomi este de 38. Se crede că sterilitatea hibridului se datorează problemelor de segregare apărute în timpul meiozei . Alte cauze posibile aduse pentru a explica această disfuncție sunt azoospermia și un număr mai mic de celule germinale în secțiunea transversală a tubilor seminiferi [32] .
Două forme hibride rezultate din încrucișarea între subspecii diferite sunt recunoscute de unele agenții care organizează vânătoare de vânat mare.
- Alcelaphus buselaphus lelwel × cokii . Hamul din Kenya Highland este rezultatul unei încrucișări între lelwel și hamul Coca-Cola. Comparativ cu arborele Coca-Cola, acest hibrid are o culoare mai deschisă și o dimensiune mai mare. Paltonul are o culoare ușoară de piele de căprioară, iar capul este mai lung decât cel al harelopei Coca-Cola. Coarnele, prezente la ambele sexe, sunt mai grele și mai lungi decât cele ale părinților. Răspândit în trecut de-a lungul tuturor zonelor de munte vestice ale Keniei, între Lacul Victoria și Muntele Kenya , astăzi este retrogradat în valea Lambwe (sud-vestul Keniei) și în Laikipia și zonele învecinate din centrul-vestul Keniei [33] .
- Alcelaphus buselaphus lelwel × swaynei . Neumann's ale-pod este numit în onoarea călătorului și sportivului AH Neumann. Este rezultatul încrucișării dintre lelwel și hamul lui Swayne. Potrivit zoologului american Edmund Heller ar fi o încrucișare între A. b. nakura , o subspecie descrisă de același autor și A. b. lelwel [34] . Fața este mai lungă decât cea a copacului lui Swayne. Culoarea hainei este maro-auriu, mai palidă spre regiunile inferioare. Barbia este negricioasă, iar smocul cozii este negru. Ambele sexe au coarne mai lungi decât harelopa lui Swayne. Coarnele se dezvoltă pentru a forma un „V” mărit și sunt diferite de cele din forma parantezelor lărgite ale hamului Swayne și de cele înguste în formă de „V” ale lelwel-ului. Răspândit în Etiopia, este prezent într-o mică zonă la est de râul Omo și la nord de lacul Turkana extins între zona de nord-est a lacului Chew Bahir și lacul Chamo din apropiere [35] .
Descriere

Harelapul are o înălțime de puțin peste 1 m la greabăn și măsoară 150–245 cm lungime [36] . Femelele cântăresc 116–185 kg, masculii 125–218 kg [37] . Coada, lungă de 30–70 cm, se termină cu un smoc negru [36] . Celelalte caracteristici principale ale arlequinului sunt picioarele lungi (care au adesea pete negre) [14] , gâtul scurt și urechile ascuțite [37] . Pe lângă fața lungă, harelopa se poate distinge de alte antilope și pentru pieptul mare și pentru spatele foarte înclinat, mai înalt la greabăn decât la crestă [3] . Poate trăi 11-20 de ani în sălbăticie și până la 19 ani în captivitate [36] . Alcelafo împărtășește unele caracteristici fizice cu damaliscul (genul Damaliscus ), cum ar fi fața îngustă și alungită, forma coarnelor, textura și culoarea hainei și smocul terminal al părului de coadă. Gnu , pe de altă parte, are un craniu și coarne mai specializate decât hamul [38] .
Învelișul este în general scurt și strălucitor [38] . Colorarea sa variază în funcție de subspecie; alcelafo mare de vest are o mantie ușoară și uniformă, de culoare maro-nisip [24] , în timp ce cea a lucernei tora este întunecată [27] . Arborele roșu, așa cum indică și numele său, are un strat complet de această culoare [39] . Arborele Coca-Cola este roșiatic-brunet în regiunile superioare și de culoare mai deschisă în cele inferioare [40] . Arborele lelwel este maro roșiatic [20] . În corpul lui Lichtenstein, regiunile superioare sunt maroniu-roșiatic, dar flancurile sunt maronii, iar posteriorul albicios [41] . De asemenea, are dungi întunecate pe picioarele din față [14] [41] . Harlands-urile Swayne și Torah sunt foarte asemănătoare ca aspect și ambele au capete mici, haine întunecate și coarne similare. Harelaphe-ul lui Swayne este cel mai mic dintre cele două și are coarne puțin mai scurte și mai grele [25] [38] . Firele de păr care acoperă corpul, de textură fină, au o lungime de aproximativ 25 mm [11] . Arborele are glande preorbitale cu un canal central. Ele secretă un fluid întunecat cu miros urât în alehaphus de Coca-Cola și Lichtenstein, în timp ce în lelwel produc o secreție incoloră [38] .
La toate subspeciile ambele sexe au coarne, dar cele ale femelelor sunt mai subțiri [37] . Coarnele pot atinge 45-70 cm lungime [36] . Sunt ușor curbate la exterior și, spre vârf, la interior. Aproape toată partea bazală a coarnelor are inele caracteristice [37] . Forma coarnelor variază de la o subspecie la alta. Arborele roșu are coarne în formă de „Z” [39] , în timp ce cel al lui Lichtenstein are forma unui „S” ridat [41] . Ambele harelape ale lui Swayne și Torah au coarne în formă de liră [27] . Cele din lelwel alcelafo sunt groase și în formă de „V” [20], iar cele ale alehafoum de Coca-Cola sunt scurte, groase și în formă de paranteze [40] . Halterul vestic are impunătoare coarne în formă de „U” [24] . Coarnele sunt folosite pentru apărarea împotriva prădătorilor și în luptele dintre masculi pentru dominație în timpul sezonului de împerechere [42] .
Alcelafo prezintă un dimorfism sexual puțin marcat, deoarece ambele sexe au coarne și sunt de dimensiuni similare. Gradul de dimorfism sexual variază în funcție de subspecie. Bărbații cântăresc cu 8% mai mult decât femelele în hamurile lui Swayne și Lichtenstein și cu 23% mai grele în hamurile roșii. Într-un studiu, cel mai mare dimorfism s-a găsit în greutatea craniului [43] . Într-un alt studiu, lungimea sezonului de împerechere a fost corelată cu înălțimea pedunculilor (structurile osoase pe care cresc coarnele) și greutatea craniului, precum și cu circumferința coarnelor [42] .
Biologie

La fel ca majoritatea antilopelor, harelopa este un animal de zi . Pășește dimineața devreme și după-amiaza târziu și se odihnește la umbră în cele mai fierbinți ore ale zilei. Este un animal social și formează turme de până la 300 de exemplare. Cele mai numeroase efective se întâlnesc în locurile cele mai bogate în iarbă [37] . Cea mai mare turmă despre care știm era 10.000 de animale. Membrii unei haite pot fi împărțiți în patru grupuri: bărbați adulți teritoriali, bărbați adulți neteritoriali, bărbați juvenili și femele cu tineri. Femelele formează grupuri de 5-12 animale, în care pot fi găsite până la patru generații de tineri. Femelele luptă pentru dominarea turmei [44] . Certurile între bărbați și femele sunt frecvente [28] . La vârsta de trei sau patru ani, bărbații pot încerca să câștige supremația asupra unui teritoriu și a unei efective de femele. Un bărbat rezident își apără propriul teritoriu și își poate ataca semenii dacă este provocat [43] . Masculul marchează limitele teritoriului cu mormane de excremente [28] . Începutul unei lupte este marcat de o serie de mișcări și posturi ale capului, precum și de depunerea excrementelor în grămezi speciale. Adversarii cad în genunchi și, după ce se lovesc reciproc cu capul, încep să lupte, cu coarnele lipite. Fiecare încearcă să îndoaie capul adversarului într-o parte pentru a înjunghia gâtul și umerii cu coarnele [43] . În general, bărbații își pierd controlul asupra teritoriului lor după șapte sau opt ani [36] . Cazul documentat al unui cap chel al copacului care sparte un biciclist a fost interpretat ca un comportament teritorial [45] .
În timp ce pășune, un exemplar rămâne în pază, urcând adesea pe o movilă de termite pentru a putea privi mai departe. În momentele de pericol, întreaga turmă scapă aranjându-se într-un singur dosar imediat ce primul exemplar începe să ruleze [44] . Arlechinul este mai alert și mai precaut decât alți ungulați [46] . Alcelafi adulți sunt pradați de lei , leoparzi , hiene și câini sălbatici ; ghepardii și șacalii capturează doar tineri [44] . Atât alcelafi, cât și damaliscii produc zgomote ușoare și mormăi. Arlechinul folosește movilele excrementelor ca semnal olfactiv și vizual [38] . Turmele migrează numai în perioade de extremă necesitate, cum ar fi în timpul dezastrelor naturale și al secetei [47] . Alcelafo este cel mai permanent dintre Alcelafini [38] . De asemenea, consumă mai puțină apă și are cea mai mică rată metabolică dintre toți membrii subfamiliei [38] .
Paraziți
Diferiti paraziți au fost izolați de alchimist . Un șoim roșu din parcul național Kalahari Gemsbok adăpostea specii de Cooperia , Impalaia nudicollis , Parabronema și Trichostrongylus [48] . Exemplare de Estrini au fost găsite în nouă din alechifere ale lui Lichtenstein. Larve aparținând genurilor Gedoelstia, Oestrus și Kirkioestrus au fost izolate din nazale carii și paranazale sinusurilor. Au fost găsite maximum 252 de larve în capul unui singur animal, dar nu s-a găsit nicio patogenitate [49] . În Gobabis (Africa de Sud-Vest) un șoim roșu a fost găsit infestat cu viermi lungi și subțiri. Acestea au fost numite Longistrongylus meyeri în onoarea descoperitorului lor, T. Meyer, și clasificate în genul Longistrongylus [50] . Într-un alt caz, un șoim roșu a fost afectat de o teileroză din cauza paraziților Rhipicephalus evertsi și ai genului Theileria [51] . Paraziții copacilor pot fi găsiți și în gazele și gnu [52] . La sud de Sahara, lucerna poate fi infestată cu Loewioestrus variolosus , Gedoelstia cristata și G. hassleri . Ultimele două specii pot provoca boli grave, cum ar fi „boala ochiului bombat”, care poate duce la encefalită [53] . În anii 1960, Robustostrongylus aferensis , un nematod abomasum , a fost descoperit într-un kongoni din Uganda [54] . Nematodele precum Haemonchus contortus , Trichostrongylus axei și Cooperia curticei , tenii precum Moniezia expansa , Avitellina centripunctata și Stilesia globipunctata și paramphistomele precum Setaria labiato- papillosa au fost găsite în tractul digestiv al unui copac occidental [55] .
Dietă
Alcelafi sunt erbivore, iar dieta lor constă în principal din iarbă [56] . Într-un studiu realizat în ferma de joc Nazinga din Burkina Faso, s-a constatat că structura craniului copacului facilitează achiziționarea și mestecarea alimentelor cu fibre ridicate. Comparativ cu antilopa roană, harelopa este mai bine adaptată pentru obținerea și mestecarea regresului rar al ierburilor perene în perioadele în care furajele sunt mai puțin disponibile. Iarba reprezintă în general cel puțin 80% din dieta cu lucernă, dar în timpul sezonului uscat, din octombrie până în mai, această valoare poate crește cu peste 95%. Jasminium kerstingii face parte din dieta arlequin la începutul sezonului ploios. Între cele două sezoane, alcelafi se hrănesc în principal cu iarbă din genul Culms . În toate perioadele anului consumă cantități mici de părți verzi și păstăi de plante din genul Hyparrhenia [57] . Arlequinul poate digera o cantitate mai mare de alimente decât alte bovide [58] . În zonele în care apa este puțină, poate mânca pepeni, rădăcini și tuberculi [38] .
Într-un studiu menit să stabilească selectivitatea alimentară a gnu-ului, a zebrelor și a cocosului, acesta din urmă sa dovedit a fi cel mai selectiv. Toate cele trei animale au preferat Themeda triandra decât Pennisetum mezianum și Digitaria macroblephara . Au fost consumate mai multe specii de iarbă în timpul sezonului uscat decât în sezonul ploios [59] .
Reproducere

Alcelafi se pot împerechea în orice moment al anului. Spicile pot fi afectate de disponibilitatea alimentelor [56] . Atât bărbații, cât și femelele ating maturitatea sexuală la vârsta de unu sau doi ani. Reproducerea variază în funcție de subspecie și populație în perioada de împerechere [36] . Împerecherea are loc în teritoriile apărate de un singur mascul, mai ales în zone deschise ale zonelor înalte sau pe creste [56] . Bărbații pot lupta cu înverșunare pentru dominație [43] . Masculul dominant sniffs organele genitale feminine și, în cazul în care ea este în călduri , ea urmează. Uneori, o femelă flutură ușor coada pentru a semnala că este în căldură [38] . Masculul încearcă să blocheze calea pentru femelă. Când se oprește, permite masculului să-l monteze. Împerecherea are loc rapid și se repetă adesea, chiar de două sau mai multe ori pe minut [38] . În această perioadă, orice intrus este alungat [44] . La turme mari, femela se împerechează cu diverși masculi [38] . Gestația durează aproximativ 240 de zile, după care se naște un singur copil. Nou-născutul cântărește aproximativ 9 kg. Femelele nasc în tufiș, spre deosebire de cele de gnu, care nasc în grupuri în câmpie. Bebelușul este înțărcat la patru luni [36] . Bărbații tineri își însoțesc mamele timp de doi ani și jumătate, mai mult decât alți Alcelafini [38] .
Distribuție și habitat
Alcelafi locuiesc savane aride și pajiști împădurite [11] și se mută adesea în zone mai uscate după precipitații [37] . Sunt mai toleranți față de zonele împădurite decât alte Alcelafini și sunt adesea întâlnite la marginile pădurilor [56] . Pe Muntele Kenya s-au găsit alcelafi până la 4000 m altitudine [1] . Șoimul roșu se știe că se deplasează pe zone întinse, iar femelele călătoresc pe teritorii de peste 1000 km², în timp ce teritoriile masculilor măsoară aproximativ 200 km² în extensie [60] . În Parcul Național Nairobi (Kenya), femelele ocupă teritorii individuale de 3,7-5,5 km², care nu sunt asociate în mod special cu niciun grup de femele. În medie, teritoriile femelelor sunt suficient de mari pentru a le include chiar și pe cele de 20-30 de bărbați [14] .
În trecut, alchimistul era foarte frecvent în Africa. Numărul de exemplare a scăzut dramatic din cauza distrugerii habitatului, vânătorii, extinderii așezărilor umane și concurenței pentru hrana cu vitele domestice [1] [36] . Mărimea diferitelor subspecii s-a corelat cu productivitatea habitatului și cu precipitațiile [61] . Alcelafo este prezent în Angola, Benin , Botswana , Burkina Faso, Camerun, Republica Centrafricană, Ciad, Republica Democrată Congo, Eritreea, Etiopia, Gambia , Ghana , Guineea , Guineea-Bissau , Coasta de Fildeș , Kenya, Mali , Namibia , Niger , Nigeria , Senegal, Africa de Sud, Sudan, Sudan de Sud, Tanzania, Togo și Uganda . Este dispărut în Algeria, Egipt, Lesotho, Libia, Maroc, Somalia și Tunisia și a fost introdus în Swaziland și Zimbabwe [1] . Gama diferitelor subspecii ale arlequinului diferă foarte mult între ele. După ce a fost reintrodus în ariile și fermele protejate, șoimul roșu a devenit foarte popular și este singurul heli-șoim cu o populație în creștere [1] . Este răspândit în aproape toată Africa de Sud [1] . Toate subspecii de arlequin, cu excepția arlequinului roșu (cu o populație în creștere) [18] și arlequinului lui Lichtenstein (cu o populație stabilă) [23] , sunt în declin și trei dintre ele sunt în pericol de dispariție: alcelafo torà, lelwel-ul și cel al lui Swayne. Torah aleholic este limitat la Eritreea și Etiopia, Swayne aleholic la patru arii protejate, iar lelwel aleholic la câteva zone protejate [1] .
depozitare

Fiecare subspecie a șoimului este clasificată într-o stare de conservare diferită de către Uniunea Internațională pentru Conservarea Naturii (IUCN). Cu toate acestea, în general, specia este clasificată printre cele cu „cel mai mic risc” [1] . Arborele roșu este cel mai răspândit, iar după reintroducerea sa în zone protejate și private numărul de exemplare crește [1] . Clasificată drept o specie cu „cel mai mic risc”, populația sa este estimată la peste 130.000 de exemplare [18] , în mare parte găsite în sudul Africii [60] . Bubalul a fost declarat dispărut în 1994 [15] . Exploratorul german Heinrich Barth , în scrierile sale din 1857, a citat armele de foc și intruziunea europenilor printre motivele care stau la baza scăderii numărului de exemplare ale acestei subspecii [62] . A dispărut în Tunisia la sfârșitul secolului al XIX-lea [63] și ultimul exemplar a fost doborât între 1945 și 1954 în Algeria [15] .
Arborele de cocs este în prezent clasificat ca o specie cu „cel mai mic risc”. Il numero di appartenenti a questa sottospecie è diminuito notevolmente a causa della distruzione dell'habitat, e oggi sono presenti circa 42.000 esemplari nella regione del Mara , nel parco nazionale del Serengeti e nel parco nazionale del Tarangire in Tanzania e nel parco nazionale dello Tsavo orientale in Kenya. La popolazione è in diminuzione, e il 70% degli esemplari vive in aree protette [64] . L'alcelafo occidentale è considerato «prossimo alla minaccia»; ne rimangono circa 36.000 esemplari. Più del 95% degli esemplari vive in aree protette (come il Parco nazionale del Comoé ) o nei loro dintorni, ma il numero di individui è in diminuzione perfino in queste zone [65] . L'alcelafo di Lichtenstein è attualmente considerato a «rischio minimo», ed è presente in aree protette quali la Riserva di caccia del Selous e allo stato selvatico in Tanzania meridionale e occidentale e nello Zambia [23] .
Le sottospecie più minacciate sono l'alcelafo torà, lelwel e di Swayne. L'alcelafo torà è classificato tra le specie « in pericolo critico », dal momento che ne rimangono meno di 250 esemplari adulti. Probabilmente scomparso in Sudan, sopravvive in numero ridotto in Eritrea ed Etiopia [66] . L'alcelafo di Swayne è classificato tra le specie «in pericolo», ma rischia di essere riclassificato tra quelle «in pericolo critico». Ne rimangono in tutto meno di 600 esemplari, tra cui circa 250 esemplari adulti, confinati in quattro aree protette principali: il santuario naturale di Senkele, il parco nazionale di Nechisar, il parco nazionale d'Awash e il parco nazionale di Mazie [67] . Gli alcelafi di Senkele sono costretti a competere con gli animali domestici del popolo Oromo [26] . Uno studio effettuato nel parco nazionale di Nechisar nel 2009 e 2010 ha indicato il notevole incremento del bestiame degli Oromo (aumentato del 49,9% e del 56,5% nel 2006 e nel 2010, rispettivamente), lo sfruttamento illegale di risorse naturali e la perdita dell'habitat come maggiori minacce per la sopravvivenza delle popolazioni di alcelafi di Swayne ivi presenti [68] . L'alcelafo lelwel è considerato « in pericolo », e il numero di esemplari è diminuito notevolmente dagli anni ottanta, quando se ne contavano oltre 285.000. All'epoca era diffuso prevalentemente nella Repubblica Centrafricana e nell'attuale Sudan del Sud [64] . Oggi ne rimangono meno di 70.000 [21] . Questo alcelafo è presente in alcune zone dell'Omo meridionale, in Etiopia [69] .
Importanza economica
Gli alcelafi sono prede molto popolari tra gli appassionati di caccia grossa e di trofei a causa della loro carne, che è molto apprezzata. Pacchetti di viaggio per la caccia all'alcelafo sono disponibili online [36] . L'alcelafo è facile da cacciare a causa della sua visibilità [44] . In uno studio effettuato sugli esemplari abbattuti, tenendo conto del luogo e del sesso degli animali, il peso medio di carne ricavata dai maschi di alcelafo rosso catturati era di 79,3 kg e quello della carne ricavata dalle femmine di 56 kg. La carne degli animali provenienti dalla regione di Qua-Qua presentava il maggiore contenuto lipidico - 1,3 g ogni 100 g di carne. Differenze trascurabili vennero riscontrate nelle concentrazioni individuali di acidi grassi , amminoacidi e sali minerali . Lo studio considerò la carne di alcelafo molto sana, dal momento che presentava un rapporto tra acidi grassi polinsaturi e saturi di 0,78, leggermente superiore allo 0,7 raccomandato [70] .
Nel corso di uno studio del 2013 sono stati analizzati campioni di carne di selvaggina in vendita in supermercati, all'ingrosso e in altri punti vendita. Lo studio ha rivelato che alcuni tipi di biltong di «cudù», « antilope saltante » o « struzzo » contenevano in realtà carne di alcelafo. Su 146 etichette, 100 presentavano una dicitura erronea, il che ha rivelato un problema importante nell'etichettatura della carne in Sudafrica [71] .
È nota la proverbiale alta vitalità delle specie africane cacciabili, soprattutto nell'ambito delle varie antilopi. L'alcefalo (o "hartebeest" come è più comunemente conosciuto in lingua inglese), è una delle antilopi più resistenti ai colpi di arma da fuoco, per cui la sua caccia richiede calibri robusti e non di rado più colpi per l'abbattimento.
Note
- ^ a b c d e f g h i j ( EN ) Mallon, DP (Antelope Red List Authority) & Hoffmann, M. (Global Mammal Assessment) 2008, Alcelaphus buselaphus , su IUCN Red List of Threatened Species , Versione 2020.2, IUCN , 2020.
- ^ a b c d e f DE Wilson and DM Reeder, Mammal Species of the World: A Taxonomic and Geographic Reference , 3ª ed., Baltimore, Maryland, Johns Hopkins University Press, 2005, p. 674, ISBN 978-0-8018-8221-0 .
- ^ a b MA Mares, Encyclopedia of Deserts , Norman, Oklahoma, University of Oklahoma Press, 1999, p. 265, ISBN 978-0-8061-3146-7 .
- ^ a b EC Llewellyn, Chapter XIV The Influence of South African Dutch or Afrikaans on the English Vocabulary , in The Influence of Low Dutch on the English Vocabulary , London, Oxford University Press, 1936, p. 163.
- ^ JD Skinner and CT Chimimba, The Mammals of the Southern African Subregion , 3ª ed., Cambridge, Cambridge University Press, 2005, p. 649, ISBN 978-0-521-84418-5 .
- ^ J. Harris and M. Leakey, Lothagam: The Dawn of Humanity in Eastern Africa , New York, Columbia University Press, 2001, p. 547, ISBN 978-0-231-11870-5 .
- ^ Ø. Flagstad, PO Syvertsen, NC Stenseth and KS Jakobsen, Environmental change and rates of evolution: the phylogeographic pattern within the hartebeest complex as related to climatic variation , in Proceedings of the Royal Society of London B , 268 (1468), 2001, pp. 667–77, DOI : 10.1098/rspb.2000.1416 , PMC 1088655 , PMID 11321054 .
- ^ LR Berger and B. Hilton-Barber, Field Guide to the Cradle of Humankind: Sterkfontein, Swartkrans, Kromdraai & Environs World Heritage Site , 2ª ed., Cape Town, Struik, 2004, p. 163, ISBN 978-1-77007-065-3 .
- ^ E. Tsahar, I. Izhaki, S. Lev-Yadun, G. Bar-Oz and DM Hansen, Distribution and extinction of ungulates during the Holocene of the southern Levant , in PLoS ONE , 4 (4), 2009, e5316, DOI : 10.1371/journal.pone.0005316 .
- ^ DE Wilson, Mammal Species of the World: A Taxonomic and Geographic Reference , 2005, p. 675.
- ^ a b c RM Nowak,Walker's Mammals of the World , 6ª ed., Baltimore, Maryland, Johns Hopkins University Press, 1999, pp. 1181–3, ISBN 978-0-8018-5789-8 .
- ^ C. Groves and P. Grubb, Ungulate Taxonomy , Baltimore, Maryland, Johns Hopkins University Press, 2011, p. 208, ISBN 978-1-4214-0093-8 .
- ^ C. Matthee and TJ Robinson, Cytochrome b phylogeny of the family Bovidae: resolution within the Alcelaphini, Antilopini, Neotragini, and Tragelaphini , in Molecular Phylogenetics and Evolution , 12 (1), 1992, pp. 31–46, DOI : 10.1006/mpev.1998.0573 , PMID 10222159 .
- ^ a b c d David Macdonald, The Encyclopedia of Mammals , New York, Facts on File, 1987, pp. 564–71, ISBN 0-87196-871-1 .
- ^ a b c Mallon, DP (Antelope Red List Authority) & Hoffmann, M. (Global Mammal Assessment) 2008, Alcelaphus buselaphus buselaphus in IUCN Red List of Threatened Species , Versione 2014.3, IUCN, 2014.
- ^ DP Mallon and SC Kingswood, Antelopes: North Africa, the Middle East, and Asia , Gland, Switzerland, IUCN, 2001, p. 25, ISBN 978-2-8317-0594-1 .
- ^ Trophy Hunting Red Hartebeest , su African Sky Safaris and Tours , African Sky. URL consultato il 20 gennaio 2013 .
- ^ a b c Mallon, DP (Antelope Red List Authority) & Hoffmann, M. (Global Mammal Assessment) 2008, Alcelaphus buselaphus caama in IUCN Red List of Threatened Species , Versione 2014.3, IUCN, 2014.
- ^ JO Kokwaro and T. Johns, Luo Biological Dictionary , Nairobi, East African Educational Publishers, 1998, p. 217, ISBN 978-9966-46-841-3 .
- ^ a b c Lelwel Hartebeest , su Safari Club International , SCI Online Record Book. URL consultato il 19 gennaio 2013 .
- ^ a b Mallon, DP (Antelope Red List Authority) & Hoffmann, M. (Global Mammal Assessment) 2008, Alcelaphus buselaphus lelwel in IUCN Red List of Threatened Species , Versione 2014.3, IUCN, 2014.
- ^ J. Rafferty, Grazers , 1ª ed., New York, Britannica Educational Publications, 2010, p. 121, ISBN 978-1-61530-465-3 .
- ^ a b c Mallon, DP (Antelope Red List Authority) & Hoffmann, M. (Global Mammal Assessment) 2008, Alcelaphus buselaphus lichtensteinii in IUCN Red List of Threatened Species , Versione 2014.3, IUCN, 2014.
- ^ a b c Western Hartebeest , su Safari Club International , SCI Online Record Book. URL consultato il 19 gennaio 2013 .
- ^ a b Swayne Hartebeest , su Safari Club International , SCI Online Record Book. URL consultato il 19 gennaio 2013 .
- ^ a b JG Lewis and RT Wilson, The Plight of Swayne's Hartebeest , in Oryx , 13 (5), 1977, pp. 491–4, DOI : 10.1017/S0030605300014551 .
- ^ a b c A. Hildyard, Endangered Wildlife and Plants of the World , New York, Marshall Cavendish, 2001, pp. 674–5, ISBN 0-7614-7199-5 .
- ^ a b c d e S. Shurter and D. Beetem, Jackson's hartebeest ( Alcelaphus buselaphus jacksoni ) ( PDF ), su antelopetag.com , Antelope & Giraffe Tag. URL consultato il 28 aprile 2013 (archiviato dall'url originale il 4 luglio 2013) .
- ^ R. East,African Antelope Database 1998 , IUCN, 1999, p. 190, ISBN 978-2-8317-0477-7 .
- ^ P. Briggs and A. Roberts, Uganda: The Bradt Travel Guide , 6ª ed., Chalfont St. Peter, Bradt Travel Guides, 2010, p. 44, ISBN 978-1-84162-309-2 .
- ^ Ø. Flagstad, PO Syvertsen, N. ChR. Stenseth, JE Stacy, I. Olsaker, KH Røed and KS Jakobsen, Genetic variability in Swayne's hartebeest, an endangered antelope of Ethiopia , in Conservation Biology , 14 (1), 2000, pp. 254–64, DOI : 10.1046/j.1523-1739.2000.98339.x .
- ^ TJ Robinson, DJ Morris and N. Fairall, Interspecific hybridization in the bovidae: Sterility of Alcelaphus buselaphus × Damaliscus dorcas F1 progeny , in Biological Conservation , 58 (3), 1991, pp. 345–56, DOI : 10.1016/0006-3207(91)90100-N .
- ^ Kenya Highland Hartebeest , su Safari Club International , SCI Online Record Book. URL consultato il 18 gennaio 2013 .
- ^ AE Ruxton and E. Schwarz, On hybrid hartebeests and on the distribution of the Alcelaphus buselaphus group , in Proceedings of the Zoological Society of London , 99 (3), 2010, pp. 567–83, DOI : 10.1111/j.1469-7998.1929.tb07706.x .
- ^ Neumann's Hartebeest , su Safari Club International , SCI Online Book Record. URL consultato il 18 gennaio 2013 .
- ^ a b c d e f g h i K. Batty, Alcelaphus buselaphus , su University of Michigan Museum of Zoology , Animal Diversity Web. URL consultato il 22 gennaio 2013 .
- ^ a b c d e f Hartebeest fact file , su Wildscreen , Arkive. URL consultato il 27 gennaio 2013 (archiviato dall' url originale il 12 maggio 2014) .
- ^ a b c d e f g h i j k l RD Estes, The Behavior Guide to African Mammals: Including Hoofed Mammals, Carnivores, Primates , 4ª ed., Berkeley, University of California Press, 2004, pp. 133–42, ISBN 978-0-520-08085-0 .
- ^ a b M. Firestone, Watching Wildlife: Southern Africa; South Africa, Namibia, Botswana, Zimbabwe, Malawi, Zambia , 2ª ed., Footscray, Lonely Planet, 2009, pp. 228–9, ISBN 978-1-74104-210-8 .
- ^ a b Coke's Hartebeest , su Safari Club International , SCI: Online Record Book. URL consultato il 24 gennaio 2013 .
- ^ a b c Lichtenstein Hartebeest , su Safari Club International , SCI: Online Record Book. URL consultato il 22 marzo 2013 .
- ^ a b I. Capellini and LM Gosling, The evolution of fighting structures in hartebeest , in Evolutionary Ecology Research , vol. 8, 2006, pp. 997–1011.
- ^ a b c d I. Capellini, Dimorphism in the hartebeest , in Sex, Size and Gender Roles , 2007, pp. 124–32, DOI : 10.1093/acprof:oso/9780199208784.003.0014 , ISBN 978-0-19-920878-4 .
- ^ a b c d e J. Kingdon, East African Mammals: An Atlas of Evolution in Africa (Volume 3, Part D: Bovids) , Chicago, University of Chicago Press, 1989, ISBN 0-226-43725-6 .
- ^ FAIL LAB Episode One: Evolution, Featuring Professor Theodore Garland, Jr. , su youtube.com .
- ^ GB Schaller, The Serengeti Lion: A Study of Predator-Prey Relations (Pbk. ed.) , Chicago, University of Chicago Press, 1976, pp. 461–5, ISBN 978-0-226-73640-2 .
- ^ A. Verlinden, Seasonal movement patterns of some ungulates in the Kalahari ecosystem of Botswana between 1990 and 1995 , in African Journal of Ecology , 36 (2), 1998, pp. 117–28, DOI : 10.1046/j.1365-2028.1998.00112.x .
- ^ J. Boomker, IG Horak and V. De Vos, The helminth parasites of various artiodactylids from some South African nature reserves , in The Onderstepoort Journal of Veterinary Research , 53 (2), 1986, pp. 93–102, PMID 3725333 .
- ^ ( EN ) GW Howard, Prevalence of nasal bots (Diptera: Oestridiae) in some Zambian hartebeest , in Journal of Wildlife Diseases , 13 (4), 1977, p. 400–4, DOI : 10.7589/0090-3558-13.4.400 . URL consultato il 9 novembre 2020 (archiviato dall' url originale il 15 aprile 2013) .
- ^ PL le Roux, On Longistrongylus meyeri gen. and sp. nov., a trichostrongyle parasitizing the Red Hartebeest Bubalis caama , in Journal of Helminthology , 9 (3), 1931, p. 141, DOI : 10.1017/S0022149X00030376 .
- ^ E. Spitalska, M. Riddell, H. Heyne and OAE Sparagano, Prevalence of theileriosis in Red Hartebeest ( Alcelaphus buselaphus caama ) in Namibia , in Parasitology Research , 97 (1), 1995, pp. 77–9, DOI : 10.1007/s00436-005-1390-y , ISSN 1432-1955 , PMID 15986252 .
- ^ FRN Pester and BR Laurence, The parasite load of some African game animals , in Journal of Zoology , 174 (3), 1974, pp. 397–406, DOI : 10.1111/j.1469-7998.1974.tb03167.x .
- ^ CA Spinage, African Ecology - Benchmarks and Historical Perspectives , Berlin, Springer, 2012, p. 1176, ISBN 978-3-642-22872-8 .
- ^ EP Hoberg, A. Abrams and PA Pilitt, Robustostrongylus aferensis gen. nov. et sp. nov. (Nematoda: Trichostrongyloidea) in kob ( Kobus kob ) and hartebeest ( Alcelaphus buselaphus jacksoni ) (Artiodactyla) from Sub-Saharan Africa, with further ruminations on the Ostertagiinae , in Journal of Parasitology , 95 (3), 2009, pp. 702–717, DOI : 10.1645/GE-1859.1 , ISSN 1937-2345 .
- ^ AMG Belem and É. U. Bakoné, Gastro-intestinal parasites of antelopes and buffaloes ( Syncerus caffer brachyceros ) from the Nazinga game ranch in Burkina Faso , in Biotechnologie, Agronomie, Société et Environnement , 13 (4), 2009, pp. 493–8, ISSN 1370-6233 .
- ^ a b c d Hartebeest , su awf.org , African Wildlife Foundation. URL consultato il 20 gennaio 2013 .
- ^ JR Schuette, DM Leslie, RL Lochmiller and JA Jenks, Diets of hartebeest and roan antelope in Burkina Faso: support of the long-faced hypothesis , in Journal of Mammalogy , 79 (2), 1998, pp. 426–36, DOI : 10.2307/1382973 .
- ^ MG Murray, Comparative nutrition of wildebeest, hartebeest and topi in the Serengeti , in African Journal of Ecology , 31 (2), 1993, pp. 172–7, DOI : 10.1111/j.1365-2028.1993.tb00530.x .
- ^ RL Casebeer and GG Koss, Food habits of wildebeest, zebra, hartebeest and cattle in Kenya Masailand , in African Journal of Ecology , 8 (1), 1970, pp. 25–36, DOI : 10.1111/j.1365-2028.1970.tb00827.x .
- ^ a b G. Mills and L. Hes, The Complete Book of Southern African Mammals , Cape Town, Struik Publishers, 1997, p. 255, ISBN 978-0-947430-55-9 .
- ^ I. Capellini and LM Gosling, Habitat primary production and the evolution of body size within the hartebeest clade , in Biological Journal of the Linnean Society , 92 (3), 2007, pp. 431–40, DOI : 10.1111/j.1095-8312.2007.00883.x .
- ^ PR Yadav, Vanishing and Endangered Species , New Delhi, Discovery Publishing House, 2004, pp. 139–40, ISBN 978-81-7141-776-6 .
- ^ DP Mallon and SC Kingswood, Antelopes: North Africa, the Middle East, and Asia , Gland, Switzerland, IUCN, 2001, ISBN 978-2-8317-0594-1 .
- ^ a b R. East and the IUCN/SSC Antelope Specialist Group,African Antelope Database 1998 , Gland, Switzerland, The IUCN Species Survival Commission, 1999, ISBN 978-2-8317-0477-7 .
- ^ Mallon, DP (Antelope Red List Authority) & Hoffmann, M. (Global Mammal Assessment) 2008, Alcelaphus buselaphus major in IUCN Red List of Threatened Species , Versione 2014.3, IUCN, 2014.
- ^ Mallon, DP (Antelope Red List Authority) & Hoffmann, M. (Global Mammal Assessment) 2008, Alcelaphus buselaphus tora in IUCN Red List of Threatened Species , Versione 2014.3, IUCN, 2014.
- ^ Mallon, DP (Antelope Red List Authority) & Hoffmann, M. (Global Mammal Assessment) 2008, Alcelaphus buselaphus swaynei in IUCN Red List of Threatened Species , Versione 2014.3, IUCN, 2014.
- ^ D. Datiko and A. Bekele, Population status and human impact on the endangered Swayne's hartebeest (Alcelaphus buselaphus swaynei) in Nechisar Plains, Nechisar National Park, Ethiopia , in African Journal of Ecology , 49 (3), 2011, pp. 311–9, DOI : 10.1111/j.1365-2028.2011.01266.x .
- ^ P. Briggs, Ethiopia , 6ª ed., Chalfont St. Peter, Bradt Travel Guides, 2013, p. 60, ISBN 978-1-84162-414-3 .
- ^ LC Hoffman, K. Smit and N. Muller, Chemical characteristics of red hartebeest ( Alcelaphus buselaphus caama ) meat , in South African Journal of Animal Science , 40 (3), 2010, pp. 221–8, DOI : 10.4314/sajas.v40i3.6 .
- ^ BioMed Central, Problems with identifying meat? The answer is to check the barcode , su biomedcentral.com , BioMed Central, 2013. URL consultato il 10 marzo 2013 .
Altri progetti
-
 Wikimedia Commons contiene immagini o altri file su Alcelaphus buselaphus
Wikimedia Commons contiene immagini o altri file su Alcelaphus buselaphus -
 Wikispecies contiene informazioni su Alcelaphus buselaphus
Wikispecies contiene informazioni su Alcelaphus buselaphus
Collegamenti esterni
- ( EN ) Alcelaphus buselaphus , su Enciclopedia Britannica , Encyclopædia Britannica, Inc.
- ( EN ) Alcelaphus buselaphus , su Fossilworks.org .
| Controllo di autorità | LCCN ( EN ) sh85059111 |
|---|