Fosforilarea oxidativă
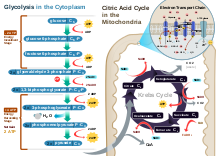
Fosforilarea oxidativă este un proces biochimic celular fundamental și omniprezent pentru producerea de energie prin sinteza ATP . Aceasta este faza finală a respirației celulare , după glicoliză , decarboxilarea oxidativă a piruvatului și ciclul Krebs .
Locația fizică a procesului este întotdeauna în calea unei membrane : în eucariote apare la membrana mitocondrială internă , în procariote la membrana celulară .
Se compune din două faze:
- Lanțul de transport al electronilor : în acest proces, electronii transportați de NADH și FADH 2 sunt schimbați de lanțul enzimatic transmembranar generând un gradient de protoni
- sinteza ATP prin fosforilarea ADP de către enzima ATP sintază
Lanțul de transport al electronilor
Coenzimele „încărcate” în matricea mitocondrială în timpul ciclului Krebs sunt utilizate în așa-numitul lanț de transport al electronilor . Acesta „deversări“ cele două coenzimi NADH și FADH 2 (oxidarea ei la NAD + și FAD), transferarea electronii la unele de transport molecule , în esență , citocromilor (particulare enzimatice complexe proteice coenzima capabile de oxigen de legare). Citocromii sunt responsabili de transportul electronilor către enzimele NADH-Q reductază , succinat dehidrogenază , citocrom reductază și citocrom c oxidază , capabile să „pompeze” ionii H + care provin din oxidarea NAD și FAD din matrice în spațiul intermembranar printr-un transport activ prin membrana interioară, generând un gradient de protoni . Protonii nu pot intra din nou în matrice pentru a restabili echilibrul, deoarece membrana interioară nu este permeabilă spontan la trecerea lor. Reintrarea are loc la sfârșitul procesului prin intermediul enzimei ATP-sintază , care valorifică energia potențialului lor electrochimic pentru sinteza ATP (vezi mai jos). Din reducerea acceptorului final de electroni, oxigenul și din ionii H + derivă molecule de apă care se adaugă celor produse prin glicoliză .
Generarea ATP
Când electronii ajung la oxigen, lanțul de transport este întrerupt pentru o fracțiune din timpul necesar reechilibrării protonului. Diferența de potențial și pH este anulată, iar protonii revin la matrice prin complexul proteic (canalul proteic) al ATP sintazei ( chemosmoza ). Energia potențială eliberată activează reacția endoergonică dintre grupurile ADP și fosfat cu sinteză de ATP. Reacția este:
ADP 3- + H + + P i ATP 4- + H 2 O ADP 3- + H + + P i ATP 4- + H 2 O

Vom avea apoi ultima formă de conservare a energiei, stocată în legăturile a 38 de molecule ATP prin glicoliză, ciclul Krebs și fosforilarea oxidativă. Trebuie amintit că acesta este randamentul maxim teoretic al unei molecule de glucoză, dar realul este între 36 și 32 de molecule de ATP, datorită unei permeabilități minime a membranelor la electroni și a randamentului real mai scăzut de NADH și FADH2, care este respectiv de 2,5 și 1,5 ATP și nu de 3 și 2 (vezi sub „Stoichiometrie”). Celula va putea elibera energia conținută în ATP atunci când este necesar, prin procesul de hidroliză , catalizat de enzima ATPază dependentă de Mg ++ .
După sinteza ATP, sistemul de navetă malat-aspartat , un sistem de doi transportori, este utilizat pentru a restabili concentrația de NADH în interiorul matricei. NADH nu poate traversa membrana mitocondrială, astfel încât electronii din NADH sunt transportați prin ea. În citosol, NADH oxidează și transformă oxaloacetat în malat datorită enzimei malat dehidrogenazei . Acesta din urmă traversează membrana mitocondrială internă și este reoxidat de NAD + . Oxaloacetatul printr-o reacție de transaminare cu glutamat devine aspartat grație enzimei aspartat transaminază și este transportat în spațiul intermembranar.
Stoichiometrie
Se produc în total 2,5 molecule de ATP pentru fiecare moleculă NADH oxidată. Pentru fiecare moleculă de FADH 2 se produc în schimb 1,5 molecule de ATP.
Prin oxidarea glicolitică a unei molecule de glucoză , conversia ulterioară a moleculelor de piruvat în acetil-CoA și în final ciclul Krebs , sunt sintetizate un total de 10 molecule de NADH și 2 de FADH 2 . Valoarea energetică stocată în aceste molecule este convertită într-un total de 28 de molecule de ATP [(10x2.5) + (2x1.5)].
Cu toate acestea, este important să se ia în considerare faptul că aceste valori sunt doar teoretice, în timp ce în realitate randamentul reacției de sinteză a ATP este mai mic, datorită permeabilității, deși scăzută, a membranei la protoni.
Inhibitori
Unele molecule sunt capabile să inhibe unele etape de fosforilare oxidativă. Din acest motiv, acestea sunt efectiv toxine .
- Cianura blochează lanțul de transport al electronilor, deoarece se leagă de Fe 3+ în citocromul 3 mai eficient decât oxigenul , împiedicând citocromul să combine electronii cu oxigenul.
- Oligomicina inhibă ATP sintaza , prevenind reîncărcarea moleculelor ATP de-a lungul gradientului de protoni .
- CCCP ( m-clor-carbonilcianură-fenilhidrazină ) distruge gradientul de protoni permițând protonilor să traverseze membrana. Fără gradient, de fapt, sintaza ATP nu poate funcționa.
- Chiar și un detergent (sau o substanță capabilă să distrugă membranele celulare) este capabil să distrugă gradientul de protoni , pur și simplu prin eliminarea membranei prin care este generat.
- Rotenona împiedică transferul de electroni din centrele Fe-S ale complexului I către ubiquinonă . Electronii care intră în lanț prin acest complex sunt cei care provin din NADH și constituie partea preponderentă a potențialului capabil să genereze gradient de protoni .
Pentru fiecare dintre aceste toxine, acțiunea toxică nu se limitează doar la procesul de fosforilare oxidativă, ci are efecte de impact asupra unei mari părți a metabolismului celular. Dacă se administrează oligomicină, de exemplu, H + nu pot trece prin ATP sintază. Acest lucru induce o stabilizare a gradientului de protoni , care pe termen lung face pompele de protoni ineficiente, care nu funcționează în prezența unor gradienți excesiv de mari. Dacă aceste pompe cedează, moleculele NADH și FADH 2 nu vor mai fi oxidate din nou în NAD + și FAD. Aceasta înseamnă că celelalte procese metabolice care îl utilizează, precum ciclul Krebs și glicoliza , nu vor mai putea funcționa.
Compuși reactivi ai oxigenului
În timpul fosforilării oxidative, se pot forma compuși de oxigen tranzitori cu reactivitate puternică, și anume peroxid de hidrogen , anion superoxid și radical hidroxil . Cu toate acestea, citocrom c oxidaza, complexul IV, este capabil să mențină controlul steric asupra moleculelor create, reducând oxigenul în apă.
Peter Dennis Mitchell
„Aspectul situației actuale de consens pe care l-am găsit cel mai remarcabil și admirabil este altruismul și generozitatea cu care primii oponenți ai ipotezei chemosmotice nu numai că au acceptat-o, ci au contribuit în mod activ la promovarea acesteia la statutul unei teorii.” |
| ( Peter Mitchell, Discurs la ceremonia Premiului Nobel, 1978 ) |
Fosforilarea oxidativă a rămas un mister până la începutul anilor 1960 și datorită biochimistului englez Peter D. Mitchell (și cercetătorilor ulteriori) cunoaștem acum acest proces în detaliu. Datorită acestei cercetări, în 1978 a primit Premiul Nobel pentru chimie .
Bibliografie
- Lehninger, D., Cox, M. Principiile Biochimiei Ediția a III-a, Worth Publications, New York, NT., 2001.
- David L. Nelson, Michael M. Cox, Principiile de biochimie ale lui Lehninger , ediția a III-a, Bologna, Zanichelli , februarie 2002, ISBN 88-08-09035-3 .
- Jeremy M. Berg, John L. Tymoczko; Lubert Stryer, Biochimie , ediția a V-a, Bologna, Zanichelli , octombrie 2003, ISBN 88-08-07893-0 .
Elemente conexe
- Respirație celulară
- Lanțul de transport al electronilor
- Sistem de navetă malat-aspartat
- Sistem de transfer glicerol fosfat
- Gradient de protoni electrochimici transmembranari
- Decuplarea proteinelor
Alte proiecte
-
 Wikimedia Commons conține imagini sau alte fișiere despre fosforilarea oxidativă
Wikimedia Commons conține imagini sau alte fișiere despre fosforilarea oxidativă
linkuri externe
- ( EN ) Model molecular interactiv de succinat dehidrogenază , pe ufp.pt. Adus la 8 august 2006 (arhivat din original la 25 septembrie 2006) .
- ( EN ) Model molecular interactiv al coenzimei Q - citocrom c reductază , pe ufp.pt. Adus la 8 august 2006 (arhivat din original la 5 august 2008) .
- ( EN ) Model molecular interactiv al citocrom c oxidazei , pe ufp.pt. Adus la 8 august 2006 (arhivat din original la 25 septembrie 2006) .