Aripa insectelor
| |
Insights: Insecte Detaliu mărit al aripii unui Diptera |
| Anatomie |
| Cap · piept · abdomen |
| Antene · Piese bucale |
| Exoschelet · endoschelet |
| Aparate |
| Nervos · Digestiv Respirator · Excretor Secretor · Circulator Reproducător |
| Biologie |
| Dezvoltare post-embrionară |
Aripa insectelor este un organ, delegat în primul rând locomoției aeriene, prezent în toate pterigoții din stadiul imaginii , cu excepția grupurilor și speciilor care au dobândit apoteza secundară prin adaptare sau specializare. Este analog, dar nu omolog cu aripa altor animale zburătoare, cum ar fi păsările și liliecii . Organul este absent în ordinea Thysanura ( apterigoti ) pentru atterismul primitiv. În unele grupuri sistematice aripile suferă modificări morfologice și structurale care le transformă în organe atribuite altor funcții.
Caracteristici generale

Aripile sunt lamine extrem de turtite provenite din expansiunile unghiurilor posterioare - pleurale ale mezo- toracelui . Există maximum patru și pot fi reduse ca număr sau chiar absente.
Sunt organe vii, în care, în conductele speciale, numite coaste sau vene , hemolimfa curge și trec nervii și traheele . Nervurile ajută la confecționarea schelelor plăcilor de aripi și sunt denumite diferite în funcție de poziția lor pe aripă. Zonele delimitate de coaste se numesc celule . Numărul, cursul, gradul de ramificare a coastelor și, în consecință, numărul și forma celulelor, sunt caractere de mare importanță sistematică, în special în diptere și himenoptere . Coasta, care se întinde de-a lungul marginii anterioare, formează adesea o mărire, numită pterostigmă sau stigmat , a cărei funcție este mecanică, deoarece este asociată cu distribuția forțelor în timpul zborului.
Suprafața aripii este împărțită în trei regiuni morfologice. Cea mai extinsă parte, care include cele mai robuste nervuri (de la coastă până la cot) se numește regiunea remiges . În spatele acestui lucru se află regiunea anală sau go , în general traversată de coaste mai subțiri și ramificate sau chiar complet lipsită de venă. La multe insecte, între regiunea anală și extremitatea axilară posterioară a aripii există o extensie suplimentară, de dezvoltare redusă, numită regiunea jugală sau alula , aproape întotdeauna fără coaste. Regiunea jugală este absentă în paleoptere , un grup mai primitiv al pterigoților , reprezentat în prezent de ordinele odonata și efemeropterele . Regiunile anale și jugale sunt ușor de recunoscut prin venarea mai slabă și prin profilul marginii posterioare, deoarece are, în general, o adâncime mai mult sau mai puțin accentuată, trecând de la remiges la anal și de la acesta la jugal.
Sclerite axilare

În tipul primitiv, atribuibil celui al paleopterilor , aripa se conectează la torace fără nicio articulație: lamina dorsală este în continuitate cu tergitul, cea ventrală cu pleura. Odată cu evoluția, trecerea de la aripă la torace are loc printr-o articulație , formată din trei sclerite , numite pterale ; acestea, la rândul lor, sunt conectate la alte două sclerite articulare, numite bazale și subalare . Bazalul și subalarul se diferențiază de extremitatea dorsală a pleurei , respectiv de epistern și de epimer ; între ele intervine un proces pleural, situat exact deasupra suturii pleurale și care participă la articulația axilară.
În unele ordine ( Diptera , Himenoptera ), atacul costal al aripii anterioare, reprezentat de articulația dintre primul pter și bazal, este protejat de un sclerit numit tegula , diferențiat de mezonot .
Legarea organelor
Aripile, dacă sunt patru la număr, sincronizează adesea bătăile prin intermediul dispozitivelor de cuplare care au structuri și nume diferite în funcție de grupuri. Organele de legătură se găsesc între Rincoti , Lepidoptera și Hymenoptera .
În Rincoti legătura are loc prin intermediul unei articulații între structuri speciale ale marginilor celor două aripi: în Heteroptera marginea anterioară a aripii posterioare formează o îngroșare care se potrivește între două creste sclerificate prezente pe marginea posterioară a aripii anterioare. ; opusul apare la Homoptera . Lepidopterele au marginea posterioară a aripii anterioare îngroșate pentru a reține una sau mai multe peri dispuse de-a lungul marginii anterioare a aripii posterioare. În himenoptere există o structură bazată pe același principiu: marginea posterioară a aripii anterioare prezintă o îngroșare, pe care sunt agățate o serie de cârlige (numite hamuli ) dispuse de-a lungul marginii anterioare a aripii posterioare.
Tipuri morfofuncționale

În general, insectele din subclasa Pterygota au două perechi de aripi, ambele de consistență membrană, de aspect transparent și mai mult sau mai puțin irizat și, în cele din urmă, utilizate pentru zbor. Indiferent de formele meiotterismului , anumite adaptări morfologice, anatomice și funcționale pot fi găsite în unele grupuri sistematice. Cele mai importante sunt următoarele.
Organe complementare zborului
În aceste cazuri, aripa suferă astfel de modificări încât să-și piardă funcția primitivă și capătă alte funcții asociate întotdeauna cu zborul. În acest caz, aripa suferă o reducere puternică și se transformă într-o roată de echilibru , asumând funcții giroscopice . Aripile din spate ale Dipterelor și masculii Rincoti Coccidi și aripile din față ale masculilor din Strepsitteri sunt transformate în echilibratori .
Aripile au sclerificat

În aceste cazuri, tegumentul aripilor din față suferă o îngroșare mai mult sau mai puțin marcată a cuticulei (sclerificare). Îngroșarea afectează doar o parte a aripii sau se extinde la întregul organ și își asumă o funcție de protecție spre aripile posterioare. Gradul de sclerificare a tegumentului variază în funcție de grupul sistematic, iar aripile din față iau un nume specific în funcție de caracteristici:
- Elytra : acestea sunt aripile din față ale gândacilor , complet sclerificate și deosebit de robuste. Funcția de protecție atinge gradul maxim în unele coleoptere, unde elitrele fuzionează de-a lungul marginii lor posterioare, pentru a forma o singură structură de protecție. În timpul zborului, elitrele sunt ținute în aer, astfel încât aripile din spate să poată ieși. Acestea din urmă au o articulație specială a coastei, astfel încât aripile pot fi pliate complet, într-o stare de repaus, sub elitre. Unele gândaci au mecanisme extrem de sofisticate, care permit aripilor din spate să se desfășoare glisându-le sub elitre fără ca acestea să fie ridicate.

- Tegmină : sunt aripile din față ale ortopterelor și ale unor grupuri de homoptere Rincoti (superfamilii Membracoidea și Cercopoidea , inclusiv Rincoti cunoscute în mod obișnuit sub numele de puieți și scuipătoare ). Sunt total sclerificate, dar în ansamblu sunt mai puțin robuste decât elitrele gândacilor. [1]
- Emielitre : sunt aripile frontale ale generalității heteropterelor , cu excepția celor care au o dezvoltare a scutellum, astfel încât să acopere întregul abdomen. Hememiștii sunt sclerificați doar în jumătatea proximală, în timp ce restul este membranos. Aripile anterioare ale Heteropterelor, care au o dezvoltare notabilă a scutellumului, sunt în întregime membranare.
Acoperiri
În unele grupuri sistematice aripile, în timp ce rămân membrane, devin opace deoarece sunt acoperite cu peri, solzi sau ceară. Acoperirea poate da naștere și culorilor zonale, adesea de intensitate considerabilă. Învelișul de ceară se găsește, de exemplu, la muștele albi ( Rhynchota : Homoptera ). Învelișul solzos se găsește în majoritatea lepidopterelor . Acoperirea cu fire de păr sau peri se găsește în generalitatea trichopterelor și în mai multe himenoptere calcidoide .
Cereale și celule
După cum sa menționat anterior, coastele (sau venele) sunt tubuli sclerificați în interiorul cărora trec traheea , hemolimfa și nervii; în afară de funcția specifică, autorii au acordat o importanță deosebită acestui element morfologic, fundamental pentru determinarea taxonomică în unele ordine. Numărul, distribuția, cursul, ramificarea sunt caractere care identifică grupuri sistematice specifice, distingându-le de alte grupuri similare, prin urmare a fost acordată o atenție specială nomenclaturii referitoare la bob.
Celulele sunt zonele, de obicei membranate și transparente, delimitate de rețeaua de coaste. Acestea sunt împărțite în celule deschise și închise . Primele sunt situate de-a lungul marginii aripii, cele din urmă mai intern și complet delimitate de vene. Numărul de celule depinde de densitatea rețelei de nervuri și, în special, de gradul de ramificare a venelor longitudinale și de numărul de vene transversale. În majoritatea categoriilor sistematice rețeaua de coaste este relativ simplificată, dar în unele ordine, cum ar fi Odonata și Neuroptera , numărul de celule este mare, datorită prezenței numeroaselor coaste transversale.

Având în vedere diferențele considerabile dintre diferitele ordine, este adesea dificil, dacă nu chiar imposibil, să interpretăm cu certitudine omologiile: în unele cazuri este ușor să recunoaștem o coastă specifică în diferitele tipuri de aripi, în altele apare o simplificare structurală cu fuziunea sau dispariția coastelor sau ramificarea acestora. Rezultă că literatura nu este universal de acord cu standardele de nomenclatură; pe de altă parte, diferențele considerabile înseamnă că modelele teoretice de referință sunt mai potrivite pentru determinarea sistematică în cadrul comenzilor individuale. Prin urmare, în cursul istoriei entomologiei au fost propuse diferite modele de referință, care se suprapun parțial prin adoptarea aceleiași terminologii. Întrucât terminologia este fundamentală pentru clasificarea în ordinele Diptera și Hymenoptera , aici aprofundăm standardele de denumire cele mai utilizate în aceste zone și care se referă practic la sistemul Comstock-Needham , conceput pentru Diptera, și la cel al lui Ross, conceput pentru Himenoptere.
Sistem Comstock-Needham
Sistemul Comstock-Needham a fost conceput de John Henry Comstock și James George Needham în 1898 și este deosebit de potrivit pentru a defini nomenclatura nervurilor aripilor din Diptera . Convențiile de bază adoptate în acest sistem sunt următoarele:
- Coaste sunt împărțite în longitudinale și transversale ; primele sunt indicate cu o inițială capitală, cele din urmă cu o inițială mică. Nervurile longitudinale merg de la zona axilară către una dintre margini, în timp ce nervurile transversale leagă venele longitudinale între ele. Deoarece dispunerea nervurilor nu este întotdeauna bine definită, Autorii consideră unele nervuri transversale drept ramificații ale celor longitudinale.
- Fiecare celulă dintre două vene longitudinale ia numele venei care o delimitează anterior. De exemplu, celulele dintre sub-coastă și rază se numesc sub-coastă .
- Fiecare celulă dintre două ramuri ia numele ramurii care o delimitează posterior. De exemplu, celula dintre prima și a doua ramură a razei (R 2 și R 3 ) se numește radiu 3 (R 3 ).
- Când mai multe celule sunt delimitate de coaste transversale, toate iau același nume cu o numerotare ordinală care urmează ordinea proximal-distală. De exemplu, cele două celule dintre coaste și coaste și separate de prima humerală, sunt numite respectiv prima coaste (cea proximală) și a doua coaste (cea distală).
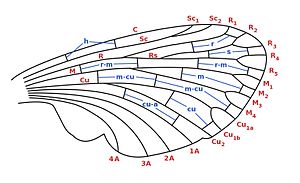 | Nomenclatura referitoare la coastele (stânga) și celulele aripilor (dreapta), conform sistemului Comstock-Needham. În roșu: nervuri longitudinale. În albastru: coaste transversale. Legendă: C : costuri; Sc : sub-coastă; R : radio; Rs : sectorul radio; M : mediu; Cu : cot; A : anal; h : humeral; r : radial; s : sectorial; rm : radio-media; m-cu : de dimensiuni medii; cu-a : cubito-anal. |  |
Procesând de la marginea anterioară la marginea posterioară, coastele longitudinale sunt denumite după cum urmează:
- Coasta ( C ): rulează de-a lungul marginii frontale. Are adesea întreruperi (una până la trei). În unele categorii sistematice există adaptări particulare în aripa posterioară care permit cuplarea marginii costale la aripa anterioară. În coleoptere , coasta este întreruptă de o articulație care permite aripii posterioare să se plieze în formă de ventilator în timpul fazei de repaus, astfel încât să fie complet acoperită de elitre.
- Subcosta ( Sc ): rulează paralel cu coasta și se termină curgând în aceeași coasta sau pe marginea anterioară în apropierea capătului distal. Se poate împărți și în două ramuri (anterioară și posterioară), indicate respectiv cu Sc 1 și Sc 2 .
- Raza ( R ): rulează în regiunea anterioară a aripii care se termină cu ramurile sale la capătul distal. În general este împărțit în două ramuri, una anterioară, numită ramură de rază ( R 1 ) și una posterioară, numită sector radial ( Rs ). La rândul său, sectorul radial se poate subdiviza de două ori, formând patru ramuri terminale indicate respectiv cu R 2 , R 3 , R 4 și R 5 în ordinea anterio-posterioară.
- Mijlociu ( M ): este cea mai robustă coastă, care se desfășoară în regiunea mediană a aripii până la marginea distală. Această coastă este împărțită în două ramuri, dintre care cea anterioară se consideră că a dispărut, în timp ce cea posterioară se împarte de obicei de două ori, formând un total de 4 ramuri indicate respectiv cu M 1 , M 2 , M 3 și M 4 în anterioară- ordinul posterior.
- Cubit ( Cu ): rulează în regiunea posterioară a aripii și poate fi împărțit în două ramuri, indicate respectiv cu Cu 1 și Cu 2 în ordine anterio-posterioară. Potrivit altor autori, ramura anterioară este împărțită în două ramuri indicate respectiv cu Cu 1a și Cu 1b .
- Anal : au un număr de 1-3 sau, uneori, chiar mai mare; în general nu sunt ramificate și se desfășoară paralel în regiunea anală a aripii. Acestea sunt identificate cu o numerotare ordinală după o ordine anterioară posterioară: primul anal ( 1A ), al doilea anal ( 2A ) etc;
- Jugali : pot fi prezenți în regiunea jugală.
Nervurile transversale leagă nervurile longitudinale între ele. Nomenclatura adoptată în sistemul Comstock-Needham este în general derivată din cea a nervurilor longitudinale și se referă la venele aferente. În simbolologie sunt folosite doar caractere mici pentru a le distinge de cele longitudinale. De la marginea costală la cea anală, coastele transversale sunt denumite după cum urmează:
- Omere ( h ): sunt incluse între coastă și subcostă .
- Radial ( r ): între ramura razei (R 1 ) și sectorul radial (Rs și R 2 ).
- Sector ( uri ): printre ramurile sectorului radial.
- Radio-media ( rm ): între ele radio și media.
- Medial ( m ): între ramificațiile mediei.
- Medio-cubitali ( m-cu ): între medie și cot.
- Cubitali ( cu ): între ramurile cubitului .
- Cubitus-anal ( cu-a ): între cubitus și primul anal.
- Anal ( a ): între analul longitudinal, dar în general acestea nu sunt prezente.
Modelul Ross
Sistemul Comstock-Needham, conceput fundamental pentru vena de aripă a Dipterelor, are unele limite interpretative (în raport cu omologiile) atunci când este aplicat pe deplin la himenoptere . De la sfârșitul secolului al XIX-lea până în anii 1960, s-au propus diferite criterii de nomenclatură pentru himenoptere; controversele, care au făcut dificilă atingerea unui standard interpretativ, se datorează încercărilor de interpretare a bobului pe diferite criterii: evolutive, anatomice (relația dintre coaste și trahee), morfo-anatomice (relația dintre coastele longitudinale și scleritele axilare). Unele modele au propus derivări din sistemul Comstock-Needham, altele au ignorat omologiile cu celelalte ordine ale insectelor, altele au încercat să extrapoleze teoriile anterioare într-o sinteză. Diatriba fundamentală constă în interpretarea coastei medii: conform interpretărilor, mijlocul ar fi dispărut sau ar avea un curs comun cu Raza în tractul proximal și apoi ar deriva într-un tract liber în regiunea remigilor distali.
 | Nomenclatură referitoare la coastele aripii frontale, conform modelului Ross. Legendă: C : costuri; Sc : sub-coastă; R : radio; Rs : sectorul radio; M : mediu; Cu : cot; A : anal; h : humeral; r : radial transversal; rm : radio-media; m-cu : mijlociu sau recurent; cu-a : cubito-anal; cv : transvers anal. |
În 1936 , Herbert H. Ross a elaborat un model bazat pe ipoteza unei adaptări a venelor la distribuția forțelor de tracțiune în regiunea remigentă a aripii [2] . Angrenarea hamulilor aripii posterioare pe marginea anterioare, determină o tracțiune în direcție transversală în zona mediană a aripii anterioare (la înălțimea stigmatului). Coaste longitudinale ar suferi, prin urmare, abateri marcate în direcția antero-posterioară, astfel încât să ofere aripii o rezistență intrinsecă la tracțiune. Prin adoptarea modelului Ross, nomenclatura specifică pentru aripa din față ar fi practic împrumutată din sistemul Comstock-Needham. Nervurile longitudinale sunt aceleași ca în sistemul Comstock-Needham, dar suferă fuziuni și abateri în direcția transversală:
- Costa ( C ). Se desfășoară de-a lungul frontierei anterioare până la pterostigma, de care este separată printr-o întrerupere evidentă.
- Subcosta ( Sc ). Este împărțit în două ramuri, înainte de pterostigmă: ramura superioară (Sc 1 ) se conectează la coastă, cea inferioară deviază posterior și fuzionează cu ramura de rază (Sc 2 + R 1 ) și apoi se separă din nou la stigmă.
- Radio ( R ). Este fuzionat cu media din tractul proximal, apoi se separă de acesta și se împarte în ramura de rază (R 1 ) și sectorul radial (Rs). Ramura de rază fuzionează cu sub-coasta 1 și apoi se separă la stigmat. Sectorul radial deviază posterior și fuzionează cu media. La înălțimea stigmatului se separă de medie cu o abatere înainte și continuă longitudinal.
- Mediu ( M ). Această coastă este cea care a dat naștere diatribelor pe nomenclatură. Conform schemei lui Ross, acesta este fuzionat inițial cu radiu, apoi se separă și se îmbină cu cotul 1 (M + Cu 1 ). Într-o secțiune ulterioară se separă și deviază înainte pentru a fuziona cu sectorul radial (M + Rs). La înălțimea stigmatului se separă din nou și rulează longitudinal în partea mediană a regiunii distale remigate.
- Cubit ( Cu ). După tractul bazal se împarte în cele două ramuri anterioare și posterioare. Cubit 1 suferă o deviere înainte și se contopește cu media (M + Cu 1 ). Mai târziu se separă și are o cale caracterizată prin abateri transversale marcate la înălțimea stigmatului. Aici este împărțit în ramurile Cu 1a , cu un curs longitudinal, și Cu 1b cu un curs transversal.
- Anal longitudinal ( 1A , 2A , 3A , 4A ).
Coaste transversale mențin, de asemenea, aceeași terminologie:
- Humeral ( h ). Conectează coasta de coastă. Se află în general în partea proximală a aripii, separând o celulă costală cu dezvoltare redusă.
- Radial ( 1r , 2r ). Sunt două la număr și conectează ramura razei la sectorul radial.
- Radio-medial ( 2r-m , 3r-m ). Acestea sunt două la număr (uneori trei) și conectează ramura posterioară a sectorului radial la medie.
- De dimensiuni medii ( 1m-cu , 2m-cu ). Acestea sunt una sau două la număr și conectează media cu ramura anterioară a cotului. În himenoptere, acestea sunt foarte importante, în scopuri sistematice, și sunt, de asemenea, numite vene recurente de mulți autori.
- Cubitus-anal ( cu-a ). Conectează ramura anterioară a cubitusului la primul anal.
- Anal transversal ( CV ). Conectează primul anal la al doilea anal.
În prezent nu există un standard de nomenclatură universal împărțit pentru himenoptere, dar cea mai răspândită orientare [3] [4] este adoptarea schemei Herbert H. Ross, deși modelul original a suferit unele revizuiri în cursul unor ani.
Mecanisme de zbor

Mișcările aripilor insectelor sunt extrem de complicate și diferite în funcție de ordine. Acestea includ, în general, acțiuni de ridicare, depresie, extensie, flexie, torsiune (pronație și supinație), precum și modificări ale formei lor (pentru pliere etc.). Înălțimea și depresia sunt, în general, guvernate de mușchii indirecți ai aripilor și de mușchii accesorii indirecți ai aripilor . Extinderea, torsiunea și flexia sunt guvernate în schimb de mușchii alari direcți .
Mușchii alari indirecți sunt grupați în două grupe funcționale, dorsal și tergo-sternal , care acționează în opoziție cu primii. Mușchii alari accesori indirecți sunt împărțiți în tergo-pleural, pleuro-sternal, tergo-coxal și tergo-trohanteric. În general, mușchii indirecți deformează cutia toracică prin transmiterea deformării către expansiunile sale, ceea ce duce la o înălțime sau o depresiune a aripilor.
Mușchii alari direcți sunt împărțiți în mușchi axilari , bazali și subalari . Acești mușchi conectează scleritele axilare la alte sclerite toracice și, prin urmare, acționează direct pe aripi, complicându-și mișcările.
Compararea insectelor cu alte organisme înaripate
Dintre animalele care în prezent pot zbura, păsările și liliecii au rezolvat problema în moduri diferite și cu diverse repercusiuni în morfologia și eco-etologia lor.
Păsările ( homeoterme ) au modificat membrele anterioare din segmentul terminal (autopodium), oferindu-le pene (remiguri și acoperiri). Nu le pot folosi în alte moduri și merg doar cu ajutorul membrelor posterioare.
Liliecii (de asemenea homeotermi) au extins pe scară largă pielea corpului periferic într-un pli subțire membranos ( patagio ) care valorifică picioarele din față, picioarele din spate și coada și este susținut de degetele de la picioare (2 ° -5 °) ale picioarele din față, având falange foarte alungite. În consecință, practic nu mai pot merge. Mai mult, acest pliu este în mare parte gol, dispersează sau colectează multă căldură și permite evaporarea unei cantități mari de apă. Prin urmare, aceste mamifere au trebuit să se adapteze la o activitate nocturnă sau de seară la toate latitudinile. În timpul zilei trebuie să se refugieze, înveliți în mantaua patagio-ului lor, în medii întunecate și să închidă sistemul de termoreglare intrând în hipotermie . Pentru a se orienta în zborul spațial și a captura prada, au diferențiat apoi un aparat de ecolocație complex și perfect.
Pe de altă parte, insectele ( peciloterme ) nu au renunțat la nicio pereche de picioare și și-au dezvoltat aripile în mod independent, ca expansiuni dorsal-laterale ale tegumentului din ultimele două segmente ale toracelui, care au aceeași structură și același tip de protecție. modalități ale tegumentului general al corpului. Din acest motiv și din alte motive, ei s-au putut adapta la cele mai variate medii și la cele mai diverse condiții de viață, invadând ținuturile emergente și apele dulci și salmastre.
Meiotterism
La insectele cu aripi (Pterygotes), meiotterismul este frecvent. Acest termen se referă la reducerea mai mult sau mai puțin marcată a dezvoltării aripilor, de la forme de brahterism ușor (aripi mai scurte, dar care păstrează în mare măsură forma și coastele tipice) până la tobe reale (dispariția totală a aripilor).
Meiotterismul este o adaptare secundară, adesea corelată cu factorii de mediu, etologici, funcționali, care se manifestă în totalitatea speciilor unui ordin sau în grupuri sistematice unice.
Notă
- ^ tegmine in Vocabulary - Treccani , pe www.treccani.it . Adus la 5 ianuarie 2020 (Arhivat din original la 9 iulie 2021) .
- ^ Herbert H. Ross, The ancestries and wing venation of the Hymenoptera , în Annals of the Entomological Society of America , XXIX, n. 1, 1936, pp. 99-111.
- ^ Viggiani. Op. Cit. , p. 163.
- ^ Francesco Intoppa, Maria Gioia Piazza, Graziella Bolchi Serini, Nomenclatura și interpretarea venulației aripilor Apoidea: o revizuire critică ( PDF ), în Redia , LXXXIII, Anexă, 2000, pp. 1-24.
Bibliografie
- Ermenegildo Tremblay, Entomologie aplicată. Volumul I , ediția a III-a, Napoli, Liguori Editore, 1985, ISBN 88-207-0681-4 .
- Ermenegildo Tremblay, Entomologie aplicată. Volumul II Partea I , ediția I, Napoli, Liguori Editore, 1981, ISBN 978-88-207-1025-5 .
- Ermenegildo Tremblay, Entomologie aplicată. Volumul II Partea II , ediția I, Napoli, Liguori Editore, 1986, ISBN 88-207-1405-1 .
- Ermenegildo Tremblay, Entomologie aplicată. Volumul III Partea I , ediția I, Napoli, Liguori Editore, 1991, ISBN 88-207-2021-3 .
- Antonio Servadei, Sergio Zangheri; Luigi Masutti, Entomologie generală și aplicată , Padova, CEDAM, 1972.
- Guido Grandi, Institutions of general entomology , Bologna, Calderini, 1966, ISBN 88-7019-084-6 .
- Aldo Pollini, Manual de entomologie aplicată , Bologna, Edagricole, 2002, ISBN 88-506-3954-6 .
- Gennaro Viggiani, Lupta biologică și integrată , Napoli, editor Liguori, 1977, ISBN 88-207-0706-3 .
- John Henry Comstock, James George Needham, Aripile insectelor , Ithaca, New York, The Comstock Publishing Company, 1898.
- (EN) Takahiro Ohde, Toshinobu Yaginuma, Teruyuki Niimi, Insect Morphological Diversification Through the modification of Wing Serial homologs ( abstract ), în Science, vol. 340, n. 6131, 26 aprilie 2013, pp. 495-498, DOI : 10.1126 / science.1234219 .
Elemente conexe
Alte proiecte
-
 Wikimedia Commons conține imagini sau alte fișiere pe aripa insectelor
Wikimedia Commons conține imagini sau alte fișiere pe aripa insectelor