Equus hemionus
| Fundul sălbatic asiatic | |
|---|---|
 | |
| Starea de conservare | |
 Aproape de amenințare (nt) [1] | |
| Clasificare științifică | |
| Domeniu | Eukaryota |
| Regatul | Animalia |
| Phylum | Chordata |
| Clasă | Mammalia |
| Ordin | Perissodactyla |
| Familie | Echidele |
| Tip | Equus |
| Specii | E. hemionus |
| Nomenclatura binominala | |
| Equus hemionus Pallas , 1775 | |
Mărul sălbatic asiatic ( Equus hemionus Pallas , 1775 ) este un reprezentant al familiei Equidae . În ciuda faptului că este asemănător exterior cu măgarul sălbatic african ( Equus africanus ) - strămoșul măgăriilor domestice -, are și multe caracteristici comune cu calul , atât de mult încât în limba germană este cunoscut și sub numele de Halbesel , „jumătate de fund”. , sau Pferdeesel , „Calul măgarului”. Este, de asemenea, cunoscut sub numeroase denumiri regionale, precum „khur” sau „kulan” (vezi Taxonomia ). Mâna sălbatică asiatică locuiește în regiuni aride, semi-deșertice și de stepă din vestul până în centrul și nordul Asiei și se hrănește în principal cu ierburi piele. Structura sa socială este considerată a fi foarte complexă, dar specia nu a fost încă studiată în detaliu. În general, se consideră că este în pericol de dispariție.
Descriere
Aspect

Cu o lungime a trunchiului capului de peste 200 cm, la care se adaugă o coadă lungă de 40 cm, o înălțime la greaban de 97-138 cm și o greutate de 200-260 kg, fundul sălbatic asiatic este puțin mai mare decât media măgar domestic . Dimorfismul sexual este doar ușor pronunțat. De obicei, este o creatură subțire, cu picioare lungi, fiecare se termină într-o copită largă mai mare decât cea a măgarului sălbatic african ( Equus africanus ). La fel ca în toate zebrele și măgarii, „castanele” caracteristice, mici plăci cornene maronii similare calusurilor, sunt prezente doar pe picioarele din față, spre deosebire de caii sălbatici, unde apar și pe picioarele din spate. Urechile sunt, de asemenea, mai lungi decât cele ale măgarilor adevărați; în plus, ciucura cozii și coama sunt mai puțin pronunțate, în timp ce zona albă caracteristică de la capătul botului tipică măgarilor este prezentă. Culoarea și lungimea hainei variază în funcție de anotimpuri. În haina scurtă de vară, părțile superioare ale corpului pot fi gri, de la galben pal la ocru sau roșu maroniu; partea inferioară, partea inferioară a gâtului și picioarele sunt de culoare albă sau crem. Subspeciile individuale diferă între ele prin intensitatea culorilor stratului. Tranziția culorilor pe părțile laterale ale corpului creează un model de semilună în toate formele, care se ridică parțial pe șolduri, dar diferă semnificativ de linia în formă de "M" tipică pentru haina cailor sălbatici, pe care, de obicei, o au culoare mai închisă pe picioare. Blana lungă de iarnă este în general mai închisă. Linia maro închis care se întinde de-a lungul coloanei vertebrale are o lățime de 60-80mm vara și 70-90mm iarna. Această dungă dorsală este adesea înconjurată de zone de culoare mai deschisă. Modelul cu dungi orizontale al picioarelor, tipic măgarului sălbatic african, este rar întâlnit la speciile asiatice [1] [2][3] .
Craniul și dinții

Craniul măgarului sălbatic asiatic măsoară între 42 și 52 cm și este foarte asemănător cu cel al kiangului ( Equus kiang ), cu botul scurt tipic. Linia frunții este relativ dreaptă, osul occipital este îngust și vertical și se extinde chiar dincolo de suprafețele unde este fixată prima vertebră cervicală (condilii). Osul nazal are o lungime de până la 22cm și are o ușoară curbă în față. Cavitatea nazală extinsă între oasele nazale și premaxilare și maxilarul superior este relativ mare. Oculația este uneori foarte adâncă în craniu, dar întotdeauna situată în spatele ultimului molar [2] [4] .
Mandibula masivă poate ajunge la 42 cm lungime, astfel încât osul maxilarului poate avea o lățime de până la 5 cm. În special, articulațiile maxilarului sunt foarte puternice. Formula dentară la exemplarele adulte este: . Incisivii sunt foarte largi și în maxilarul superior sunt vizibil mai înclinați înainte decât în kiang ( Equus kiang ); caninul este de obicei mic. Diastema dintre dentiția anterioară și cea posterioară poate avea o lățime de până la 8,6 cm. Premolarii au o structură similară cu molarii , adică sunt clar molarizați și, uneori, primul dinte molar este rudimentar (dintele de lup). În general, coroana molarilor are o mulțime de ciment și benzi de email răsucite. Pe suprafața de mestecat a molarilor inferiori, smalțul curge între cele două proeminențe numite metaconidă și metastilidă a zonei posterioare (latura linguală), parțial sub forma unui „V”, dar mai presus de toate și mai clar sub forma unui „U” [2] .

Sensuri și vocalizări
Ca și în alte ecvidee, comunicarea între măgarii sălbatici asiatici are loc în principal pe calea olfactivă , prin secreții excretate, dar aceste animale au și vedere și auz bine dezvoltate. Acestea fug de obicei deplasându-se la aproximativ 2 km de potențiali prădători [5] [6] . Până în prezent, nu a fost efectuat niciun studiu exhaustiv asupra vocalizărilor emise.
Distribuție și habitat
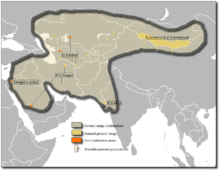
Gama nativă a măgăriei sălbatice asiatice s-a extins din regiunea mediteraneană a vestului Asiei , prin Transcaucasia și Asia Centrală , până în zonele vestice ale sudului Asiei și sudului Asiei de Nord . Astăzi această gamă este foarte fragmentată, atât de mult încât specia este acum limitată la anumite zone din Mongolia , Turkmenistan , Kazahstan , India , Iran , Ucraina și Arabia Saudită . Emione locuiește în principal în deșertul Gobi și khur în Rann of Kutch . Cele mai mari populații ale kulanului se găsesc în rezervația naturală Badkhyz și rezervația naturală Andassay, în timp ce onagerul este încă prezent în număr mare în parcul național Touran și în Bahram-e-Goor [1] .
Mâna sălbatică asiatică trăiește în medii aride: semi-deșert , stepe , stepe de munte și, în unele cazuri, deșerturi. Aceste habitate extreme, care pot ajunge la La 2 000 m deasupra nivelului mării, acestea se caracterizează prin temperaturi ridicate vara (până la 40 ° C) și temperaturi scăzute iarna (până la -35 ° C) și prin precipitații reduse, uneori sub 100 mm pe an. În plus, toate aceste zone sunt caracterizate de vegetație foarte rar [1] [7] . În partea de vest a arealului, măgarul sălbatic asiatic și-a împărțit habitatul cu măgarul sălbatic african, deși preferă zonele inferioare. Astăzi, însă, ambele specii sunt aici dispărute în natură [8] . La est, însă, raza sa de acțiune se suprapune parțial cu cea a calului lui Przewalski ( Equus ferus przewalskii ) [6] .
Biologie
Comportamentul teritorial

Structura socială a măgarilor sălbatici asiatici este foarte diversă. Subspecii nordice, precum hemionul deșertului Gobi și kulanul din Asia Centrală, tind să formeze turme formate dintr-un armăsar și mai multe femele cu mânzi și ocupă zone de acasă mari. Extinderea acestor zone, de la 4 500 a 40 000 km² , poate varia considerabil în funcție de anotimpuri, fiind mai puțin vara și mai mare iarna. În unele cazuri, se formează grupuri mari care pot include până la 450 de exemplare, dar acest lucru apare de obicei în zone deosebit de bogate în alimente sau la aburi. Deoarece aceste asociații se dizolvă rapid într-o zi, nu pare să existe nicio formă de ierarhie în ele. Există, de asemenea, „grupuri de burlaci” formate din masculi tineri care se formează adesea iarna. Astfel de grupuri sunt cunoscute și printre caii sălbatici și zebrele de stepă ( Equus quagga ) și de munte ( Equus zebra ) [6] [9] [10] . Cu toate acestea, ocazional, se pot forma și teritorii temporare, care sunt apoi apărate agresiv [11] .
Subspecii sudice, cum ar fi khur și onager, tind spre un stil de viață pur teritorial, cu teritorii care se suprapun parțial. Armăsarii dominanți ocupă teritorii care ating 9 km², dar pot fi, de asemenea, considerabil mai mari. Aceste teritorii conțin locuri de hrănire și odihnă și surse de apă permanente sau temporare. Cu toate acestea, jgheaburile sunt de obicei situate la marginea teritoriului și nu în centru. Traseele utilizate frecvent sunt marcate cu fecale și urină , folosind adesea aceleași puncte de marcare. Femelele cu mânzi se adună uneori în grupuri mici care pășunesc în zone de până la 20 km² suprapunându-se pe cele ale altor grupuri dominante și armăsari. Comportament similar a fost găsit la zebra lui Grévy ( Equus grevyi ) și la fundul sălbatic african [5] [12][13] .
Această diversitate a comportamentului social al diferitelor populații de fund sălbatic nu a fost încă pe deplin înțeleasă. Poate că factorii climatici sau presiunea de vânătoare de la prădători pot avea o anumită influență. În zona khur și a onagerilor, prădătorii mari sunt aproape absenți, astfel încât aceste animale au menținut stilul de viață original al unei specii teritoriale. Pe de altă parte, kulanii și emisiile sunt supuse atacurilor frecvente ale lupilor, care ar fi putut stimula formarea haitelor, deoarece grupurile mai stabile au șanse mai mari de supraviețuire. Mai mult, în aceste cazuri, armăsarul își apără activ turma [9] .
Dietă
Există doar câteva studii detaliate despre dieta măgarului sălbatic asiatic; în general, putem spune că molarii săi cu o coroană înaltă și caracterizați printr-o cantitate mare de ciment sunt potriviți pentru mestecarea substanțelor vegetale pe bază de siliciu ( pășunat ). Populațiile din Gobi preferă specii precum Stipa care se găsesc în număr mare în stepele în care trăiesc ( stepele Stipa ), dar consumă și buruieni , ierburi din genul Achnatherum , paie de mlaștină și stuf . În dieta măgarului sălbatic asiatic s-a înregistrat și prezența Artemisia , Anabasis , salsola , saxaul și caragana [9] [14] . Mai mult, în unele regiuni, prazul , Zygophyllum și tamariscurile ocupă o parte substanțială a dietei [15] . Meiul indian , sporobolele , eragrostidele și ierburile din genul Dicanthium sunt, de asemenea, incluse în dieta celor mai sudice populații [16] . Cu toate acestea, în timpul sezonului uscat, când ierburile se ofilesc, măgarul sălbatic își poate îndrepta atenția și asupra plantelor lemnoase, folosindu-și copitele pentru a rupe stratul de lemn pentru a ajunge în zonele cele mai bogate în lichide. Exemplarele au fost observate, de asemenea, hrănindu-se cu păstăi încărcate cu semințe [1] .
Datorită condițiilor extrem de aride ale zonelor în care trăiește, măgarul sălbatic are nevoie de surse de apă disponibile în mod constant, care nu ar trebui să se afle la mai mult de 10-15 km distanță - chiar dacă mișcări lungi de până la 30 km spre punctele de apă. În căutarea apei, măgarul sălbatic poate săpa găuri de 60 cm în pământ. Iarna, ingeră și zăpadă [1] [9] .
Reproducere

Comportamentul reproductiv a fost, de asemenea, rar studiat și numai în cazuri rare a fost observat în natură. În medie, fundul sălbatic asiatic atinge maturitatea sexuală în jurul vârstei de doi ani, deși prima împerechere are loc de obicei la vârsta de trei sau patru ani. Împerecherea are loc între aprilie și septembrie, cu un vârf în iunie și iulie. Gestația durează aproximativ unsprezece luni, după care se naște un singur bebeluș, după o scurtă naștere care durează puțin peste zece minute. Bebelușul poate merge după 15-20 de minute și începe să suge lapte după o oră și jumătate. În timpul creșterii, tânărul și mama rămân aproape: celelalte exemplare, inclusiv copiii anteriori, sunt îndepărtați de la mamă. Ocazional, armăsarii din populațiile teritoriale încearcă să-i alunge pe tineri și apoi să se împerecheze cu femela. Măgarii sălbatici pot trăi până la 14 ani în sălbăticie și până la 26 de ani în captivitate [9] [17] [18] .
Interacțiuni cu alte specii
Cel mai mare prădător al măgarului sălbatic este lupul . Cercetările au arătat că măgarii reprezintă 23% din prada acestui carnivor. Cu toate acestea, măgarul este capabil să se apere cu înverșunare de agresiune, chiar dacă preferă să găsească evadare în evadare: de fapt este foarte rapid și este capabil să galopeze la viteze de 70 km / h, menținând 50 km / h peste distanțe. Ocazional, în apropierea turmelor de măgari sălbatici, apar exemplare de gazelă gușă [6] [9] .
Paraziți
Printre cei mai comuni paraziți interni sunt nematodele , precum Trichostrongylus , Parascaris , Strongylus vulgaris , Dictyocaulus viviparus și Strongyloides stercoralis . De asemenea, viermii sunt prezenți și sunt cunoscute infestările cu coccidii , inclusiv Eimeria . Merită menționat faptul că pot apărea infecții reciproce cu calul lui Przewalski și cu calul domestic [19] .
Taxonomie
| Sistematica genului Equus conform Vilstrup și colab. , 2013 [20] | |||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
La fel ca toate ecvideele moderne, măgarul sălbatic asiatic aparține genului Equus . Deși morfologia molarilor săi inferiori este foarte variabilă, aceasta aparține în continuare grupului de stenonine sau ecvidee care nu sunt cal [21] . Analizele ADN efectuate în 2009 sugerează că ruda sa cea mai apropiată este kiang ( Equus kiang ) [22] . O specie fosilă strâns legată de măgarul sălbatic asiatic a fost măgarul sălbatic european ( Equus hydruntinus ), care a fost răspândit în Europa și Asia în Pleistocenul mediu și superior și poate că a dispărut doar în Holocen . Acesta a fost mai subțire și mai mare decât fundul sălbatic asiatic actual și a atins o înălțime de 160 cm la greabăn[3] . Inițial, unii experți abordaseră acest equin la măgarul sălbatic african, dar studiile anatomice au arătat că acesta este mai strâns legat de speciile asiatice [23] , ipoteză care a fost confirmată și de analizele genetice [24] . Atât măgarul sălbatic asiatic, cât și kiangul și măgarul sălbatic european sunt considerați un grup separat în cadrul equidelor stenonine, numit Hemionini , un nume care derivă din numele științific al măgarului sălbatic asiatic ( Equus hemionus ) și exprimă relația lor strânsă. Echidele cele mai apropiate de grupul Hemionini sunt măgarii adevărați și zebrele: calul domestic și Przewalski sunt înrudite doar cu ele, deși sunt clasificate și în genul Equus [22] [25] [26] . Denumirea științifică Equus hemionus a fost introdusă de Peter Simon Pallas în 1775; prima descriere științifică a avut la bază un exemplar din nord-estul Mongoliei [9] [27] .
În prezent există șase subspecii ale măgarului sălbatic asiatic, dintre care două au dispărut [28] :
- măgar sălbatic din Anatolia ( E. h. anatoliensis Haltenorth și Trense , 1956): Anatolia ; dispărut;
- Măgar sălbatic mongol sau hemi, cunoscut local ca chigetai ( E. h. Hemionus Pallas , 1775): Mongolia ;
- măgar sălbatic din Siria sau hemippo, cunoscut local ca achdari ( E. h. hemippus I. Goeffroy , 1855): Siria , Mesopotamia și Arabia ; dispărut în 1927;
- măgar sălbatic din India sau khur ( lecția E. h. khur , 1827): sudul Iranului , Pakistanului și nord-vestului Indiei ;
- măgar sălbatic din Turkestan sau kulan ( E. h. kulan ( Groves și Mazák , 1967)): Kazahstan și Turkmenistan ;
- Mâna sau onagerul persan, cunoscut local ca gur-khar ( E. h. Onager Boddaert , 1785): Iranul de Nord.
În trecut, a existat un mare dezacord în rândul cărturarilor cu privire la împărțirea în subspecii. În multe lucrări mai vechi, de exemplu, se disting șapte sau opt specii de măgari sălbatici asiatici, care astăzi sunt clasificați în cea mai mare parte ca subspecii. Excepția este kiangul , care astăzi este considerat o specie în sine [2] . Într-o recenzie a taxonomiei animalelor cu copite din 2011, prezentată de Colin Peter Groves și Peter Grubb , khur (Equus khur) și emippo (Equus hemippus) sunt recunoscute ca specii separate [29] .

Diferitele subspecii se caracterizează printr-o creștere generală a dimensiunii în direcția sud-vest / nord-est. Hemippo, care a locuit în zonele sud-vestice ale gamei, a atins o înălțime la greaban de doar 100 cm, în timp ce hemiul, prezent în zonele de nord-est, măsoară între 127 și 138 cm la greabăn. Kulanul are o înălțime de 108 până la 116 cm, iar onagerul și khur au ambele o înălțime care variază între 112 și 120 cm [2] . Potrivit unor cercetători, onagerul și kulanul ar aparține aceleiași subspecii, dar conform studiilor genetice moleculare actuale asupra filogeniei ecvideelor, cele două populații pot fi clar diferențiate între ele. O altă subspecie, măgarul sălbatic Gobi ( Equus hemionus luteus ), este uneori separat de hemiu datorită culorii diferite a hainei [2] [30] , dar experții observă cât de mult culoarea este foarte variabilă chiar și în aceeași populație [9] . Cu toate acestea, faptul că toate subspeciile sunt de origine monofiletică nu a fost confirmat de analizele ADN din 2009, deoarece onagerul sa dovedit a fi mai strâns legat de zebra montană [22] . Un alt studiu din același an indică totuși că toate subspeciile măgarului sălbatic asiatic sunt strâns legate [25] .
| Sistematica internă a speciei Equus hemionus conform lui Bennett și colab. , 2013 [31] | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
Analizele genetice ale anului 2017, extinse și la materialele subfosile și recente, indică o dezvoltare filogenetică mult mai complexă a măgarului sălbatic asiatic. Hemiul mongol este împărțit în cel puțin trei clade diferite, onagerul în două. Dintre acestea din urmă, linia vestică (Caucaz / Iran) este considerată în mare parte dispărută: doar câțiva indivizi prezenți în partea de est a zonei ar putea să o reprezinte în continuare. Potrivit autorilor studiului, cele trei clade diferite ale emionei ar putea fi asociate cu valuri de migrație diferite. Deoarece una dintre liniile evolutive apare amestecată cu cea a kiang-ului, este posibil ca kiang-ul să fie o formă de măgar sălbatic asiatic adaptat să trăiască în habitate de munte înalt [31] .
Echidele stenonine au fost raportate pentru prima dată în Asia între 2,5 și 3 milioane de ani în urmă și sunt probabil descendente din Equus cumminsi din America de Nord . Ruda cea mai apropiată a măgarului sălbatic asiatic este necunoscută, dar ar putea fi Equus namadicus sau Equus sivalensis , ambele datând din Pleistocen [21] . Primele fosile ale animalelor asemănătoare cu E. hemionus au fost găsite în Tologoj, Rusia și au o vechime de aproximativ un milion de ani [32] . Puțin mai tânără este subspecia fosilă E. h. nalaikhaensis , din Mongolia , ale cărei rămășițe din depozite ale râului Tuul datează de pe vremea evenimentului Jaramillo, fenomen de inversare a câmpului magnetic al Pământului care a avut loc în urmă cu aproximativ 900.000 de ani [33] . E. h. binagadensis , ale cărei fosile au fost găsite în Azerbaidjan și Iran , este o altă subspecie relativ mică a Pleistocenului mijlociu și superior care ar fi putut supraviețui până la Holocen [4] . Descoperiri din Tadjikistan , precum cele din Chudji și Ogzi-Kichik, datează din Pleistocenul superior. În special, în timpul Pleistocenului mijlociu și superior, măgarul sălbatic asiatic a apărut adesea simpatric cu măgarul sălbatic european ( Equus hydruntinus ) [34] . Această formă de măgar, care a dispărut la sfârșitul Pleistocenului, poate fi considerată, de asemenea, o simplă subspecie a măgarului sălbatic asiatic, deoarece studiile genetice din 2017 arată că avea legături de rudenie mai puternice cu reprezentanții Asiei Centrale și de Sud [31] .
Relațiile cu omul
Domesticire
Studiile ADN actuale confirmă faptul că astăzi toți măgarii domestici provin din măgarul sălbatic african . Cercetările filogenetice împart măgarii în mod distinct într-o linie evolutivă africană și asiatică, căreia îi aparține măgarul sălbatic asiatic ( Equus hemionus ). Întrebarea dacă această specie poate fi de asemenea domesticită și dacă acest lucru s-a întâmplat în trecut a fost întotdeauna controversată. În reprezentările Mesopotamiei antice (de exemplu pe stindardul Ur ), erudiții credeau că recunosc animale care nu erau nici cai, nici măgari, prin urmare au ajuns la concluzia destul de prematur că sumerienii și akkadienii au domesticit măgarii sălbatici asiatici, folosindu-i pentru a trage de carele.[35] . Cu toate acestea, în experimente de domesticire mai recente, nu a fost niciodată posibil să se antreneze aceste animale sau să le facă să-și piardă frica înnăscută față de oameni. Prin urmare, este mai plauzibil ca animalele domesticite în Mesopotamia să fie măgari sălbatici africani (care, în ciuda numelui, au fost prezenți și în Asia de Vest în timpurile preistorice).
Cu toate acestea, alți cercetători au emis ipoteza că sumerienii au folosit exemplare născute din trecerea dintre măgari și onager [36] .
depozitare
În vremurile istorice, măgarii sălbatici cutreierau semisertul asiatic în turme mari. Nevoia de a bea în mod regulat i-a împins în câteva puncte de apă prezente în habitatul lor. Marco Polo , în secolul al XIII-lea, a raportat despre prezența lor în număr mare în Arabia, Persia, Turkestan și Gobi [37] . Astăzi, principala amenințare la adresa speciei este pierderea habitatului datorată răspândirii așezărilor umane și dezvoltării economice a zonelor deseori inospitaliere, dar bogate în resurse . Concurența cu animalele mari de la fermă este, de asemenea, o mare problemă, deoarece măgarul sălbatic pierde de obicei atunci când vine vorba de a concura pentru pășuni și surse de apă. În plus, vânătoarea ilegală de măgari sălbatici, pe de o parte pentru hrană și, pe de altă parte, pentru pielea sa, reprezintă un pericol în creștere. Din anii 1990, populația a scăzut cu 52%. Toate subspeciile sunt amenințate, deși în diferite grade. Întreaga populație este clasificată de IUCN ca fiind aproape amenințată [1] .
Măsurile de protecție coordonate de Grupul de specialiști Equid al IUCN includ programe de relocare și reintroducere pentru populațiile individuale de măgari sălbatici asiatici, studii suplimentare privind nevoile ecologice ale speciilor și subspecii individuale, inclusiv utilizarea habitatului, un control mai mare al comerțului cu carne și sensibilizare proiecte pentru populația locală [1] .
Măgari sălbatici din Anatolia și Siria

Fundul sălbatic al Anatoliei a fost deja exterminat în antichitate . Mâna sălbatică siriană era încă numeroasă în secolul al XIX-lea în ceea ce este acum Irakul , unde se plimba în turme mari. Devenind din ce în ce mai rar, a fost vânat atât de des de soldații britanici și otomani în timpul primului război mondial, încât a fost împins în pragul dispariției. Ultimul măgar sălbatic al Siriei a fost ucis în 1927; un alt exemplar a murit în același an la Grădina Zoologică din Viena .
Emione
Hemi, cunoscut și sub numele de măgar sălbatic mongol, este cea mai comună subspecie, cu aproximativ 41.900 de exemplare care trăiesc în Mongolia, reprezentând trei sferturi din populația totală a speciei. Subspecia prezintă o tendință stabilă de creștere. Zonele protejate importante pentru conservarea sa sunt zona strict protejată a Marelui Gobi A și a Marelui Gobi B , în sudul Mongoliei. IUCN estimează o scădere a populației între 5 și 10% pe an. Deși hemionul este protejat oficial, specia intră adesea în conflict pentru pășuni și surse de apă cu populația nomadă și animalele sale domestice. Braconajul a crescut, de asemenea, din anii 1990, care este un alt motiv pentru declinul demografic accentuat [6] [9] . Alte 5 000 de exemplare trăiesc în nordul Chinei, mai mult de 3 300 dintre ele în rezervația naturală Kalamaili . Inițial, în anii 1980, rezerva găzduia doar aproximativ 360 de emisiuni și abia în 2000, după ce mai multe grupuri mai mari au imigrat din Mongolia, numărul acestora a început să crească considerabil [1] [14] [38] .
Khur

În anii 1960, khur a dispărut din părțile iraniene și pakistaneze din gama sa, precum și din aproape toate părțile indiene. A supraviețuit doar în Little Rann of Kutch din Gujarat , India . Aici, rezervația naturală Dhrangadhra a fost transformată într-un sanctuar special pentru conservarea acestor măgari rare. Khur este singura subspecie a măgarului sălbatic asiatic al cărui număr de indivizi a crescut constant în trecutul recent. În 2014, 4.000 de exemplare au fost înregistrate în Piccolo Rann. Aproximativ 30% dintre ei locuiesc în zona protejată. Animalele se răspândesc acum și în regiunile învecinate, precum Great Rann of Kutch și Blackbuck National Park . Populația a crescut constant, dar canalul Narmada pune în pericol starea de conservare a rezervației, deoarece atrage numeroși fermieri și numeroasele animale de companie cu apele sale dulci, intrând astfel în competiție directă cu khur. De asemenea, au fost solicitate permise pentru extragerea sării în Kutch . În afara sanctuarului, khur sunt împușcați de fermieri, deoarece sunt considerați dăunători culturilor sau sunt considerați competitori ai bovinelor pentru pășuni [1][13] .
Kulan

După ce kulanul a fost exterminat în Kazahstan și într-o mare parte din Turkmenistan, în anii '40 Uniunea Sovietică a creatrezervația naturală Badkhys , unde în următorii cincizeci de ani a constituit o turmă de 5000 de exemplare. Populația Kulan din Turkmenistan a scăzut recent dramatic: de la 5.000 în 1993 la aproximativ 580 în 2001. Odată cu independența Turkmenistanului , braconajul a crescut. Singura turmă originală rămasă, cea arezervației naturale Badkhys , găzduiește în prezent aproximativ 420 de animale. Il secondo gruppo più numeroso del Turkmenistan risiede nel bacino del Sarykamyš , dove vivono forse da 350 a 400 animali. Nel frattempo questa sottospecie è stata reintrodotta con successo anche altrove, ad esempio nella riserva naturale di Kaplankyr , nella riserva naturale di Barsakel'mes e nel parco nazionale Altyn-Ėmel' ; secondo informazioni del 2014, quest'ultimo ospita circa 2 500-3 000 esemplari. Tuttavia, la crescita della popolazione è limitata dal pesante sfruttamento agricolo dell'area circostante. La IUCN stima che la popolazione totale di kulan sia costituita al massimo da 2 000 individui adulti. Il calo della popolazione è dovuto soprattutto al venir meno dell'efficace controllo sulla caccia che esisteva ai tempi dell'Unione Sovietica [1] [39]
Dodici zoo europei gestiscono una popolazione di circa 50 kulan nell'ambito dell' EEP dell' Associazione europea degli zoo ( EAZA ). Coordinatore dell'EEP è Anna Mekarska dello zoo di Cracovia. La Società per il ripristino della cava di ghiaia di Weilbacher ( Gesellschaft zur Rekultivierung der Kiesgrubenlandschaft Weilbach , GRKW) sta attualmente cercando di insediare un gruppo di 6 maschi di kulan semiselvatici in una riserva naturale della Germania.
Onagro

Il numero di onagri nell'Iran settentrionale viene stimato in circa 780 individui suddivisi in due popolazioni separate. La popolazione più numerosa, di circa 630 esemplari, vive all'interno dell' area protetta di Turan , di 14 000 km² ; la seconda e più piccola vive nell'area protetta di Bahram-e-Goor, che copre 3 850 km² e ospita quasi 150 individui. Il governo iraniano è all'opera per garantire la sopravvivenza della sottospecie attraverso sanzioni draconiane contro il bracconaggio e la creazione di punti d'acqua supplementari [1] [40] .
Nell'ambito del Programma europeo per le specie minacciate ( EEP ) dell' Associazione europea degli zoo ( EAZA ) sono presenti attualmente circa 150 onagri ospitati in zoo gestiti con metodo scientifico. Coordinatore dell'EEP è Stephan Hering-Hagenbeck dello zoo Hagenbeck di Amburgo. Alcuni di questi onagri cresciuti in cattività sono stati rilasciati nel cratere Makhtesh Ramon nel deserto del Negev in Israele , ma gran parte della popolazione qui presente proviene da incroci con esemplari di kulan. Questa popolazione è costituita da circa 250 animali. Un gruppo molto più piccolo è stato introdotto in Arabia Saudita [1] [12] .
Etimologia
L'epiteto specifico utilizzato da Pallas, hemionus , deriva dalla parola greca ἡμιόνος ( hemiónos ), composta da ἡμί ( hemí ), «mezzo», e ὄνος ( ónos ), «asino». Il termine hemionos era già stato utilizzato nell' antichità da Omero e Aristotele per indicare gli animali dell' Anatolia e della Persia , ma originariamente si riferiva a muli e bardotti .
Anche l'origine del nome onager , usato da Pieter Boddaert , è greca: deriva da ὄναγρος ( ónagros ), una parola composta da ὄνος ( ónos ), «asino», e ἄγριος ( ágrios ), «selvatico». Da questo termine deriva il nome latino onagrus od onager . Greci e romani usavano queste parole esclusivamente per indicare l'asino selvatico africano, ma nella Vulgata si riferiscono anche all'asino selvatico asiatico.
Achdari , chigetai , gur-khar , khur e kulan sono nomi locali usati per indicare singole popolazioni dell'asino selvatico [2] .
Note
- ^ a b c d e f g h i j k l m ( EN ) Kaczensky, P., Lkhagvasuren, B., Pereladova, O., Hemami, M. & Bouskila, A. 2020, Equus hemionus , su IUCN Red List of Threatened Species , Versione 2020.2, IUCN , 2020.
- ^ a b c d e f g Colin Peter Groves e V. Mazák, On some taxonomic problems of Asiatic wild asses; with the description of a new subspecies (Perissodactyla; Equidae) , in Journal of Mammals , vol. 32, 1967, pp. 321-355.
- ^ a b Ingmar M Braun e Wolfgang Zessin, Pferdedarstellungen in der paläolithischen Wandkunst und der Versuch ihrer zoologisch-ethologischen Interpretation , in Ursus, Mitteilungsblatt des Zoovereins und des Zoos Schwerin , vol. 17, n. 1, 2011, pp. 4-26.
- ^ a b Véra Eisenmann e Marjan Mashkour, The small equids of Binagady (Azerbaidjan) and Qazvin (Iran): E. Hemionus Binagadensis nov. subsp. and E. Hydruntinus , in Geobios , vol. 32, n. 1, 1999, pp. 105-122.
- ^ a b Hans Klingel, Observations on social organization and behaviour of African and Asiatic Wild Asses (Equus africanus and Equus hemionus) , in Applied Animal Behaviour Science , vol. 60, 1998, pp. 103-113.
- ^ a b c d e Petra Kaczensky e Chris Walzer, Der Asiatische Wildesel - bedrohter Überlebenskünstler in der Wüste Gobi , in Zeitschrift des Kölner Zoos , vol. 51, n. 3, 2008, pp. 147-163.
- ^ Richard P. Reading, Henry M. Mix, Badamjaviin Lhagvasuren, Claudia Feh, David P. Kane, S. Dulamtseren e Sumyain Enkhbold, Status and distribution of khulan (Equus hemionus) in Mongolia , in Journal of the Zoological Society , vol. 254, 2001, pp. 381-389.
- ^ Martha I. Grinder, Paul R. Krausman e Robert S. Hoffmann, Equus asinus , in Mammalian Species , vol. 794, 2006, pp. 1-9.
- ^ a b c d e f g h i C. Feh, B. Munkhtuya, S. Enkhbold e T. Sukhbaatar, Ecology and social structure of the Gobi khulan Equus hemionus subsp. in the Gobi B National Park, Mongolia , in Biological Conservation , vol. 101, 2001, pp. 51-61.
- ^ Petra Kaczensky, Ralph Kuehn, Badamjav Lhagvasuren, Stephanie Pietsch, Weikang Yang e Chris Walzer, Connectivity of the Asiatic wild ass population in the Mongolian Gobi , in Biological Conservation , vol. 144, 2011, pp. 920-929.
- ^ G. Neumann-Denzau e H. Denzau, Remarks on the social system of the Mongolian wild ass (Equus hemionus hemionus) , in Erforschung der biologischen Ressourcen der Mongolei , vol. 10, 2007, pp. 177-187.
- ^ a b David Saltz, Mary Rowen e Daniel I. Rubenstein, The Effect of Space-Use Patterns of Reintroduced Asiatic Wild Asses On Effective Population Size , in Conservation Biology , vol. 14, n. 6, 2000, pp. 1852-1861.
- ^ a b N. Shah e Q. Qureshi, Social organization and spatial distribution of Khur (Equus hemionus khur) , in Erforschung der biologischen Ressourcen der Mongolei , vol. 10, 2007, pp. 189-200.
- ^ a b Ecology Habits of Equus hemionus and Gazella subgutturosa in Kalamaili Ungulate Reserve in Autumn , in Journal of Forestry Research , vol. 9, n. 2, 1998, pp. 131-132.
- ^ J. Lengger, F. Tataruch e Ch. Walzer, Feeding ecology of Asiatic Wild ass Equus hemionus , in Erforschung der biologischen Ressourcen der Mongolei , vol. 10, 2007, pp. 93-97.
- ^ SP Goyal, B. Sinha, N. Shah e HS Panwar, Sardar Sarovar Project - a conservation threat to the Indian wild ass (Equus hemionus khur) , in Biological Conservation , vol. 88, 1999, pp. 277-284.
- ^ David Saltz e Daniel I. Rubenstein, Population dynamics of reintroduced Asiatic wild ass (Equus hemionus) herd , in Ecological Applications , vol. 5, n. 2, 1995, pp. 327-335.
- ^ G. Neumann-Denzau, The birth of a Wild ass (Equus hemionus khur) in India's Little Rann of Kutch , in Erforschung der biologischen Ressourcen der Mongolei , vol. 10, 2007, pp. 219-226.
- ^ Johanna Painer, Petra Kaczensky, Oyunsaikhan Ganbaatar, Klaus Huber e Chris Walzer, Comparative parasitological examination on sympatric equids in the Great Gobi "B" Strictly Protected Area, Mongolia , in European Journal of Wildlife Research , 2010, DOI : 10.1007/s10344-010-0415-8 .
- ^ Julia T. Vilstrup, Andaine Seguin-Orlando, Mathias Stiller, Aurelien Ginolhac, Maanasa Raghavan, Sandra CA Nielsen, Jacobo Weinstock, Duane Froese, Sergei K. Vasiliev, Nikolai D. Ovodov, Joel Clary, Kristofer M. Helgen, Robert C. Fleischer, Alan Cooper, Beth Shapiro e Ludovic Orlando, Mitochondrial Phylogenomics of Modern and Ancient Equids , in PLoS ONE , vol. 8, n. 2, 2013, p. e55950.
- ^ a b Ann Forstén, Mitochondrial-DNA time-table and the evolution of Equus: comparison of molecular and paleontological evidence , in Annales Zoologici Fennici , vol. 28, 1992, pp. 301-309.
- ^ a b c Samantha A. Price e Olaf RP Bininda-Emonds, A comprehensive phylogeny of extant horses, rhinos and tapirs (Perissodactyla) through data combination , in Zoosystematics and Evolution , vol. 85, n. 2, 2009, pp. 277-292.
- ^ Ariane Burke, Vera Eisenmann e Graeme K. Ambler, The systematic position of Equus hydruntinus , an extinct species of Pleistocene equid , in Quaternary Research , vol. 59, n. 3, 2003, pp. 459-469.
- ^ Ludovic Orlando, Marjan Mashikour, Ariane Burke, Christophe J. Douady, Véra Eisenmann e Catherine Hänni, Geographic distribution of an extinct equid ( Equus hydruntinus : Mammalia, Equidae) revealed by morphological and genetical analyses of fossils , in Molecular Ecology , vol. 15, n. 8, 2006, pp. 2083-2093.
- ^ a b Ludovic Orlando, Jessica L. Metcalf, Maria T. Alberdi, Miguel Telles-Antunes, Dominique Bonjean, Marcel Otte, Fabiana Martin, Véra Eisenmann, Marjan Mashkour, Flavia Morello, Jose L. Prado, Rodolfo Salas-Gismondi, Bruce J. Shockey, Patrick J. Wrinn, Sergei K. Vasil'ev, Nikolai D. Ovodov, Michael I. Cherry Blair Hopwood, Dean Male, Jeremy J. Austin, Catherine Hänni e Alan Cooper, Revising the recent evolutionary history of equids using ancient DNA , in PNAS , vol. 106, 2009, pp. 21754-21759.
- ^ Hákon Jónsson, Mikkel Schubert, Andaine Seguin-Orlando, Aurélien Ginolhac, Lillian Petersen, Matteo Fumagallic, Anders Albrechtsen, Bent Petersen, Thorfinn S. Korneliussen, Julia T. Vilstrup, Teri Lear, Jennifer Leigh Myka, Judith Lundquist, Donald C. Miller, Ahmed H. Alfarhan, Saleh A. Alquraishi, Khaled AS Al-Rasheid, Julia Stagegaard, Günter Strauss, Mads Frost Bertelsen, Thomas Sicheritz-Ponten, Douglas F. Antczak, Ernest Bailey, Rasmus Nielsen, Eske Willerslev e Ludovic Orlando, Speciation with gene flow in equids despite extensive chromosomal plasticity , in PNAS , vol. 111, n. 52, 2014, pp. 18655-18660.
- ^ Peter Simon Pallas, Equus hemionus, Mongolis Dishikketaei dictus, describente , in Novi commentarii Academiae Scientiarum Imperialis Petropolitanae , vol. 19, 1774, pp. 394-414.
- ^ Daniel Rubenstein, Family Equidae (Horses and relatives) , in Don E. Wilson e Russell A. Mittermeier (a cura di), Handbook of the Mammals of the World. Volume 2: Hooved Mammals , Barcellona, Lynx Edicions, 2011, pp. 104-143, ISBN 978-84-96553-77-4 .
- ^ Colin Groves e Peter Grubb, Ungulate Taxonomy , Johns Hopkins University Press, 2011, pp. 13-17.
- ^ Karin Krüger, Vergleichende molekulargenetische Untersuchungen zur Phylogenese der Equiden unter besonderer Berücksichtigung von E. hemionus kulan und E. hemionus onager , Zurigo, 2003.
- ^ a b c E. Andrew Bennett, Sophie Champlot, Joris Peters, Benjamin S. Arbuckle, Silvia Guimaraes, Mélanie Pruvost, Shirli Bar-David, Simon JM Davis, Mathieu Gautier, Petra Kaczensky, Ralph Kuehn, Marjan Mashkour, Arturo Morales-Muñiz, Erich Pucher, Jean-François Tournepiche, Hans Peter Uerpmann, Adrian Bălăşescu, Mietje Germonpré, Can Y. Gündem, Mahmoud-Reza Hemami, Pierre-Elie Moullé, Aliye Ötzan, Margarete Uerpmann, Chris Walzer, Thierry Grange e Eva-Maria Geigl, Taming the late Quaternary phylogeography of the Eurasiatic wild ass through ancient and modern DNA , in PLoS ONE , vol. 12, n. 4, 2017, p. e0174216, DOI : 10.1371/journal.pone.0174216 .
- ^ Véra Eisenmann, Pliocene and Pleistocene Equids: palaeontology versus molecular biology , in Courier des Forschungsinstitutes Senckenberg , vol. 256, 2006, pp. 71-89.
- ^ TV Kuznetsova e VI Zhegallo, Equus (Hemionus) nalaikhaensis from the Pleistocene of Mongolia, the Earliest Kulan of Central Asia , in Paleontological Journal , vol. 43, 2009, pp. 574-583.
- ^ Ann Forsten e Sharif Sharapov, Fossil equids (Mammalia, Equidae) from the Neogene and Pleistocene of Tadzhikistan , in Geodiversitas , vol. 22, n. 2, 2000, pp. 293-314.
- ^ P. Ucko e G. Dimbleby, The Domestication and Exploitation of Plants and Animals , Aldine Pub., 2007, ISBN 978-0-202-36169-7 .
- ^ Juliet Clutton-Brock, Horse Power: A History of the Horse and the Donkey in Human Societies , 1992, ISBN 0-674-40646-X .
- ^ Rohini Rangarajan, The Influence of Ecto-Parasites on the behavior of the Indian Wild Ass (Equus hemionus khur), in the Wild Ass Sanctuary, Kutch, Gujarat , Manipal University, 2008.
- ^ Yang Weikang, An overview on the state of Equus hemionus in whole China , in Erforschung der biologischen Ressourcen der Mongolei , vol. 10, 2007, pp. 155-158.
- ^ VS Lukarevskiy e Yu. K. Goerlov, Kulan (Equus hemionus PALLAS 1775) in Turkmenistan , in Erforschung der biologischen Ressourcen der Mongolei , vol. 10, 2007, pp. 231-240.
- ^ L. Tatin, BF Darreh-Shoori, C. Tourenq, D. Tatin e B. Azmayesh, Up-date on the behaviour and status of the critically endangered Onager, Equus hemionus onager, from Iran , in Erforschung der biologischen Ressourcen der Mongolei , vol. 10, 2007, pp. 253-259.
Bibliografia
- Petra Kaczensky e Chris Walzer, Der Asiatische Wildesel - bedrohter Überlebenskünstler in der Wüste Gobi , in Zeitschrift des Kölner Zoos , vol. 51, n. 3, 2008, pp. 147-163.
- Hans Klingel, Observations on social organization and behaviour of African and Asiatic wild asses (Equus africanus and Equus hemionus) , in Zeitschrift für Tierpsychologie , vol. 44, 1975, pp. 323-331.
- P. Moehlman (a cura di), Equids: Zebras, Asses and Horses. Status survey and conservation action plan , Gland, Svizzera, IUCN, 2002.
- Ronald M. Nowak,Walker's Mammals of the World , Johns Hopkins University Press, 1999, ISBN 0-8018-5789-9 .
- Daniel Rubenstein, Family Equidae (Horses and relatives) , in Don E. Wilson e Russell A. Mittermeier (a cura di), Handbook of the Mammals of the World. Volume 2: Hooved Mammals , Barcellona, Lynx Edicions, 2011, pp. 104-143, ISBN 978-84-96553-77-4 .
Altri progetti
-
 Wikiquote contiene citazioni sull' onagro
Wikiquote contiene citazioni sull' onagro -
 Wikizionario contiene il lemma di dizionario « onagro »
Wikizionario contiene il lemma di dizionario « onagro » -
 Wikimedia Commons contiene immagini o altri file sull' onagro
Wikimedia Commons contiene immagini o altri file sull' onagro -
 Wikispecies contiene informazioni sull' onagro
Wikispecies contiene informazioni sull' onagro
Collegamenti esterni
- Asiatic wild ass videos, photos and facts - Equus hemionus , su ARKive (archiviato dall' url originale il 4 maggio 2016) .