Parasaurolophus
| Parasaurolophus | |
|---|---|
 | |
| Gama geologică | |
| Starea de conservare | |
Fosil | |
| Clasificare științifică | |
| Domeniu | Eukaryota |
| Regatul | Animalia |
| Phylum | Chordata |
| Clasă | Sauropsida |
| Superordine | Dinozauria |
| Ordin | † Ornitischia |
| Subordine | † Ornitopoda |
| Familie | † Hadrosauridae |
| Trib | † Parasaurolophini Prieto-Márquez și colab. , 2013 |
| Tip | † Parasaurolophus Parcuri , 1922 |
| Nomenclatura binominala | |
| † Parasaurolophus walkeri Parcuri, 1992 | |
| Sinonime | |
| |
| Specii | |
Parasaurolofo (al cărui nume înseamnă „lângă șopârla cu crestă ” în referință la Saurolophus [1] ) este un gen dispărut de dinozaur ornitopod care a trăit în Cretacicul târziu , în urmă cu 76,5-74,5 milioane de ani ( Campanian ), în statele Alberta ( Canada) ), New Mexico și Utah ( Statele Unite ), America de Nord . Parasaurolofo era un erbivor capabil să meargă atât în posturile bipede, cât și în cele patrupedale. Genul conține trei specii: specia tip P. walkeri , P. tubicen și P. cyrtocristatus . Genul a fost descris în 1922 de William Parks pe baza unui craniu și a unui schelet parțial, din Alberta .
Creasta goală conținea o serie de tuburi care alungeau pasajele nazale: un fel de versiune osoasă a trunchiului unui elefant. Poate că l-a folosit pentru a da apeluri și pentru a putea ține legătura cu semenii săi din inima pădurilor. Fiecare specie avea o creastă diferită, deci este probabil ca și apelurile lor să fie diferite. Creșterea osoasă extinsă făcea parte din capul animalului și în specia parasaurolophus walkeri, (deasupra) era lungă ca și craniul. Era acoperit cu piele care probabil îl lega de gât.
Parasaurolophus era un hadrosaurid , o familie de dinozauri mijlocii până la mari care s-a diversificat în timpul Cretacicului târziu și cunoscută pentru facturile lor de rață și gama largă de forme ale crestelor lor craniene. Parasaurolophus în special se distinge prin creasta craniană tubulară lungă și largă care se proiectează spre partea din spate a craniului . Charonosaurus din China deținea, de asemenea, o structură similară, prin urmare este considerată ruda sa cea mai apropiată, în timp ce alții consideră că este o specie asiatică a genului. Printre diferitele ipoteze propuse cu privire la funcționarea creastei se numără utilizarea acesteia ca afișaj sexual, ca cutie de rezonanță acustică și ca mijloc de termoreglare . În ciuda faptului că este bine cunoscut atât în paleontologi, cât și în mass-media culturală, Parasaurolophus este unul dintre cele mai rare hadrosauride și este cunoscut doar dintr-o mână de exemplare bine conservate.
Descriere
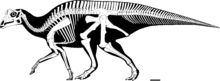
La fel ca majoritatea dinozaurilor, scheletul cunoscut al Parasaurolophus este încă incomplet. Lungimea exemplarului tip P. walkeri este estimată la 9,5 metri (31 picioare), pentru o greutate estimată la 2,5 tone (2,8 tone scurte). Craniul are aproximativ 1,6 metri lungime, inclusiv creasta, în timp ce craniul de tip P. tubicen are o lungime de peste 2 metri (6 picioare, 7 inci), indicând un animal mult mai mare. Singurul membru cunoscut este relativ scurt pentru un hadrosaurid, cu un omoplat scurt, dar lat. Femurul măsoară 103 centimetri (41 inci) în lungime în P. walkeri și este extrem de robust pentru lungimea sa, în comparație cu alte hadrosauride. Humerusul și oasele pelvine sunt, de asemenea, extrem de puternice și bine construite.
La fel ca multe alte hadrosauride, Parasaurolophus a fost capabil să meargă pe ambele picioare și pe toate patru. Probabil în timp ce căuta hrană, animalul a adoptat o locomoție cvadrupedală mai lentă, în timp ce în caz de pericol sau pentru a se deplasa mai repede a adoptat o poziție bipedă. Spinele neuronale ale vertebrelor erau foarte mari, o caracteristică comună la lambeosaurini ; mai sus pe șolduri, au crescut în înălțime de-a lungul spatelui până la coadă. Unele amprente ale pielii, cunoscute din speciile P. walkeri , prezintă pielea acoperită cu tuberculi uniformi mici care nu se suprapun, dar nu sunt raportate structuri mai mari.
Craniu
Cea mai evidentă caracteristică a Parasaurolophus este, fără îndoială, creasta craniană lungă și elaborată, care ieșea din spatele capului și este compusă dintr-o extensie a premaxilei și a oaselor nazale . William Parks, când a descris animalul, a emis ipoteza că creasta ar putea fi atașată la ceafă a animalului printr-un ligament , citând prezența unei crestături posibil patologice ca dovadă. Deși această idee pare oarecum puțin probabilă, în reconstrucțiile paleontologice, Parasaurolophus este uneori reconstruit cu o clapă de piele trasă între creastă și ceafă. Creasta, pe interior, era goală, cu tuburi principale distincte pentru fiecare nară, care se extindeau până la capătul creastei înainte de a inversa direcția înapoi spre craniu. Aceste conducte au fost mai simple la P. walkeri și mai complexe la P. tubicen , unde unele conducte erau orb în timp ce altele s-au încrucișat și s-au separat. În timp ce speciile P. walkeri și P. tubicen aveau creste lungi și voluminoase ușor curbate, speciile P. cyrtocristatus aveau o creastă foarte scurtă, cu un profil mai circular.
Clasificare

După cum sugerează și numele, Parasaurolophus a fost inițial asociat cu genul Saurolophus , datorită prezenței în ambele genuri a unei creste craniene proiectate în spate. Cu toate acestea, ulterior a fost reevaluat ca membru al lambeosaurini , o subfamilie a familiei hadrosauridi, în timp ce Saurolophus este clasificat ca adrosaurini . PARASAUROLOPHUS este clasificată de obicei într - o ramură separată de lambeosaurins clasice caracterizate prin crestele cupole , cum ar fi Corythosaurus , Hypacrosaurus și lambeozaur . De fapt, se pare că ruda sa cea mai apropiată este Charonosaurus , un lambeosaur cu un craniu similar (deși creasta craniană nu este atât de pronunțată), din regiunea Amur , din nordul Chinei , cu care formează clada Parasaurolophini . Specia P. cyrtocristatus , cu creasta sa rotunjită scurtă, pare a fi cea mai primitivă dintre cele trei specii, deși unii consideră că această specie este exemplare sub-adulte sau femele din specia P. tubicen .
Următoarea cladogramă expune subfamilia lambeosaurine, după redescrierea Lambeosaurus magnicristatus (Evans & Reisz, 2007):
| Hadrosauridae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Istoria descoperirii

Numele de PARASAUROLOPHUS înseamnă „lângă șopârla cu creastă“, deriva din greaca veche cu para / παρα care înseamnă „lângă“ sau „aproape“, saurus / σαυρος sau „șopârlă“ și lophos / λοφος sau „creasta“. Specia de tip P. walkeri se bazează pe exemplarul ROM 768, format dintr-un craniu și un schelet parțial care lipsesc cea mai mare parte a cozii și picioarelor din spate sub genunchi, găsit de o expediție de la Universitatea din Toronto în 1920, lângă Sand Creek de-a lungul râul Red Deer din Alberta , Canada . Aceste roci sunt acum cunoscute sub numele de Formația Parcului dinozaurilor, datând din Cretaceul Campanian , superior . William Parks a numit noua specie Parasaurolophus walkeri în onoarea lui Sir Byron Edmund Walker, președintele consiliului de administrație al Muzeului Regal Ontario. Rămășițele de Parasaurolophus sunt rare în Alberta, iar singurele artefacte găsite după holotip sunt un craniu parțial, probabil din formațiunea Dinosaur Park și trei exemplare fără sculele care ar putea aparține genului, din aceeași localitate. În unele liste faunistice, se menționează posibila descoperire a materialelor P. walkeri în formațiunea Hell Creek din Montana , o formațiune stâncă datând de la sfârșitul Maastrichtianului . [3]
În 1921, Charles H. Sternberg a recuperat un craniu parțial ( PMU. R1250) din ceea ce este acum cunoscut sub numele de Formația Kirtland din județul San Juan , New Mexico . Acest exemplar a fost trimis la Uppsala, Suedia, unde Carl Wiman l-a descris ca a doua specie de Parasaurolophus , adică P. tubicen , în 1931. Epitetul specific, tubicen derivă din latinescul tǔbǐcěn sau „trompetist”. Un al doilea craniu aproape complet de P. tubicen (NMMNH P-25100) a fost găsit în New Mexico în 1995. Folosind tomografia computerizată a craniului, Robert Sullivan și Thomas Williamson au scris o monografie a noii specii în 1999, care acoperă aspecte ale anatomia și taxonomia acesteia și funcțiile posibile ale creastei. Williamson a publicat ulterior o revizuire independentă a rămășițelor, în dezacord cu concluziile taxonomice.
John Ostrom a descris un alt exemplar ( FMNH P27393) în stare bună găsit în New Mexico, denumindu-l P. cyrtocristatus , în 1961. Acesta a inclus un craniu parțial, cu o creastă scurtă rotunjită și o mare parte a scheletului postcranian , cu excepția picioarelor. gâtul și unele părți ale cozii. Numele său specific derivă din latinescul curtus care înseamnă „scurtat” și cristatus care înseamnă „crestat”. Specimenul a fost găsit în ambele părți superioare ale formațiunii Fruitland sau, mai probabil, la baza formațiunii Kirtland aflate deasupra. Gama acestei specii a fost extinsă în 1979 când David B. Weishampel și James A. Jensen au descris un craniu parțial, cu o creastă similară ( BYU 2467) din formațiunea Kaiparowits din județul Garfield , Utah . Mai târziu, un alt craniu a fost găsit în Utah, cu o creastă scurtă și rotundă, ca cea a lui P. cyrtocristatus .
Specii
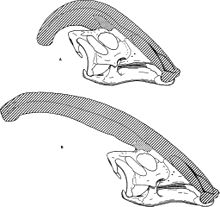
Genul Parasaurolophus are în principal trei specii : P. walkeri , P. tubicen și P. cyrtocristatus . Toate pot fi ușor distinse între ele, cu multe caracteristici diferite. Specia tip este P. walkeri , cunoscută din unele exemplare din Formația Parcului dinozaurilor. Se distinge de celelalte două specii prin creasta sa tubulară, mai scurtă și mai simplă intern decât cea a P. tubicen , dar mai mare și mai dreaptă decât cea a P. cyrtocristatus .

A doua specie, P. tubicen este cea mai mare din genul Parasaurolophus . A trăit în New Mexico și este cunoscut pentru trei exemplare și poate fi ușor recunoscut de alte specii datorită creastei sale lungi și drepte, cu o structură foarte complexă în interior comparativ cu celelalte specii. Toate exemplarele cunoscute de P. tubicen provin de la membrul De-Na-Zin al formațiunii Kirtland.
În 1961, John Ostrom a numit o a treia specie, P. cyrtocristatus . Cele trei exemplare cunoscute au fost găsite în formațiunile Fruitland și Kaiparowits, Utah și New Mexico. Al doilea exemplar, primul cunoscut din formațiunea Kaiparowits, a fost atribuit inițial unui taxon specific. Dintre speciile PARASAUROLOPHUS, P. cyrtocristatus este cele mai mici specii, si are o creasta curbată mai mică. Datorită acestei caracteristici neobișnuite, s-a speculat adesea că, în loc de o specie distinctă, ar putea fi de fapt femela P. walkeri sau P. tubicen , care erau masculi, chiar dacă P. tubicen a trăit aproximativ un milion de ani mai târziu. După cum a remarcat Thomas Williamson, materialul asemănător P. cyrtocristatus are aproximativ 72% dimensiunea P. tubicen , adică aproape de dimensiunea la care alți lambeosaurini sunt interpretați ca o demonstrație a dimorfismului sexual în creste (~ 70% dimensiunea adultului) . Deși mulți oameni de știință au susținut posibilitatea ca P. cyrtocristatus să fie pur și simplu exemplare de sex feminin, multe alte studii au arătat altfel din cauza diferențelor de vârstă, distribuție și diferențe mari în creastă și structura sa internă.
Un studiu publicat în PLoS ONE în 2014 a arătat că ar putea exista o altă specie de Parasaurolophus . Acest studiu, condus de Xing, a dovedit că Charonosaurus jiayinensis ar putea fi într-adevăr o nouă specie de Parasaurolophus , creând noua specie P. jiayensis . Dacă acest lucru ar fi adevărat, ar prelungi distribuția Parasaurolophus până la sfârșitul Maastrichtianului , observându-și prezența și în Asia .
Paleobiologie
Dietă
La fel ca toate hadrosauridele, Parasaurolophus a fost un erbivor terestru mare capabil să folosească atât locomotia bipedă, cât și cvadrupedală , capabilă să se hrănească cu un număr mare de plante datorită craniului sofisticat care i-a permis să efectueze o mișcare de rectificare similară cu cea de mestecat . Dinții lui erau înlocuiți constant; și au fost aranjate în baterii dentare care ar putea conține sute, dintre care doar o mână au fost utilizate perpetuu. Animalul și-a folosit ciocul pentru a tăia vegetația, apoi l-a împins spre dinți într-un organ asemănător obrajilor , unde a fost tăiat și înghițit. Animalul se putea hrăni cu vegetație care atingea până la 4 metri (13 picioare) deasupra solului. Bob Bakker a remarcat faptul că lambeosaurinele au ciocuri mai înguste și mai mici decât hadrosaurinele, ceea ce înseamnă că Parasaurolophus și rudele sale cele mai apropiate erau mai specializate în dietele lor decât verii lor cu factura largă, dar fără crest.
Creştere

Parasaurolophus este cunoscut pentru multe exemplare adulte și pentru un singur exemplar tânăr descris în 2013 . Scheletul tânărului animal a fost găsit în Formația Kaiparowits în 2009 datorită unei expediții comune de la The Webb Schools și Muzeul de Paleontologie (RAM) Raymond M. Alf. Conform descrierii, animalul avea doar un an când a murit și măsura 2,50 metri (8,2 picioare) în lungime. Referit la Parasaurolophus sp. , acest exemplar tânăr a fost clasificat ca RAM 14000 și este cea mai completă, precum și cea mai tânără, fosilă de Parasaurolophus găsită vreodată. În plus, datând de acum aproximativ 75 de milioane de ani, se potrivește perfect etapelor de creștere cunoscute în prezent ale Parasaurolophus . Deși nu sunt cunoscute cranii intermediare între RAM 14000 și exemplarele adulte Parasaurolophus , s-a găsit un craniu parțial care ar indica un individ la jumătatea distanței dintre tineri și adulți. La 25% din dimensiunea totală a adulților, această fosilă arată că creșterea crestei în Parasaurolophus a început mult mai devreme decât în genurile înrudite, cum ar fi Corythosaurus . Vârsta eșantionului și dimensiunea sa indică faptul că creșterea acestor animale a fost foarte rapidă. Creasta acestui exemplar nu a fost lungă și tubulară ca la adulți, ci joasă și emisferică.
Craniul RAM 14000 este aproape complet, din partea stângă lipsind doar o bucată din maxilarul superior . Cu toate acestea, craniul a fost împărțit în jumătate de eroziune , probabil când s-a așezat pe fundul albiei râului unde s-a fosilizat. Cele două părți au fost ușor deplasate, unele oase din dreapta fiind deplasate din blocul principal, tot prin eroziune. După reconstrucția scheletului, paleontologii au observat că craniul, văzut din lateral, seamănă mult cu cel al altor tineri lambeosaurini, având aproximativ forma unui trapez .
O piesă craniană parțială de RAM 14000 a fost reconstruită folosind o scanare CT , primul caz pentru un Parasaurolophus de orice stadiu ontogenetic. Distribuția a fost reconstruită în două secțiuni, o porțiune a craniului articulată cu jumătatea stângă a craniului și partea rămasă pe porțiunea dezarticulată a craniului. Poziția lor relativă a fost apoi aproximată pe baza reperelor craniene și a comparației cu alte hadrosauride. Datorită intemperiilor, multe dintre canalele neuronale mai mici și diferite foramine osoase nu pot fi identificate cu certitudine.
Creasta craniană

De la descoperirea sa, s-au avansat nenumărate ipoteze cu privire la funcțiile creastei craniene a Parasaurolophus , dar cele mai multe dintre ele au fost discreditate. Astăzi se crede că creasta ar fi putut avea mai multe funcții: de la vizualizarea intraspecifică la amplificarea sunetului pentru a comunica la termoreglare .
Diferențele dintre creste
În ceea ce privește celelalte lambeosaurine , se credea că creasta craniană a Parasaurolophus se schimbă odată cu vârsta și pe baza dimorfismului sexual prezent la adulți. James Hopson, unul dintre primii cercetători care a descris creastele lambeosaurine în termeni de astfel de distincții, a sugerat că specia P. cyrtocristatus , cu creasta mică rotunjită, reprezenta de fapt forma feminină a P. tubicen , în timp ce Thomas Williamson a sugerat că este formă tânără. Niciuna dintre aceste două ipoteze nu a fost vreodată acceptată pe scară largă, dar numai descoperirea altor cranii complete și a exemplarelor tinere, pe lângă cea deja găsită, va putea clarifica aceste relații potențiale. Williamson a observat, totuși, că tinerii Parasaurolophus aveau probabil o creastă mică, rotunjită ca P. cyrtocristatus , care probabil a devenit mai lungă și mai dreaptă pe măsură ce animalul a atins maturitatea sexuală . Reexaminarea recentă a unui craniu atribuit anterior unui tânăr Lambeosaurus , atribuit acum unui tânăr Parasaurolophus , oferă dovezi că, chiar și la exemplarele adolescente, creasta era tubulară. Acest exemplar păstrează o mică flăcare ascendentă în oasele frontale similară, dar mai mică, cu ceea ce se vede la exemplarele adulte; la adulți, frontalele formau o platformă care susținea baza creastei. Acest specimen indică, de asemenea, că creșterea creastei în Parasaurolophus și profilul facial al puilor diferă de modelul de creștere al Corythosaurus , Hypacrosaurus și Lambeosaurus , în parte deoarece creasta Parasaurolophus nu are osul subțire care alcătuiește vârful creastei celor trei. lambeosaurinele.
Ipoteze aruncate

Multe dintre primele ipoteze expuse despre funcția creastei se refereau la un presupus stil de viață acvatic, propus în acel moment pentru toate hadrosauridele, o linie de gândire comună până în 1960. Astfel, Alfred Sherwood Romer a propus ca creasta lungă să servească drept snorkel , în timp ce Martin Wilfarth susținea că creasta era un atașament pentru o proboscidă mobilă care ajuta animalul să respire în timp ce era scufundat în apă, pășunând plante marine. În schimb, Charles M. Sternberg a emis ipoteza că creasta a servit la menținerea apei din plămâni și Ned Colbert a susținut că creasta era un rezervor de aer pentru scufundări prelungite.
Alte ipoteze erau de natură mai fizică. După cum sa menționat anterior, William Parks a sugerat ca creasta să fie unită la vertebre prin ligamente și / sau mușchi, ajutând la mișcare și susținerea capului. Othenio Abel a propus ca creasta să fie folosită ca armă în luptele dintre membrii aceleiași specii, iar Andrew Milner a sugerat că avea funcția de a muta frunzele în timp ce animalul se deplasa prin vegetația densă, cum ar fi creasta osoasă a modernului. casuar . Alte propuneri au ipotezat prezența unor organe speciale: Halszka Osmólska a sugerat că creasta adăpostea glandele sărate, iar John Ostrom a ipotezat că creasta adăpostea zone mari de țesut olfactiv îmbunătățind exponențial simțul mirosului lambeosaurinelor, care nu aveau capacități defensive evidente. O sugestie oarecum neobișnuită și fantezistă a fost făcută de creaționistul Duane Gish , conform căruia creasta adăpostea glande chimice care ar permite lansarea jeturilor de substanțe chimice „înflăcărate” asupra inamicilor, similar cu gândacul bombardier modern.
Majoritatea acestor ipoteze au fost discreditate sau respinse. De exemplu, nu există o gaură la capătul creastei pentru o caracteristică de snorkeling. Nu există cicatrici musculare pentru o proboscidă și este îndoielnic dacă un animal cu cioc ar avea nevoie de o proboscidă. Chiar și utilizarea acestuia ca dispozitiv de blocare a aerului propus nu ar fi probabil, deoarece nu ar ține apa. Utilizarea acestuia ca rezervor de aer ar fi insuficientă pentru un animal de mărimea parasaurolofului . Alte hadrosauride aveau capete mari, fără a avea nevoie de creste mari, care să servească drept puncte de atașare pentru susținerea ligamentelor. Mai mult, niciuna dintre propuneri nu explică de ce creasta a fost astfel formată, iar alte lambeosaurini au posedat creaste de diferite forme, dar au îndeplinit funcții similare. Mai mult, absența cavităților sau absența totală a crestelor la alte hadrosauride nu ar explica locația glandelor sărate sau a sistemelor olfactive dezvoltate, indicând că acestea nu erau funcții primare ale creastei. Mai mult, studiul cavităților nazale ale lambeosaurinelor arată că nervii olfactivi și țesuturile senzoriale corespunzătoare se aflau în mare parte în afara porțiunii pasajelor nazale ale creastei, astfel încât expansiunea creastei a avut puțin de-a face cu simțul mirosului.
Termoreglare
Suprafața mare și vascularizația creastei sugerează o funcție de termoreglare. Primul care a propus o funcție similară cu crestele lambeosaurinelor a fost Wheeler (1978). Wheeler a propus că există o conexiune nervoasă între creastă și creier, astfel încât acesta din urmă să fie răcit de primul. Ipoteza a găsit consensul lui Maryanska și Osmólska, care și-au dat seama că, la fel ca șopârlele moderne, dinozaurii pot avea glande sărate și se pot răcori în afara osmo-reglării. În 2006, Evans a publicat o discuție despre funcțiile crestelor lambeosaurine, susținând că acesta ar putea fi unul dintre factorii din spatele evoluției creastei.
Comunicare

Una dintre cele mai acreditate ipoteze cu privire la utilizarea creastei Parasaurolophus este utilizarea acestei structuri ca o cameră de rezonanță pentru a produce sunete de joasă frecvență pentru a alerta alți membri ai unui grup sau ai speciilor sale. Această caracteristică a fost sugerată inițial de Wiman în 1931 când a descris P. tubicen . El a observat că structura internă a crestelor era similară cu cea a unei lebede și a teoretizat că animalul a folosit aceste pasaje nazale alungite pentru a crea zgomote de joasă frecvență. Cu toate acestea, tuburile nazale ale Hypacrosaurus , Corythosaurus și Lambeosaurus sunt mult mai variabile și mai complicate decât căile respiratorii ale Parasaurolophus . O cantitate mare de materiale și date susține ipoteza că creasta tubulară mare a Parasaurolophus a fost o cameră de rezonanță. Weishampel în 1981 a sugerat că Parasaurolophus ar putea emite sunete care variază în frecvență de la 55 la 720 Hz , deși aceste valori variau probabil de la specie la specie, datorită formei și dimensiunii creastei, mai evidentă la P. cyrtocristatus . Hopson a descoperit că există dovezi anatomice că hadrosauridele au auzit excelent. Există cel puțin un exemplu, în coritosaurul înrudit, al unui etrier subțire (osul urechii reptilei) care combinat cu un spațiu mare pentru un timpan indică prezența unei urechi foarte sensibile. Mai mult, lagrena hadrosauridă a fost alungită ca cea a unui crocodil , indicând faptul că porțiunea auditivă a urechii interne a fost bine dezvoltată. Pe baza asemănărilor dintre urechea internă a hadrosauridelor și cea a crocodililor, s-a propus că hadrosauridele adulte erau sensibile la frecvențe ridicate, precum cele pe care descendenții lor le-ar putea emite. Potrivit lui Weishampel, acest lucru este în concordanță cu o comunicare între părinți și copii, indicând, de asemenea, un posibil comportament de îngrijire a părinților.
Reconstrucția unui craniu excepțional bine conservat al unui exemplar de P. tubicen , cu pasaje de aer mai complexe decât cele ale lui P. walkeri , a permis reconstrucția posibilului sunet emis de creasta animalului. Calea principală rezonează la aproximativ 30 Hz, dar anatomia complicată a sinusului provoacă vârfuri și văi de sunet. Cealaltă teorie comportamentală principală este că creasta a fost utilizată pentru recunoaștere în cadrul indivizilor din aceeași specie. Aceasta înseamnă că creasta ar fi putut fi utilizată pentru recunoașterea speciilor, ca semnal de avertizare și pentru alte utilizări non-sexuale. Acestea sunt doar câteva exemple ale evoluției crestelor Parasaurolophus și ale altor hadrosauride. În schimb, funcțiile sociale și fiziologice sunt mai susținute ca funcții de creastă, concentrându-se pe identificarea vizuală și auditivă . Dimensiunea mare a ochiului hadrosaurids' prize și prezența unui inel sclerotic în ochi implica viziune acuta si obiceiuri diurn, dovezi care arată că viziunea a fost importantă pentru aceste animale. Dacă creasta ar fi fost cu adevărat atașată de ceafă printr-o membrană a pielii (așa cum este ilustrat în mod obișnuit), afișajul vizual propus ar fi fost și mai izbitor. După cum a sugerat alte cranii lambeosaurine, creasta PARASAUROLOPHUS permis probabil atât identificarea genului ( de exemplu il separa Corythosaurus sau lambeozaur) și identificarea sexuală prin formă și dimensiune.
Paleopatologie
Există un specimen de P. walkeri care ar arăta dovezi ale unei patologii . Scheletul prezintă o fantă sau o crestătură în formă de V în vertebrele de la baza gâtului. Inițial, considerat patologic, Parks a publicat o a doua interpretare, ca alocare pentru un ligament care susținea capul și care ar fi legat de crestătură prin mușchi sau ligamente. O altă posibilitate este că, în timpul pregătirii, specimenul a fost deteriorat, totuși este mai probabil să credem că această crestătură este un semn al unei patologii.
O altă posibilă patologie a fost observată de Parks, în timpul studiului crestăturii de pe vertebra cervicală. La a patra, a cincea și a șasea vertebră cervicală, direct anterior crestăturii, coloanele vertebrale neuronale au fost deteriorate. Al patrulea a avut o fractură evidentă, iar ceilalți doi au avut o umflătură la baza pauzei.
Paleoecologie
Alberta
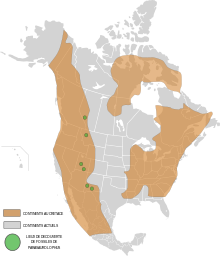
Parasaurolophus walkeri , din formațiunea Dinosaur Parck, a fost membru al unei faune diverse și bine documentate de animale preistorice, incluzând mai mulți dinozauri cunoscuți, precum ceratopsidele Centrosaurus , Chasmosaurus și Styracosaurus ; alte hadrosauride precum Gryposaurus și Corythosaurus ; tyrannosaurid Gorgosaurus și ankylosaurids Edmontonia , euoplocephalus și Dyoplosaurus . Cu toate acestea, Parasaurolophus este un membru destul de rar al acestei faune. La Formazione Dinosaur Parck è stata interpretata una pianura alluvionale a livello del mare striata di numerosi fiumi, divenendo più paludosa e influenzata dalle condizioni marine nel corso del tempo, quando il mare interno occidentale si spostò verso ovest. Il clima era più caldo di oggi, prevenendo la formazione di ghiacci, diviso in stagioni umide e secche . Le conifere erano le piante dominanti e nelle foreste formavano un baldacchino di piante, con un sottobosco di felci, felci arboree e angiosperme .
Gli hadrosauri meno comuni della Formazione Dinosaur Park, del Dinosaur Provincial Park, come ad esempio il Parasaurolophus , potrebbero rappresentare i resti di individui che sono morti durante le migrazioni attraverso la regione. È possibile che questi animali vivessero, nidificassero e si nutrissero sugli altopiani dove i processi di fossilizzazione sono più rari. La presenza di Parasaurolophus e Kritosaurus in siti fossili di latitudine nord potrebbe rappresentare uno scambio faunistico tra il nord e il sud, altrimenti distinti durante il Cretaceo superiore del Nord America . Entrambi i taxa sono rari al di fuori del bioma meridionale dove, insieme al Pentaceratops , rappresentano i membri dominanti della fauna del luogo.
Nuovo Messico

Nella formazione Fruitland del Nuovo Messico , il P. cyrtocristatus condivideva il suo habitat con altri ornitischi e teropodi . Specificamente, i suoi contemporanei erano il ceratopside Pentaceratops e il pachycephalosauro Stegoceras . Nella formazione inoltre sono stati ritrovati alcuni fossili identificati come appartenenti a tirannosauridi , Ornithomimus , troodontidi , Saurornitholestes , Struthiomimus , ornitopodi , Chasmosaurus , Corythosaurus , hadrosaurini , hadrosauridi e ceratopsidi . All'epoca del Parasaurolophus , la Formazione Fruitland era paludosa, posizionato in una pianura, e vicino alle rive del mare interno occidentale . Questa formazione risale al Cretaceo superiore, circa 75.5-74.5 milioni di anni fa.
Nonostante sia più recente di P. cyrtocristatus , anche la specie P. tubicen visse in Nuovo Messico, all'interno della Formazione Kirtland. In questa formazione vissero numerosi vertebrati, tra cui pesci , crurotarsi , ornitischi , saurischi , pterosauri e tartarughe . I pesci sono rappresentati dai due specie, Melvius chauliodous e Myledalphus bipartitus . I crurotarsi includono il Brachychampsa montana e il Denazinosuchus kirtlandicus . Tra i dinosauri , gli ornitischi dalla formazione comprendevano gli hadrosauridi Anasazisaurus , Naashoibitosaurus , Kritosaurus e lo stesso P. tubicen , gli ankylosauridi Ahshislepelta e Nodocephalosaurus ; i ceratopsidi Pentaceratops e Titanoceratops , ei pachycephalosauri Stegoceras e Sphaerotholus . I dinosauri saurischi invece contano il tirannosauride Bistahieversor ; l' ornithomimide Ornithomimus , e il troodontide " Saurornitholestes " robustus . La formazione è nota anche per uno pterosauro conosciuto sotto il nome di Navajodactylus . Anche le tartarughe sono piuttosto abbondanti, e ne sono noti i generi Denazinemys , Basilemys , Neurankylus , Plastomenus e Thescelus . Altri taxa non identificati noti, comprendono il crurotarso? Leidyosuchus canadensis , ei teropodi? Struthiomimus , troodontidi e tirannosauridi. Questa formazione risale al Cretaceo superiore, circa 74.5-73.05 milioni di anni fa.
Utah
La datazione radiometrica argon-argon indica che la Formazione Kaiparowits si è depositata tra i 76,6 ei 74,5 milioni di anni fa, durante la fase Campaniano , del Cretaceo superiore . Durante il tardo periodo Cretaceo, il sito della Formazione Kaiparowits era situato vicino alla riva occidentale del Mare interno occidentale , un grande mare interno che diviso il Nord America in due masse distinte, Laramidia ad ovest e l' Appalachia ad est. L'altopiano su cui vivevano i dinosauri era un'antica pianura alluvionale dominata da grandi canali e zone umide abbondanti di torbe paludose, stagni e laghi, delimitata da altopiani. Il clima era umido, in grado di sostenere la ricca gamma di organismi che vivevano in questo ambiente. Questa formazione contiene una delle migliori e più continue registrazioni della vita terrestre del Cretaceo superiore, in tutto il mondo.
Il Parasaurolophus condivideva questo suo paleoambiente con altri dinosauri, come i teropodi dromaeosauridi , il troodontide Talos , l' ornithomimide Ornithomimus , i tirannosauridi Albertosaurus e Teratophoneus , alcuni ankylosauridi , l' hadrosauride Gryposaurus , i ceratopsidi Utahceratops , Nasutoceratops e Kosmoceratops , l' oviraptorosauro Hagryphus . La paleofauna presente nella Formazione Kaiparowits include anche pesci cartilaginei (squali e razze), rane , salamandre , tartarughe , lucertole e coccodrilli . Vi era anche una gran varietà di piccoli mammiferi , tra cui multituberculati , marsupiali e insettivori .
Nella cultura di massa
Il Parasaurolofo compare anche nella fortunata saga cinematografica di Jurassic Park . Nel primo film compare in branco (per pochi secondi) insieme ad un gruppo di Brachiosauri. Nel secondo film, Il mondo perduto - Jurassic Park , il Parasaurolofo compare maggiormente rispetto agli altri capitoli; un esemplare viene catturato dalla InGen sotto gli occhi della squadra di Ian Malcolm .
Nel film We're Back! - 4 dinosauri a New York ( We're Back! A Dinosaur's Story ) Goffardo, uno dei quattro dinosauri protagonisti, è un Parasaurolofo.
Diversi esemplari di Parasaurolofo appaiono in due classici Disney : Fantasia e Dinosauri . Questo dinosauro appare anche nel film Il viaggio di Arlo realizzato dalla Pixar Animation Studios . Il Parasaurolofo è presente anche nel videogioco Jurassic World Evolution . Fa diverse comparse nei film d'animazione di Alla ricerca della Valle Incantata
Alcune ricostruzioni di Parasaurolofo sono presenti in diversi parchi tematici a tema preistorico: al Parco Natura Viva di Bussolengo ( VR ), al parco Il Mondo della Preistoria a Simbario ( VV ) e al Parco dei Dinosauri di Castellana Grotte ( BA )
Galleria d'immagini


Note
- ^ Dinosauri,Le creature più terrificanti della terra , DIX, p. 139, ISBN 978-88-95870-05-2 .
- ^ Senter P, Robins JH (2015) Resting Orientations of Dinosaur Scapulae and Forelimbs: A Numerical Analysis, with Implications for Reconstructions and Museum Mounts . PLoS ONE 10(12): e0144036. doi:10.1371/journal.pone.0144036
- ^ Noted as being present, although misspelled as " Thescelosaurus garbani , in " "Dinosaur distribution (Late Cretaceous; North America; South Dakota)." Weishampel, et al. (2004). Page 586.
Bibliografia
- Benton MJ, John Sibbick J. Vertebrate Palaeontology . pp. 207–209, 2000
- Evans, David C. Nasal cavity homologies and cranial crest function in lambeosaurine dinosaurs . Paleobiology 32 (1): 109–125, 2006.
- Evans, David C.; Reisz, Robert R.; and Dupuis, Kevin. A juvenile Parasaurolophus braincase from Dinosaur Provincial Park, Alberta, with comments on crest ontogeny in the genus . Journal of Vertebrate Paleontology 27 (3): 642–650, 2007.
- Glut, Donald F. Parasaurolophus . Dinosaurs: The Encyclopedia. Jefferson, North Carolina: McFarland & Co. pp. 678–684, 1997.
- Ostrom JH Cranial morphology of the Hadrosaurian dinosaurs of North America . 122(2):33-186, 1961.
- Ostrom JH Parasaurolophus cyrtocristatus, a crested hadrosaurian dinosaur from New Mexico . Chicago Natural History Museum 1963, III ed.
- Sandia National Laboratories. Scientists Use Digital Paleontology to Produce Voice of Parasaurolophus Dinosaur , 12/05/1997.
- Sullivan, Robert M.; and Williamson, Thomas E. A new skull of Parasaurolophus (Dinosauria: Hadrosauridae) from the Kirtland Formation of New Mexico and a revision of the genus . New Mexico Museum of Natural History and Science Bulletin, 15. Albuqueque, New Mexico: New Mexico Museum of Natural History and Science. pp. 1–52, 1999.
- Weishampel, David B. and Jensen, James A. Parasaurolophus (Reptilia: Hadrosauridae) from Utah . Journal of Paleontology 53 (6): 1422–1427, 1979.
- Williamson, Thomas E. Review of Hadrosauridae (Dinosauria, Ornithischia) from the San Juan Basin, New Mexico . in Lucas, SG; and Heckert, AB (eds.). Dinosaurs of New Mexico. New Mexico Museum of Natural History and Science Bulletin, 17. Albuqueque, New Mexico: New Mexico Museum of Natural History and Science. pp. 191–213, 2000.
Altri progetti
-
 Wikimedia Commons contiene immagini o altri file su Parasaurolophus
Wikimedia Commons contiene immagini o altri file su Parasaurolophus -
 Wikispecies contiene informazioni su Parasaurolophus
Wikispecies contiene informazioni su Parasaurolophus
Collegamenti esterni
- ( EN ) Parasaurolophus , su Enciclopedia Britannica , Encyclopædia Britannica, Inc.
- ( EN ) Parasaurolophus , su Fossilworks.org .