Deinocheirus mirificus
| Deinocheirus | |
|---|---|
 | |
| Starea de conservare | |
Fosil | |
| Clasificare științifică | |
| Domeniu | Eukaryota |
| Regatul | Animalia |
| Phylum | Chordata |
| Clasă | Sauropsida |
| Superordine | Dinozauria |
| Subordine | Theropoda |
| Superfamilie | † Ornitomimosaurie |
| Familie | † Deinocheiridae |
| Tip | † Deinocheirus Osmólska și Roniewicz, 1970 |
| Nomenclatura binominala | |
| † Deinocheirus mirificus Osmólska & Roniewics, 1970 | |
Deinocheirus (al cărui nume înseamnă „brațe teribile” sau „brațe monstruoase”) este un gen dispărut de mare dinozaur ornitomimosaur care a trăit în Cretacicul superior , în urmă cu aproximativ 71-69 milioane de ani ( Maastrichtian ), în ceea ce este acum Formația actuală Nemegt , Mongolia . Genul Deinocheirus conține o singură specie , și anume D. mirificus . Numele animalului derivă din primele fosile, găsite în 1965, care includeau o pereche de membre anterioare cu mâini și omoplați. În 1970, aceste fosile au devenit holotipul singurei specii atribuite genului Deinocheirus : D. mirificus ; numele, din vechea greacă , înseamnă literalmente „mână oribilă”, referindu-se la dimensiunea gigantică a brațelor animalului. În următorii 50 de ani, nu au fost găsite alte rămășițe fosile care pot fi atribuite genului, iar anatomia animalului a rămas un mister de ani de zile, până când în 2014 au fost găsite încă două exemplare complete care aruncă o nouă lumină asupra anatomiei animalului.
Deinocheirusul era un ornitomimosaur destul de neobișnuit: era cel mai mare reprezentant al cladei sale la 11 metri lungime și o greutate de 6,36 tone. Deși era un animal impunător, oasele sale goale, asemănătoare cu cele ale păsărilor, ușurau greutatea animalului. Brațele erau printre cei mai lungi dinozauri teropodi bipedali , măsurând singuri 2,4 metri (7,9 picioare) singuri și aveau și gheare mari. Picioarele, pe de altă parte, erau relativ scurte și aveau trei degete cu gheare contondente. Una dintre caracteristicile sale particulare era o extensie curioasă a vertebrelor neuronale , care formau un fel de pânză sau cocoașă, similară cu cea a cămilelor și dromedarilor de astăzi. Coada s-a terminat într-un pigostil (vertebre fuzionate la capătul cozii, asemănătoare cu cele găsite la păsări), sugerând posibila prezență a unui ventilator de pene la capătul cozii. Craniul , pe de altă parte, era destul de diferit de cel al altor ornitomimozauri, măsurând aproximativ 1.024 metri (3.36 ft) în lungime, cu o anatomie similară cu craniul unui hadrosaur .
Clasificarea Deinocheirusului a fost mult timp dezbătută și incertă: inițial animalul a fost interpretat ca un teropod carnivor al grupului carnosaurian , totuși asemănările cu familia ornitomimozaurilor au fost imediat observate de paleontologi. Ca rezultat, Deinocheirus a fost clasificat ca un ornithomimosaurus primitiv, în strânsă legătură cu genurile mai mici Garudimimus și Beishanlong , cu care a fost grupate în noua familie de Deinocheiridae . Spre deosebire de alți ornitomimozauri, membrii acestei familii nu sunt potriviți pentru a alerga. După ce a fost reclasificat ca ornitomimosaur, Deinocheirus a fost considerat un animal omnivor . Cu toate acestea, dieta sa este încă un mister: forma capului sugerează o dietă erbivoră, dar solzii de pești au fost găsiți în stomacul unui specimen împreună cu gastrolitii , ceea ce întărește ideea că este omnivor și că el a folosit probabil gastrolitii ca mijloace de măcinare a mâncării în stomac. Prin urmare, ghearele mari ale mâinilor ar fi putut fi folosite pentru a săpa fundurile râurilor și a colecta plante [1] . Cu toate acestea, arsenalul său de arme și dimensiunea lui nu l - au făcut invulnerabil, de fapt, urme de mușcături au fost descoperite pe oasele unui specimen de Deinocheirus care pot fi atribuite contemporane tyrannosaurid tarbozaur .
Descriere

Deinocheirusul este cel mai mare ornitomimosaur descoperit vreodată și unul dintre cele mai mari teropode cunoscute și unul dintre cele mai înalte; cel mai mare exemplar cunoscut măsoară până la 11 metri (36 ft) în lungime, cu o greutate estimată la 6,36 tone. Celelalte două exemplare sunt mai mici, holotipul fiind cu aproximativ 6% mai mic decât exemplarul mai mare, iar cel mai mic, un sub-adult, cu 74%. [2] Când erau cunoscute doar brațele animalului, au fost propuse diferite dimensiuni, obținute din diferite metode de calcul. Un studiu din 2010 a estimat înălțimea șoldului Deinocheirusului între 3,3 și 3,6 metri (11-12 ft). [3] În schimb, greutatea fusese deja estimată de la 2 (4.400 lb) la 12 tone (26.000 lb). Aceste dimensiuni enorme au fost făcute luând ca model tiranosauridele , o familie de dinozauri cu diferite construcții și cu brațe minuscule, proporțional cu dimensiunea corpului lor. [4] [5] [6]

Atât Deinocheirus atât therizinosauride therizinosaurus poseda cele mai lungi dintre toate membrele anterioare theropods cunoscute. [3] Membrele anterioare ale exemplarului holotipic măsoară numai 2,4 metri (7,9 picioare); humerusul (osul brațului superior) are aproximativ 93,8 cm (36,9 in) lungime, ulna aproximativ 68,8 cm (27,1 in), în timp ce mâna întreagă a animalului era lungă în jur de 77 cm (30 in), cu 19,6 cm în plus (7,7 in) ) de gheare curbate. Fiecare scapulocoracoid al centurii de umăr are o lungime de 1,53 metri (5,0 ft). Fiecare jumătate a ceratobranchialiei împerecheate măsoară aproximativ 42 cm (17 in). Omoplatul este lung și îngust, iar stema deltopectorală este pronunțată și triunghiulară. Humerusul era relativ subțire și puțin mai lung decât mâna. Ulna și raza au fost alungite și nu sunt strâns legate între ele într-o sindesmoză . Chiftelul era destul de alungit decât degetele de la picioare. Cele trei degete au aproximativ o lungime egală, dintre care prima este cea mai puternică și a doua este cea mai lungă. Mai multe zone aspre și impresii pe membrele anterioare indică prezența mușchilor mari. Majoritatea suprafețelor articulare ale oaselor brațului erau brazdate adânc, indicând faptul că animalul avea plăci groase de cartilaj între articulații . Deși brațele Deinocheirusului erau foarte dezvoltate, relația lor cu restul corpului era inferioară celei a ornitomimosaurului Ornithomimus , [7] în timp ce erau similare în proporție cu cele ale micului teropod Compsognathus . [8] Forcula (sau osul dorit), un element necunoscut la ornitomimozauri, avea formă de U. Membrele posterioare erau relativ scurte, iar osul coapsei ( femurul ) este mai lung decât tibia , așa cum este obișnuit. animale. Metatarsianul era mai scurt și prezenta metatarsul arcului, ca în majoritatea celorlalte teropode. Ghearele picioarelor erau ascuțite și cu vârf larg, mai degrabă decât conice ca cele ale majorității teropodelor. Proporțiile oaselor picioarelor erau foarte asemănătoare cu cele ale tiranozaurilor , datorită greutății mari pe care trebuiau să o susțină. [2]
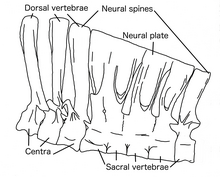
În ciuda faptului că Deinocheirusul este un animal gigantic și voluminos, coastele sale dorsale erau înalte și relativ simple, indicând faptul că corpul era relativ îngust. [9] Cele zece vertebre ale gâtului erau joase și lungi, devenind din ce în ce mai scurte, îndreptându-se spre craniu, oferind gâtului curbura clasică în formă de S tipică ornitomimozaurilor, datorită proporțiilor craniului. Spinele neuronale ale celor douăsprezece vertebre posterioare au crescut din ce în ce mai mult, ultima dintre acestea fiind de aproximativ 8,5 ori înălțimea părții centrale. Acest raport este același cu cel al coloanelor neuronale prezente în giganticul teropod Spinosaurus . Spinele neuronale aveau un sistem de ligamente , care probabil erau folosite pentru a susține abdomenul prin conectarea la șolduri și picioarele din spate. [2] În general, coloanele neuronale formau un fel de „pânză” sau o cocoașă grasă, care mergea de la mijlocul spatelui până la baza cozii, oarecum asemănătoare cu cele găsite în Spinosaurus . [10]
Toate vertebrele au fost în mare parte pneumatizate de sacii de aer, cu excepția osului atlas și a vertebrelor din spate ale cozii și au fost conectate la sistemul respirator. Vertebrele din spate au fost pneumatizate ca cele ale sauropodelor , cu un sistem extins de depresiuni. Aceste adaptări pot fi legate de gigantism , deoarece reduc greutatea animalului. Cele șase vertebre ale sacrului erau înalte și unite împreună atât la bază, cât și la vârful extensiilor osoase (vezi imaginea de mai sus din stânga), formând o placă neuronală compactă. Ileonul a fost , de asemenea , parțial pneomatized lângă vertebre sacrale. O parte a bazinului este hipertrofică , în comparație cu alți ornitomimozauri, pentru a susține greutatea animalului cu inserții musculare puternice. Oasele anterioare ale șoldului au fost înclinate în sus în talie. Vârful cozii Deinocheirusului s-a încheiat cu cel puțin două vertebre topite, care împreună au format un pigostil, similar cu cel găsit în alți dinozauri teropodi, cum ar fi oviraptorosaurii și terizinosauroizii . Mai mult, se crede că ornitomimozaurii posedă pene penacee , astfel încât această caracteristică ar susține ipoteza că animalul din viață ar avea un ventilator de pene la capătul cozii. [2]

Singurul craniu cunoscut, aparținând celui mai mare exemplar și măsurat la aproximativ 1.024 metri (3.36 ft) de la premaxilă, în partea anterioară, până în partea posterioară a condilului occipital . Cea mai lată parte a craniului se afla în spatele ochilor și avea o lățime de numai 23 cm (9,1 in). În anumite privințe, craniul era similar cu cel al altor ornitomimozauri, datorită formei sale înguste și joase, dar diferea prin faptul că botul era mai alungit. Pereții osoși ai craniului erau destul de subțiri, aproximativ 6 mm (0,24 in). La capătul botului avea un fel de cioc aplatizat și rotunjit, care la talie trebuie să fi fost acoperit cu keratină . Nările erau îndreptate în sus, iar osul nazal era o curea îngustă care se întindea peste orificiile ochiului. Diametrul exterior al inelelor sclerale a fost mic, de aproximativ 8,4 cm (3,3 in), relativ la dimensiunea craniului. Cea mai mică fereastră temporală a fost aproape de deschiderea ochiului, care a fost parțial închisă de osul jugal, similar cu Gallimimus . Fălcile erau fără dinți, maxilarul era înalt și masiv, în timp ce maxilarul era destul de alungit și subțiat și ușor curbat în jos. Proporțiile maxilarului-mandibulei au fost mai asemănătoare cu cele ale tiranozauridelor decât cele ale ornitomimozaurilor . Botul a fost în formă de spatulă (evazat în exterior pe laturi) și lățimea de aproximativ 25 cm (9,8 in), mai lată decât lățimea craniului. [2] Anatomia era similară cu craniul hadrosaurilor . [10]
Clasificare


Inițial, când Deinocheirus era cunoscut pentru membrele anterioare, clasificarea sa era destul de dificil de determinat și au fost propuse mai multe ipoteze. [10] Osmólska și Roniewicz au emis inițial ipoteza că Deinocheirusul nu aparținea niciunei familii teropode deja cunoscute, așa că au creat o nouă familie monospecifică , Deinocheiridae , plasată în infraordinea Carnosauria . Acest lucru s-a datorat oaselor mari și groase ale membrelor anterioare, deși cei doi paleontologi au găsit similitudini cu Ornithomimus și, într-o măsură mai mică, cu Allosaurus . [7] În 1971, John Ostrom a fost primul care a emis ipoteza că Deinocheirusul face parte din ornitomimosaurii, observând că animalul avea caracteristici similare și nu cu ornitomimozaurii. [2] [10] În 1976, Rhinchen Barsbold a numit ordinul Deinocheirosauria , care a inclus genurile aparent legate Deinocheirus și Therizinosaurus . O relație între Deinocheirus și terizinozauri , dotată și cu brațe lungi, a fost susținută de unii scriitori de mai târziu, dar astăzi această ipoteză a fost complet exclusă. [11]
În 2004, Peter Makovicky, Kobayashi și Currie au afirmat că Deinocheirus era probabil un ornitomimosaur primitiv, deoarece nu avea unele dintre caracteristicile tipice ale familiei Ornithomimidae . [12] Caracteristicile primitive includ ghearele sale curbate, raportul scăzut al omerului-omoplatului și lipsa unei sindesmoze. Un studiu realizat în 2006 de Kobayashi și Barsbold a constatat că Deinocheirus este probabil cel mai primitiv ornitomimosaur, deși nu au reușit să-și rezolve afinitățile în continuare, din cauza lipsei de elemente ale craniului și membrelor posterioare. [11] O cladistic analiză care însoțește 2014 descrierea celor două specimene mult mai complete a constatat că Deinocheirus a format un clade cu Garudimimus și Beishanlong , care au fost apoi incluse în Deinocheiridae.
Următoarea cladogramă arată poziția Deinocheirusului în Ornitomimosaurie : [2]
| Ornitomimosaurie |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Studiul din 2014 a definit Deinocheiridae ca o cladă care include „toți taxonii cu un strămoș comun mai recent cu Deinocheirus mirificus decât cu Ornithomimus velox ”. Cei trei membri împărtășesc diverse caracteristici anatomice la nivelul membrelor. Cladograma din 2014 a sugerat că ornitomimozaurii s-au împărțit în două linii principale în timpul Cretacicului inferior; Deinocheiridae și Ornithomimidae. Spre deosebire de alți ornitomimozauri, deinocheiridele nu sunt specializate în alergare. Particularitățile anatomice ale Deinocheirusului în comparație cu alți ornitomimozauri mult mai mici, pot fi explicate pe larg prin dimensiunea și greutatea sa mare. [2] Deinocheiridele și ornitomimidele mai mici nu aveau dinți, spre deosebire de ornitomimozaurii primitivi. [10]
Istoria descoperirii

Primele rămășițe fosile cunoscute ale Deinocheirusului au fost descoperite de paleontologul polonez Zofia Kielan-Jaworowska la 9 iulie 1965, pe situl Altan Ula III ( coordonate : 43 ° 33.987'N 100 ° 28.959'E) în bazinul Nemegt din deșertul Gobi. . Zofia a făcut parte dintr-o expediție paleontologică poloneză-mongolă, însoțită de paleontologul mongol Rinchen Barsbold , în perioada 1963-1965, organizată de Academia de Științe din Polonia și Academia de Științe din Mongolia. Expediția a durat trei zile, în perioada 9-11 iulie, pentru a excava exemplarul și a-l încărca pe un vehicul. Un raport din 1968 al lui Kielan-Jaworowska și Naydin Dovchin, care rezuma rezultatele expedițiilor, anunța că rămășițele reprezentau o nouă familie de dinozauri teropodi. [2] [13] Specimenul a fost descoperit într-un mic deal de gresie și constă dintr-un schelet parțial disjunct, majoritatea probabil erodat la descoperire. Specimenul consta din ambele membre anterioare, excluzând ghearele mâinii drepte, întregul brâu al umărului, centrul a trei vertebre dorsale, cinci coaste, gastralia (coaste abdominale) și două ceratobranșiale. Specimenul a fost făcut holotip al Deinocheirus mirificus , numit de Halszka Osmólska și Ewa Roniewicz în 1970. Denumirea generică, Deinocheirus provine din vechiul grec deinos (δεινός), care înseamnă „teribil” și cheir (χείρ), care înseamnă „mână” ", datorită dimensiunii și ghearelor puternice ale membrelor anterioare. Numele specific, mirificus , provine din latinesc și înseamnă „neobișnuit” sau „minunat”, ales pentru structura neobișnuită a membrelor anterioare. [7] Expedițiile polonezo-mongole s-au remarcat prin faptul că au fost conduse de femei, printre primele care au numit noi dinozauri. [14] Numărul de catalog al specimenului original de holotip a fost ZPal MgD-I / 6, dar de atunci a fost recatalogat ca MPC-D 100/18. [2]

Raritatea cunoscuților Deinocheirus rămâne inhibată pentru o înțelegere aprofundată a animalului timp de aproape jumătate de secol încoace, iar literatura științifică l-a descris adesea drept unul dintre cei mai „enigmatici”, „misterioși” și „bizari” dinozauri descoperiți vreodată. [2] [11] [15] Brațele holotipice au continuat să facă parte dintr-o expoziție itinerantă de fosile de dinozauri mongoli, vizitând diferite țări. [16] În 2012, Phil R. Bell, Philip J. Currie și Yuong-Nam Lee au anunțat descoperirea altor elemente ale eșantionului de holotip, inclusiv fragmente de gastralia, găsite de o echipă coreeană-mongolă care a repoziționat cariera originală. 2008. Semnele de mușcătură găsite pe cele două gastralii au fost atribuite unui Tarbosaurus . Acest lucru ar putea explica de ce specimenul Deinocheirus a fost găsit într-o stare împrăștiată și neasociată [15]
În 2013, descoperirea a două noi probe de Deinocheirus a fost anunțată înaintea conferinței anuale Society of Vertebrate Paleontology (SVP) de către Lee, Barsbold, Currie și colegii săi. Găzduite la Academia Mongolă de Științe, aceste exemplare fără scule au fost catalogate ca MPC-D 100/127 și MPC-D 100/128. MPC-D 100/128, este un exemplar sub-adult, găsit de oamenii de știință din Altan Ula IV (coordonate: 43 ° 36.091'N 100 ° 27.066'E) al Formației Nemegt în timpul Expediției Internaționale Dinosaice Coreea-Mongolia din 2006, dar a fost deja afectată de comercianții ilegali de fosile. Al doilea specimen, MPC-D 100/127, a fost găsit de oamenii de știință în Bugiin Tsav (coordonate: 43 ° 54.025'N 99 ° 58.359'E) în 2009. Este puțin mai mare decât holotipul și ar putea fi identificat în mod clar ca Deinocheirus de la labă din față stângă și, astfel, a ajutat la identificarea specimenului colectat anterior ca Deinocheirus . Acest specimen a fost recuperat și de dealerii de fosile, care au scos craniul, mâinile și picioarele, dar au lăsat un singur deget de la picioare în urmă. Probabil că fusese jefuit după 2002, pe baza banilor rămași în carieră. [2] [9] Craniile, ghearele și dinții sunt adesea vizate selectiv de braconieri în detrimentul restului scheletului (care este adesea vandalizat), deoarece au o piață mai mare. [17] Într-un interviu, Currie a declarat că politica echipei lor era să investigheze carierele după ce au fost jefuite și să recupereze ceva semnificativ și că găsirea de noi fosile de Deinocheirus a fost un motiv de bucurie, în ciuda oaselor vandalizate. Un model virtual de Deinocheirus a fost prezentat la SVP din acel an, provocând aplauze și complimente din partea mulțimii de paleontologi prezenți acolo. Paleontologul scoțian Stephen L. Brusatte a declarat că nu a fost niciodată atât de surprins de un discurs SVP, deși la conferință sunt prezentate în mod regulat noi fosile. [18]

După ce au fost anunțate noile specimene, s-a zvonit că un craniu jefuit și-a găsit drumul către un muzeu european prin intermediul pieței negre . [18] Elementele jefuite au fost observate într-o colecție privată europeană de către comerciantul francez de fosile François Escuillé, care a vorbit despre acesta din urmă paleontologului belgian Pascal Godefroit în 2011. Au suspectat că rămășițele aparțin Deinocheirusului , au contactat mongol-coreeanul echipa. Ulterior, Escuillé a achiziționat fosilele și le-a donat Institutului Regal Belgian de Științe ale Naturii. [19] Materialul recuperat a constat dintr-un craniu, o mână stângă și picioare, care fuseseră colectate în Mongolia, vândute unui cumpărător japonez și revândute la o petrecere germană. Echipa a ajuns la concluzia că aceste elemente aparțin specimenului MPC-D 100/127, întrucât singurul os restant al piciorului se încadrează perfect în materialul sechestrat, osul și matricea se potrivesc în culoare și, deoarece elementele aparțineau unui individ de aceleași dimensiuni , fără suprapunerea elementelor scheletice. [2] La 1 mai 2014, fosilele au fost repatriate în Mongolia de către o delegație de la Muzeul Belgian, în cadrul unei ceremonii desfășurate la Academia Mongolă de Științe. [20] Scheletul asamblat a fost depus la Muzeul Central al Dinozaurilor Mongolilor din Ulan Bator , împreună cu un schelet Tarbosaurus , adus și înapoi în Mongolia după exportul ilegal. Paleontologul american Thomas R. Holtz a declarat într-un interviu că noul Deinocheirus rămâne similar cu „produsul unei relații amoroase secrete între un hadrosaur și un Gallimimus ”. [19]
Prin combinarea elementelor matricei, ambele specimene noi reprezintă aproape întregul schelet al Deinocheirusului , deoarece MPC-D 100/127 include tot materialul în afară de vertebrele dorsale centrale, majoritatea vertebrelor caudale și membrele anterioare drepte; MPC-D 100/128 umple majoritatea golurilor din celălalt schelet, cu aproape toate vertebrele dorsale și caudale, ileonul, un ischiu parțial și majoritatea membrelor posterioare stângi. În 2014, exemplarele au fost descrise în revista Nature de Lee, Barsbold, Currie, Yoshitsugu Kobayashi, Hang-Jae Lee Lee, Godefroit, Escuillié și Tsogtbaatar Chinzorig. [2] În mod curios, o serie similară de evenimente a fost raportată la începutul anului 2014 cu Spinosaurus, un alt teropod navigat, cunoscut doar din câteva rămășițe din 1912. Rămășițele matricei au fost reunite cu exemplarele obținute de oamenii de știință, iar Spinosaurus s- a dovedit a fi foarte diferit de alte spinosauride . Cele două cazuri au arătat că stilul de viață și aspectul animalelor dispărute necunoscute nu pot fi întotdeauna deduse în siguranță de la rudele apropiate. [10] Până în 2017, guvernul mongol și-a intensificat eforturile de a captura fosile matriciale de la colecționari și de a le repatria, deoarece dovedirea provenienței lor a devenit o preocupare științifică și politică. Prin urmare, un studiu a testat posibilitatea identificării fosilelor matriciale prin metode geochimice , folosind Deinocheirus și alți dinozauri Nemegt ca exemple. [17]
Paleobiologie
Scurtele ghearele tocite ale Deinocheirus sunt foarte similare cu cele ale therizinosaurus Alxasaurus , indicând faptul că brațele lungi ale animalului și gheare mari au fost folosite în primul rând pentru săpat și recoltarea plantelor. Mai mult, ghearele contondente ale picioarelor ar fi permis animalului să nu se scufunde în pământul moale și noroios. Membrele posterioare solide și regiunea caracteristică a șoldului indică faptul că animalul s-a mișcat destul de încet, spre deosebire de alți ornitomimozauri. Cu toate acestea, dimensiunea sa impresionantă a fost un factor de descurajare excelent împotriva prădătorilor, cum ar fi Tarbosaurus . Atât cocoașa dorsală, cât și posibilul ventilator al penelor cozii au fost probabil folosite ca afișaj sexual. Deinocheirusul era probabil un animal diurn (activ în timpul zilei), deoarece inelele sclerotice ale ochilor erau relativ mici comparativ cu lungimea craniului. [2] Mâinile aveau o mobilitate bună față de brațul inferior, dar erau capabile de o mișcare limitată de flexie, incapabilă să se închidă în apucare. [7]
Creierul Deinocheirus a fost reconstruit printr-o scanare CT și prezentat la conferința din 2014 a Society of Vertebrate Paleontology. Creierul animalului era globular și avea o formă similară cu cea a păsărilor și teropodelor troodontide , telencefalul a fost extins similar cu majoritatea teropodelor, iar bulbii olfactivi erau relativ mari. Creierul a fost proporțional mic și compact și coeficientul său de encefalizare reptiliană ( raport creier-corp) este estimat la 0,69, ceea ce este destul de scăzut pentru teropode și similar cu cel al sauropodelor. Alți ornitomimozauri au creiere proporțional mai mari, astfel încât creierul mic al Deinocheirusului reflectă comportamentul său social și dieta. Coordonarea și echilibrul său nu erau la fel de importante ca cele ale teropodelor carnivore. [21] Un studiu asupra microstructurii osoase prezentat la cea de-a 13-a reuniune anuală a Asociației Europene a Paleontologilor de Vertebrate din 2015 a arătat că Deinocheirusul avea probabil o rată metabolică ridicată și a crescut rapid înainte de a atinge maturitatea sexuală . [22]
Dietă

Forma specială a craniului Deinocheirus arată că acesta din urmă avea o dietă mai specializată decât alți ornitomimozauri. Ciocul a fost similar cu cel al rațelor , indicând faptul că animalul s-ar fi putut hrăni cu apă sau s-ar fi hrănit cu vegetație terestră, cum ar fi unii sauropodi și hadrosauri . Punctele de atașare pentru mușchii care deschid și închid maxilarele erau foarte mici în comparație cu dimensiunea craniului, indicând faptul că Deinocheirusul avea o mușcătură destul de slabă. Prin urmare, craniul a fost probabil conceput pentru a tăia fibrele fragede ale plantelor acvatice. Adâncimea maxilarului inferior indică prezența unei limbi mari, care ar fi putut ajuta animalul să înghită mâncarea colectată de ciocul mare atunci când animalul se hrănea în corpurile de apă dulce. [2]
S-au găsit peste 1.400 de gastroliti (calculi stomacali, de la 8 la 87 milimetri) între coaste și gastralia specimenului MPC-D100 / 127. Raportul dintre masa gastrolitului și greutatea totală, 0,0022, susține teoria conform căreia aceste gastrolite au ajutat animalele fără dinți să-și macine hrana. Caracteristici precum prezența unui cioc și a maxilarului în formă de U inversat indică adesea un erbivor facultativ printre teropodele coelurosaurilor. În ciuda acestor caracteristici, vertebrele și solzii de pești au fost de asemenea găsiți în rândul gastrolitilor, sugerând că Deinocherus a fost omnivor . [2] Se crede că ornitomimozaurii au mâncat atât plante, cât și animale mici. [10]

Înainte de descoperirea unor schelete mai complete, paleontologii au emis ipoteza diferitelor comportamente și diete pentru Deinocheirus . Studiile timpurii au indicat, în general, acest animal ca fiind un carnivor, similar cu Allosaurus , care și-a folosit picioarele din față pentru a înghiți prada. [10] În descrierea lor originală, Osmólska și Roniewicz au descoperit că mâinile Deinocheirusului nu erau potrivite pentru apucare, dar puteau fi folosite în schimb pentru a-și tăia prada. [7] În 1970, paleontologul rus Anatoly Konstantinovich Rozhdestvensky a comparat membrele anterioare ale Deinocheirusului cu cele ale leneșilor , determinându-l să facă ipoteza că Deinocheirusul era un dinozaur arbore , care se hrănea cu plante și poate cu ouă sau cu orice animal mic pe care îl găsea între ramuri. . [23] În 1988, Gregory S. Paul a sugerat, în schimb, că ghearele erau prea clare pentru scopuri de pradă, dar ar fi o bună armă defensivă. [6] Nel tentativo di determinare le nicchie ecologiche per Deinocheirus e Therizinosaurus nel 2010, Phil Senter e James H. Robins conclusero che il Deinocheirus aveva il più ampio spettro di movimenti verticali di ogni altro erbivoro della Formazione Nemegt, a causa dell'altezza al bacino, e che fosse specializzato nel nutrirsi di fogliame alto. [3] Nel 2017, è stato suggerito che gli artigli del Deinocheirus si fossero adattati per estrarre grandi quantità di piante erbacee dall'acqua e per diminuire la resistenza con l'acqua. [24]
Paleopatologia
Osmólska e Roniewicz hanno riferito che il campione olotipico presenta fori, scanalature e tubercoli anormali sulla prima e seconda falange del secondo dito sinistro che potrebbero essere il risultato di ferite all'articolazione tra le due ossa. Il danno potrebbe aver causato cambiamenti nella disposizione dei legamenti dei muscoli. Anche i due coracoidi sono sviluppati diversamente. [7][25] Una costola del campione MPC-D 100/127 mostra un trauma guarito che ha rimodellato l'osso. [2] Nel 2012 sono stati riportati segni di morsi su due gastralie del campione olotipo. Le dimensioni e la forma dei segni del morso corrispondono ai denti del Tarbosaurus , il più grande predatore conosciuto della Formazione Nemegt. Sono stati identificati vari tipi di tracce di alimentazione; forature, sgorbie, striature, denti frammentari e combinazioni dei segni precedenti. I segni del morso rappresentano probabilmente un comportamento alimentare invece di un'aggressione tra specie, e il fatto che i segni dei morsi non siano stati trovati altrove sul corpo indica che il predatore si è concentrato sugli organi interni. I segni del morso di Tarbosaurus sono stati ritrovati anche nei fossili di un hadrosauro e un sauropode, ma i segni di morsi di teropodi su ossa di altri teropodi sono molto rari nella documentazione fossile. [15]
Paleocologia

I tre campioni noti di Deinocheirus sono stati recuperati dalla Formazione Nemegt , nel deserto del Gobi, nella Mongolia meridionale. Questa formazione geologica non è mai stata datata radiometricamente , ma la fauna presente nei reperti fossili indica che si fu probabilmente depositata durante la fase iniziale del Maastrichtiano , alla fine del Cretaceo superiore , circa 70 milioni di anni fa. [26] [27] [28] Le facies della roccia della formazione suggeriscono la presenza di corsi d'acqua e canali fluviali, distese fangose e laghi poco profondi. Tali grandi canali fluviali e depositi di sedimenti sono la prova di un clima molto più umido di quelli che si trovano nelle più vecchie formazioni di Barun Goyot e Djadochta. Tuttavia, i depositi di caliche indicano che si verificavano periodi di siccità periodici. I sedimenti furono depositati nei canali e nelle piane alluvionali dai fiumi. [29]

Si ritiene che il Deinocheirus fosse una presenza piuttosto comune all'interno della Formazione Nemegt, in quanto gli unici tre esemplari ritrovati sono distanti tra loro 50 km (31 miglia). I sistemi fluviali della Formazione Nemegt fornivano un'adeguata nicchia per il Deinocheirus per le sue abitudini onnivore. [2] L'ambiente era simile a quello dell'odierno Delta dell'Okavango , della Botswana . All'interno di questo ecosistema, il Deinocheirus si sarebbe nutrito sia di piante sia di piccoli animali, incluso il pesce. Il Deinocheirus competeva inoltre con altri grandi dinosauri erbivori per nutrirsi sulle cime degli alberi, come il teropode Therizinosaurus dal collo lungo, vari sauropodi titanosauri e l' hadrosauro Saurolophus . Tuttavia, grazie alla posizione elevata dell'anca, i lunghi arti ed il collo lungo, il Deinocheirus poteva raggiungere anche i fogliami più alti. Insieme al Deinocheirus , la scoperta del Therizinosaurus e del Gigantoraptor mostrano come tre gruppi di teropodi erbivori (ornitomimosauri, therizinosauri e oviraptorosauri) raggiunsero indipendentemente le loro dimensioni massime nel Cretaceo superiore dell'Asia. [10]
Gli habitat intorno ai fiumi del Nemegt dove viveva il Deinocheirus costituivano una casa per una vasta gamma di organismi. Si trovano occasionalmente fossili di molluschi e una gran varietà di altri animali acquatici come pesci e tartarughe. [26] I crocodylomorfi del Nemegt includevano diverse specie di Shamosuchus . [30] I mammiferi fossili sono rari nella formazione nemegt, ma sono stati ritrovati molti uccelli, tra cui l' enantiornithine Gurilynia , gli hesperornithiformi Judinornis e Brodavis , e il possibile anseriforme Teviornis . I dinosauri erbivori della formazione Nemegt includono gli ankylosauri Tarchia e Saichania ; il pachycephalosouro Prenocephale ; gli hadrosauridi Saurolophus e Barsboldia ed il sauropode Nemegtosaurus . [26] [31] Tra i teropodi predatori che vissero insieme al Deinocheirus si trovano i tirannosauroidi Tarbosaurus , Alioramus e Bagaraatan ei troodontidi Borogovia , Tochisaurus e Saurornithoides . Altri gruppi di teropodi onnivori e/o erbivori includono il therizinosauro Therizinosaurus ; gli oviraptorosauri Elmisaurus , Nemegtomaia e Rinchenia ; e altri ornitomimosauri Anserimimus e Gallimimus . [32]
Note
- ^ Deinocheirus Mirificus, un dinosauro misterioso - da http://www.dinosauro.eu/notizie-dinosauri/deinocheirus-mirificus-il-dinosauro-di-star-wars
- ^ a b c d e f g h i j k l m n o p q r s YN Lee, R. Barsbold , PJ Currie , Y. Kobayashi, HJ Lee, P. Godefroit , FO Escuillié e T. Chinzorig, Resolving the long-standing enigmas of a giant ornithomimosaur Deinocheirus mirificus , in Nature , vol. 515, n. 7526, 2014, pp. 257–260, DOI : 10.1038/nature13874 .
- ^ a b c P. Senter e JH Robins, Hip heights of the gigantic theropod dinosaurs Deinocheirus mirificus and Therizinosaurus cheloniformis , and implications for museum mounting and paleoecology ( PDF ), in Bulletin of the Gunma Museum of Natural History , n. 14, 2010, pp. 1–10.
- ^ GS Paul , The Princeton Field Guide to Dinosaurs , Princeton University Press, 2010, p. 112, ISBN 978-0-691-13720-9 . URL consultato il 30 aprile 2019 (archiviato dall' url originale l'8 gennaio 2017) .
- ^ BV Valkenburgh e RE Molnar, Dinosaurian and mammalian predators compared , in Paleobiology , vol. 28, n. 4, 2002, pp. 530–540, DOI : 10.1666/0094-8373(2002)028<0527:dampc>2.0.co;2 , JSTOR 3595499 .
- ^ a b GS Paul , Predatory Dinosaurs of the World , Simon & Schuster, 1988, pp. 228 , 382, ISBN 978-0-671-61946-6 .
- ^ a b c d e f H. Osmólska e E. Roniewicz, Deinocheiridae, a new family of theropod dinosaurs ( PDF ), in Palaeontologica Polonica , n. 21, 1970, pp. 5–19.
- ^ KM Middleton e SM Gatesy,Theropod forelimb design and evolution , in Zoological Journal of the Linnean Society , vol. 128, n. 2, 2000, pp. 160, 172, DOI : 10.1111/j.1096-3642.2000.tb00160.x .
- ^ a b YN Lee, R. Barsbold , PJ Currie , Y. Kobayashi e HJ Lee, New specimens of Deinocheirus mirificus from the Late Cretaceous of Mongolia ( PDF ), in Society of Vertebrate Paleontology Abstracts of Papers , 2013, p. 161 (archiviato dall' url originale il 4 dicembre 2014) .
- ^ a b c d e f g h i TR Holtz , Paleontology: Mystery of the horrible hands solved , in Nature , vol. 515, n. 7526, 2014, pp. 203–205, DOI : 10.1038/nature13930 , PMID 25337885 .
- ^ a b c Y. Kobayashi e R. Barsbold , Ornithomimids from the Nemegt Formation of Mongolia ( PDF ), in Journal of the Paleontological Society of Korea , vol. 22, n. 1, 2006, pp. 195–207.
- ^ PJ Makovicky, Y. Kobayashi e PJ Currie , Ornithomimosauria , in David B. Weishampel, Peter Dodson e Halszka Osmólska (a cura di), The Dinosauria , 2ª ed., University of California Press, 2004, pp. 137 –150, ISBN 978-0-520-24209-8 .
- ^ Z. Kielan-Jaworowska e N. Dovchin, Narrative of the Polish-Mongolian Palaeontological Expeditions 1963–1965 ( PDF ), in Palaeontologica Polonica , vol. 19, 1968, p. 24. URL consultato il 27 novembre 2017 (archiviato dall' url originale il 12 luglio 2020) .
- ^ P. Dodson, The Horned Dinosaurs: A Natural History , Princeton University Press, 1998, p. 9, ISBN 978-0-691-05900-6 .
- ^ a b c PR Bell, PJ Currie e YN Lee, Tyrannosaur feeding traces on Deinocheirus (Theropoda:?Ornithomimosauria) remains from the Nemegt Formation (Late Cretaceous), Mongolia , in Cretaceous Research , vol. 37, 2012, pp. 186–190, DOI : 10.1016/j.cretres.2012.03.018 .
- ^ Cretaceous Mongolian Dinosaurs , su dinocasts.com (archiviato dall' url originale il 2 febbraio 2015) .
- ^ a b F. Fanti, PR Bell, M. Tighe, LA Milan e E. Dinelli, Geochemical fingerprinting as a tool for repatriating poached dinosaur fossils in Mongolia: A case study for the Nemegt Locality, Gobi Desert , in Palaeogeography, Palaeoclimatology, Palaeoecology , 2017, DOI : 10.1016/j.palaeo.2017.10.032 .
- ^ a b B. Switek, Mystery Dinosaur Finally Gets a Body , in National Geographic Society , 4 novembre 2013.
- ^ a b J. Hecht, Stolen dinosaur head reveals weird hybrid species , 12 maggio 2014.
- ^ The "horrible hand" Deinocheirus dinosaur's fossils are repatriated to its home country , su infomongolia.com (archiviato dall' url originale il 12 maggio 2014) .
- ^ P. Lauters, YN Lee, R. Barsbold , PJ Currie , Y. Kobayashi, FO Escuillé e P. Godefroit , The brain of Deinocheirus mirificus , a gigantic ornithomimosaurian dinosaur from the Cretaceous of Mongolia ( PDF ), in Society of Vertebrate Paleontology Abstracts of Papers , 2014, p. 166 (archiviato dall' url originale il 4 dicembre 2014) .
- ^ M. Kundrát e YN Lee, First insights into the bone microstructure of Deinocheirus mirificus ( PDF ), in 13th Annual Meeting of the European Association of Vertebrate Palaeontologists , 2015, p. 25. URL consultato il 28 novembre 2017 (archiviato dall' url originale il 20 luglio 2015) .
- ^ ( RU ) AK Rozhdestvensky , Gigantskiye kogti zagadochnykh mezozoyskikh reptiliy [ Giant claws of enigmatic Mesozoic reptiles ], in Paleontologicheskii Zburnal , vol. 1970, n. 1, 1970, pp. 117–125.
- ^ T. Chinzorig, Y. Kobayashi, K. Tsogtbaatar, PJ Currie, R. Takasaki, T. Tanaka, M. Iijima e R. Barsbold, Ornithomimosaurs from the Nemegt Formation of Mongolia: manus morphological variation and diversity , in Palaeogeography, Palaeoclimatology, Palaeoecology , 2017, DOI : 10.1016/j.palaeo.2017.10.031 .
- ^ RE Molnar,Theropod Paleopathology: a Literature Survey , in Darren H. Tanke e Kenneth Carpenter (a cura di), Mesozoic Vertebrate Life , Indiana University Press, 2001, p. 343 , ISBN 978-0-253-33907-2 .
- ^ a b c T. Jerzykiewicz e DA Russell , Late Mesozoic stratigraphy and vertebrates of the Gobi Basin , in Cretaceous Research , vol. 12, n. 4, 1991, pp. 345–377, DOI : 10.1016/0195-6671(91)90015-5 .
- ^ RM Sullivan, A taxonomic review of the Pachycephalosauridae (Dinosauria: Ornithischia) ( PDF ), in Spencer G. Lucas e Robert M. Sullivan (a cura di), Late Cretaceous vertebrates from the Western Interior , vol. 35, New Mexico Museum of Natural History and Science Bulletin, 2006, pp. 347–366.
- ^ FM Gradstein, JG Ogg e AG Smith, A Geologic Time Scale 2004 , Cambridge University Press, 2005, pp. 344 –371, ISBN 978-0-521-78142-8 .
- ^ M. Novacek, Dinosaurs of the Flaming Cliffs , Anchor, 1996, p. 133 , ISBN 978-0-385-47775-8 .
- ^ ( RU ) MB Efimov, Peresmotr iskopayemykh krokodilov Mongolii [ Revision of the fossil crocodiles of Mongolia ], in Sovmestnaya Sovetsko-Mongol'skaya Paleontologicheskaya Ekspeditsiya Trudy , vol. 24, 1983, pp. 76–96.
- ^ JH Hurum e K. Sabath, Giant theropod dinosaurs from Asia and North America: Skulls of Tarbosaurus bataar and Tyrannosaurus rex compared ( PDF ), in Acta Palaeontologica Polonica , vol. 48, n. 2, 2003, p. 188.
- ^ TR Holtz , Tyrannosauroidea , in David B. Weishampel, Peter Dodson e Halszka Osmólska (a cura di), The Dinosauria , 2ª ed., University of California Press, 2004, p. 124 , ISBN 978-0-520-24209-8 .
Bibliografia
- Kielan-Jaworowska, Z. (1966). "Third (1965) Polish-Mongolian Palaeontological Expedition to the Gobi Desert and western Mongolia". Bulletin de l'Académie Polonaise des Sciences, C1. II 14 (4): 249–252.
- Osmólska, H. and Roniewicz, E. (1970). "Deinocheiridae, a new family of theropod dinosaurs." Palaeontologica Polonica, 21: 5-19.
- Rozhdestvensky, AK (1970). "Giant claws of enigmatic Mesozoic reptiles." Paleontological Journal, 1970(1): 117-125.
- Lambert, D. (1983). A Field Guide to Dinosaurs. New York: Avon Books.
- Paul, GS (1988). Predatory Dinosaurs of the World. New York: Simon & Schuster.
- Molnar, RE, 2001, Theropod paleopathology: a literature survey: In: Mesozoic Vertebrate Life, edited by Tanke, DH, and Carpenter, K., Indiana University Press, p. 337-363.
- Valkenburgh, B. and Molnar, RE (2002). "Dinosaurian and mammalian predators compared." Paleobiology, 28(4): 527–543.
- Makovicky, PJ, Kobayashi, Y., and Currie, PJ (2004). "Ornithomimosauria." In DB Weishampel, P. Dodson and H. Osmólska (eds.), The Dinosauria, Second Edition. University of California Press, Berkeley.
- Kobayashi, Y., and Barsbold, R. (2006). "Ornithomimids from the Nemegt Formation of Mongolia." Journal of the Paleontological Society of Korea, 22(1): 195-207.
- Paul, GS, 2010, The Princeton Field Guide to Dinosaurs, Princeton University Press p. 112
- Senter, P. and Robins, HJ (2010). "Hip heights of the gigantic theropod dinosaurs Deinocheirus mirificus and Therizinosaurus cheloniformis, and implications for museum mounting and paleoecology." Bulletin of the Gunma Museum of Natural History, 14: 1-10. [1]
- Bell, PR Currie, PJ, and Lee Y.-N. (2012). "Tyrannosaur feeding traces on Deinocheirus (Theropoda:?Ornithomimosauria) remains from the Nemegt Formation (Late Cretaceous), Mongolia." Cretaceous Research, available online 25 April 2012.
Altri progetti
-
 Wikimedia Commons contiene immagini o altri file su Deinocheirus mirificus
Wikimedia Commons contiene immagini o altri file su Deinocheirus mirificus -
 Wikispecies contiene informazioni su Deinocheirus mirificus
Wikispecies contiene informazioni su Deinocheirus mirificus
Collegamenti esterni
- ( EN ) Deinocheirus mirificus , su Fossilworks.org .
| Controllo di autorità | LCCN ( EN ) sh2007008539 |
|---|