Telencefalul
| Telencefalul | |
|---|---|
 | |
 | |
| Anatomia lui Gray | (RO) Pagina 817 |
| Nume latin | Cerebrum |
| Sistem | Sistem nervos central |
| Localizare anatomică | creierul anterior |
| Artera | anterioară a arterei cerebrale , cerebrale medii arterei și arterei cerebrale posterioare |
| Venă | venele cerebrale |
| Identificatori | |
| Plasă | A08.186.21.730.885 și A08.186.211.200.885.287 |
| TA | A14.1.03.008 și A14.1.09.001 |
| FMA | 62000 |
| TU | H3.11.03.6.00001 |
| TU | E5.14.1.0.2.0.12 |
| ID NeuroLex | birnlex_1042 |
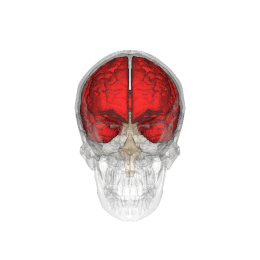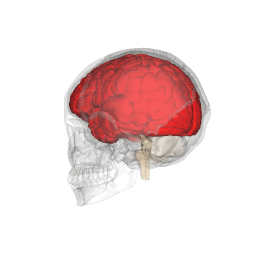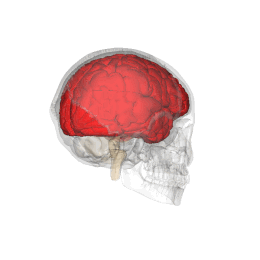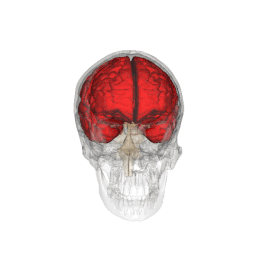
Telencephalon este partea din creier care are cea mai mare extindere la om . Împreună cu diencefalului aceasta formeaza creierul , organul principal al sistemului nervos central .
Telencephalon are o formă ovoidală, cu axa mare orientată anteroposteriorly, și este poziționat în interiorul craniului, în afara diencefalului . Acesta este împărțit în două formațiuni juxtapuse și aproape identice, numite emisfere; emisfera dreaptă primește senzațiile părții stângi a corpului, în timp ce opusul este valabil pentru cealaltă emisferă. [1]
Printre principalele sale funcții pot fi listate primirea de stimuli externi și prelucrarea unui răspuns cu motor, memorie , abilități de luare a deciziilor.
Aranjament și rapoarte
Telencephalon este poziționat în interiorul craniului și este separat de țesutul osos doar prin interpunerea trei subtiri meningeale straturi ( Dura mater , arachnoida , PIA Mater ) și, ocazional, de mici cisterne subarahnoidiană umplute cu lichior . Partea superioară sau dorsală, de suprafață a telencephalon este convexă și se potrivește pe suprafața interioară concavă a oaselor craniene, în timp ce sa inferioară, sau ventrale, suprafața este plană.
| Nomenclatură |
|---|
În descrierea relațiilor spațiale ale telencefalului este necesar să se țină cont de o mică variație semantică a termenilor folosiți în general în anatomie: în timp ce măduva spinării și toate celelalte formațiuni ale corpului, de fapt, se dezvoltă într-o direcție verticală, în creierul structurile anatomice sunt descrise în sens orizontal. Prin urmare, în anatomia telencefalului, unii termeni folosiți în mod obișnuit pentru întregul corp uman iau semnificații diferite:
|

Telencephalon este compus din două emisfere care sunt separate una de alta printr - o mare fisură interhemispheric , în care mare cerebral secere este poziționat, o sagital sepia a dura mater. Fiecare dintre cele două emisfere are trei fețe (medială, laterală, inferioară) și trei margini rotunjite (superioare, inferomediale, inferolaterale).
Fața medială a fiecărei emisfere este verticală și plană și se confruntă cu fața corespunzătoare a emisferei contralaterale, de care este separată prin interpunerea marii secere cerebrale. La marginea inferomedial, cele două fețe medial sunt puse în continuitate printr - o formațiune commessural, The calos .
Fața laterală a emisferelor este modelată pe suprafața concavă a oaselor craniului și, prin urmare, își asumă un aspect convex. Fața inferioară a fiecărei pauze emisfera de pe oasele bazei craniului și este împărțit de fisurii laterale în două porțiuni:
- anterioară, porțiunea frontală este mai mică decât cea posterioară și se sprijină pe cavităților orbitale excavate în osul frontal ;
- porțiunea posterioară, temporooccipitală, este considerabil extinsă și ia relații cu oasele care alcătuiesc baza craniană și, în regiunea sa cea mai posterioară, cu un sepiment meningeal într-o dispoziție orizontală, tentoriul cerebelului , care separă polul occipital a emisferelor telencefalice din cerebelul însuși.
Vascularizație
Derivarea embriologică
Cele derivă telencephalon din forebrain - veziculă, o ejecție de duș neuronale care se formează în jurul zilei 25 după fertilizare [2] . Aceasta se divide veziculă primitive, în jurul zilei de 32 fertilizare, în alte două vezicule: mai caudal este numit vezicula diencefalică, în timp ce mai rostral este numit vezicula telencephalic. Din acesta din urmă, prin diviziuni succesive, își au originea toate formațiunile telencefalului definitiv.
Structura

La un examen macroscopic, suprafața telencephalon umane (cortexul telencephalic sau pallium, a se vedea sub titlul Stratificarea ) apare neregulat datorită prezenței a numeroase brazde, care împreună delimitează un număr enorm de crestele numite convoluții. Această caracteristică, dezvoltat la cel mai înalt grad din specia umană, se numește gyrencephaly [3] , și dobândește o mare importanță pentru capacitatea sa de a crește în mod semnificativ suprafața telencephalic disponibile.
Printre diferitele brazde care se execută prin suprafața telencephalic, unele mai marcate și evident pot fi identificate: acestea sunt numite fisuri. Unele dintre aceste fisuri, aprofundarea în cortexul telencephalic, participa la împărțirea în lobi.
Fisuri principale
Fisurile care marchează granița dintre diferiții lobi sunt , în principal cinci [4] , și anume:
- Fisura central (de Rolando)
- despicătură laterală (Silvio)
- fisură parietoccipitală
- fisura calcarină
- Fisura limbic.

Central ( Rolando lui fisurii) începe de la marginea superioară a emisferei telencephalic, aproximativ spre mijlocul ei, și se execută de-a lungul întregii fețe laterale într - o direcție aproximativ verticală, pentru a opri la scurt timp după fisura laterală. Se împarte lobul frontal, anterior, de lobul parietal, posterior. Pe aspectul medial al emisferelor, fisurii centrală se adâncește în mijlocul unui pliu mare de trecere între cei doi lobi, lobulului paracentral.
Anterior și posterior la fisurii centrale există două spirele de mare importanță, respectiv atribuite funcției motorii și a funcției senzoriale: a precentral (sau prerolandic) girusul și postcentral (sau posttrolandic) girusul.
Fisura laterală (de Silvio ) separă lobul frontal înainte și spatele lobului parietal de lobul temporal, care este situat într - o poziție mai mică decât fisurii în sine. Mai mult sau mai puțin originare de la capătul inferior al unui plan frontal care separă lobul anterior al telencephalon în jumătate, fisura laterala merge spre polul posterior cu un curs ușor oblic în sus, și apoi furculițe într - un Y. După cum sa menționat deja, fisura laterală nu este atinsă de fisura centrală, care se oprește la mică distanță de ea.

Provine de mică adâncime parietoccipital fisurarea același punct ca și fisurii calcarine , pe peretele medial al emisferelor telencephalic, și merge în sus și oblic până la marginea superioară: de aici, se pornește de pe partea laterală, delimitând o graniță între parietal lob, anteriorly, și lobul occipital, posterior. Porțiunea fisurii care ruleaza pe fața medială a emisferelor telencephalic se numește fisurii perpendicular internă, iar cel care ruleaza pe fața laterală este numită fisurii perpendicular extern.
Fisura calcarine este singura dintre fisurile descrise aici , care nu identifică limita dintre două sau mai multe cortical lobi. Se desfășoară de-a lungul feței mediale a emisferelor telencefalice cu un curs aproape orizontal, de la confluența cu fisura perpendiculară internă până la marginea superioară, și apoi se scurge într-o scurtă întindere care traversează fața laterală a emisferelor. Primar vizual zona este amenajat în jurul fisurii calcarine, care este de o importanță fundamentală pentru mecanismul de vizibilitate.
Fisura limbică se găsește numai pe aspectul medial al emisferelor. Urmează cursul corpului calos: provenind de fapt inferior rostrului, descrie o curbă cu o concavitate mai mică și se termină în porțiunea posterioară a feței mediale, sub spleniul corpului calos. Fisura limbic imparte lobilor frontali si parietali, superioare, din lobul limbic, inferior.
În plus față de aceste cinci fisuri, numite fisuri primare, suprafața outcropping a cortexului telencephalic are alte caneluri, mai puțin importante, interne lobi. Dacă este necesar, acestea vor fi descrise în partea privind lobii individuali.
Lobi

Lobii în care este divizată cortexul telencephalic sunt șase [5] :

Acești lobi au o extensie aproximativ egală pe ambele emisfere telencefalice: uneori, totuși, ele pot avea circumvoluții speciale pe o singură emisferă și nu pe cealaltă. Această caracteristică explică conceptul de dominanță emisferică, adică principiul care atribuie unele dintre cele mai importante funcții ale vieții unei singure emisfere (în general stânga, chiar dacă 30% din populație nu pare să prezinte diferențe substanțiale între cele două emisfere) .de relație, cum ar fi limbajul . Acest concept va fi explorate în continuare în funcțiile secțiunii.
Stratificare

Telencefalul este împărțit în trei straturi care se succed de la exterior la interior și care sunt respectiv:
- cortexul cerebral sau paliumului
- substanța albă a telencefalului
- nucleii bazei.
Cortex cerebral
Arhitectura generală a cortexului telencephalic (sau pallium) a fost deja descrisă în Lobi secțiunea. Pentru a completa descrierea de la un precum și un punct anatomic de vedere funcțional, este necesar să se adauge o referire la principala strategie de localizare topografic a diferitelor zone funcționale ale cortexului, care este clasificarea Brodmann, precum și la arhitectura celulară cortexul in sine sau cytoarchitectonic.
Clasificarea Brodmann
Conform clasificării Brodmann [6] , pe suprafața cortexului telencephalic este posibil să se identifice 52 de zone (numite pentru aceasta zone Brodmann ) cu diferite cytoarchitectonic caracteristici și funcționale. O reprezentare aproximativă a acestora poate fi văzută în figura din stânga.

Zona somatic primară (în verde), care primește impulsuri ale sensibilității generale somatice a întregului corp, se află în girusul postcentral și precis în zonele 3, 1 și 2 din lobul parietal . În aceste zone este posibil să se găsească o reprezentare somatotopică a tuturor diferitelor părți ale corpului uman, care ia numele de homuncul senzorial : aceasta este o hartă reală a teritoriilor corticale care sunt excitate de stimulii care vin dintr-o anumită zonă a pielii . Termenul "homunculus" se datorează faptului că reprezentarea corpului uman pare grotesc și disproporționat: acest lucru se întâmplă deoarece fiecare parte a corpului este reprezentată în homunculus nu pe baza extinderii sale spațiale reale, ci pe baza numărul de receptori care sunt concentrați în el. Consecința acestui principiu este că zonele corticale responsabile de primirea stimulilor de pe față, buze și mână sunt nefiresc de mari, în timp ce cele atribuite trunchiului și picioarelor par mult mai mici decât extensia reală a corpului acestor teritorii.
Zona cu motor primar (sau motor) (în roșu), situat pe girusul precentral din lobul frontal , este format din zona 4. Acest teritoriu corticală este atribuit funcția de a genera impulsuri cu motor pentru toți mușchii corpului: de aceea, în ea există un homunculus cu motor analog cu cel sensibil, deși cu unele diferențe minime. În homunculul motor, de fapt, teritoriile dedicate inervației mușchilor mimici ai feței și ai mușchilor mâinii ocupă o mare măsură, în timp ce trunchiul este reprezentat minim. Anterior acestei zone este zona premotorului (sau premotorului) sau zona motorului secundar (zona 6). Adiacentă celei din urmă se află zona motorie suplimentară (AMS sau SMA), situată în poziția superomedială a emisferei cerebrale, în lobul frontal. Se invecineaza cu aspectul medial până la girusul cingului . În AMS există o reprezentare somatotopică a corpului. Acesta primeste circuite din nucleii telencephalic bazei . Se schimbă multe fibre cu contralateral prin corpul calos , este unul dintre cei cu mai multe legături cu cealaltă emisferă. Primește fibre subcorticale din lobul parietal posterior, din zona motorie parietală superioară. Ea dă efferences cortico (anterioară căii piramidale, aproximativ 30.000 de fibre) direct și fibre la cortexul motor . În caz de vătămare, apare apraxia severă cu pierderea coordonării bimanuale a membrelor. AMS reglează tonusul muscular în mișcări manuale fine, are memorie motorie complexă în gesturi și comportamente.
Zona vizual primar (în albastru) ocupă poziția 17 în zona de clasificare Brodmann, pe lobul occipital , și reprezintă zona corticală la care aferenților provin din retinei ajunge. Alături, în direcția mediolaterală, se pot observa zona vizuală secundară (zona 18) și zona vizuală terțiară (zona 19).
Zona acustică primară (sau auditiv) (în violet) este situat pe laterala aspectului al lobului temporal și corespunde Brodmann e zona 22 . Cele mai multe dintre aferenților ale căii acustice centrale a ajunge la ea. În direcția laterală continuă cu zona (zona) acustică (sau auditivă) secundară.
Zona gustativă (în galben), identificat în zona 43 a lobului insula , este considerat unul dintre centrele la care stimulii gustativ senzitivitate se referă . Mai mult, nu este centrul principal, deoarece s-a demonstrat că majoritatea stimulilor de natură gustativă ajung în zona somestetică primară.
Zona primară de vorbire sau zona lui Broca (în portocaliu) este o extensie a aerului motor primar care se ocupă exclusiv de inervația mușchilor de fonație. A fost identificat în zonele 44 și 45 ale lobului frontal.
Folii
Neocortexul, în urma dezvoltării tehnicilor de colorare precum metoda Golgi, Cajal, Nissl sau Weigert, a fost împărțit în șase lamine sau straturi paralele cu suprafața corticală cu caracteristici specifice. De la cele mai superficiale până la cele mai profunde sunt: lamina I (strat molecular), lamina II (stratul granular exterior), lamina III (stratul piramidal), lamina IV (stratul interior granular), lamina V (stratul ganglionar) și lamina VI (multiform strat).
- I - Stratul molecular sau plexiform este cel mai superficial și cel mai sărac dintre corpurile celulare. Conține corpuri celulare aproape exclusiv orizontale cu dendritele lor lungi, dendrite celulare piramidale, ramificațiile axonului celulelor Martinotti, fibre aferente din neuronii subcorticali. Dendritele și axonii celulelor orizontale au un curs orizontal, în timp ce cele ale celulelor piramidale și ale Martinotti au un curs vertical cu ramuri colaterale transversale sau orizontale. În general, însă, fibrele orizontale sunt predominante și formează un plex dens în centrul acestui strat. Este un strat pe care îl ating în principal fibrele aferente.
- II - Stratul granular exterior este un strat mai subțire decât cel molecular și este cel mai subțire după stratul granular interior. Conține corpurile celulelor mici piramidale, celulelor neurogliaforme, dendritelor celulelor nepiramidale spinoase, dendritelor celulelor orizontale (dar într-o măsură mult mai mică decât stratul molecular), axonii ascendenți ai Martinotti și celulele cu buchet dublu, dendritele de celule fusiforme sau bipolare. Fibrele sunt în principal verticale și sunt mai puțin dense decât stratul molecular. Acest strat dă naștere în special fibrelor asociative corticocorticale scurte.
- III - Stratul piramidal sau piramidal exterior este un strat foarte gros, în care celulele piramidale de fiecare tip sunt neuronii cei mai reprezentați. Acest strat conține, de asemenea, corpurile celulare ale multor celule spinoase nepiramidale, unele celule neurogliaforme și fusiforme, celule coș, axoni ascendenți ai celulelor Martinotti și dendrite ale celor duble buchet. Fibrele sunt în principal verticale și se îngroașă mai profund, apropiindu-se de stratul granular intern. Acest strat are subdiviziunile IIIa, IIIb și IIIc. Acest strat dă naștere în principal la fibre lungi de asociere corticocorticală.
- IV - Stratul granular interior este cel mai subțire strat al cortexului cerebral. Conține corpurile celulare ale celulelor mici piramidale și celulelor cu buchet dublu, celulelor nepiramidale spinoase, precum și fibrelor din alte celule piramidale, din axonii celulelor Martinotti, în cele din urmă corpuri și axoni ai celulelor coșului. Fibrele sunt foarte dense și orizontale, iar îngroșarea din interior se numește banda exterioară Baillarger. Este stratul care primește cel mai mare număr de fibre aferente.
- V - ganglionul interior sau stratul piramidal este un strat gros în care sunt cuprinse corpurile celor mai mari celule piramidale ale cortexului cerebral. De asemenea, conține corpuri celulare neurogliaforme și Martinotti și dendrite celulare cu buchet dublu. Fibrele orizontale foarte dense formează banda interioară Baillarger, dar este un strat bogat de fibre ascendente și descendente. Dă naștere la fibre proiective eferente corticostriate, corticopontin, corticobulbar și fibre corticospinale, dar și fibre asociative corticocorticale și fibre corticotalamice.
- VI - Stratul multiform este cel mai gros și mai adânc strat al cortexului cerebral continuu în stările sale cele mai profunde cu substanța albă fără o delimitare clară. Se numește așa datorită eterogenității mari a celulelor prezente în interiorul cărora, totuși, tind să aibă un pirenofor mediu sau mic. Conține foarte puține celule piramidale, dar multe celule Martinotti și neurogliaforme, precum și multe dendrite ale celulelor piramidale ale stratului de ganglion suprapus și ale celulelor în formă de panou și fus. Cursul fibrelor este variat, dar este cel mai bogat strat de fibre nervoase din cortexul cerebral (și cel mai bogat în fibre mielinizate), în special în stările sale cele mai profunde datorită apropierii de substanța albă. Din acest strat provine majoritatea fibrelor corticotalamice, dar primește și un anumit contingent de fibre aferente.
Variante regionale
Neocortexul are cinci variante regionale numite agranulare, frontale, parietale, granulare și polare. Variantele agranulare și granulare ale neocortexului conțin doar cinci dintre cele șase lamine distincte din celelalte trei tipuri de neocortex sau conțin șase, dar puțin dezvoltate. Pe baza prezenței tuturor laminelor sau nu, se disting astfel corticile homotipice (frontale, parietale și polare), care conțin toate lamele și corticile heterotipice (agranulare și granulare) unde nu sunt toate bine reprezentate.
- Cortexul agranular are lamine II slab dezvoltate (stratul granular exterior) și IV (stratul granular interior), totuși lamina V (stratul piramidal) conține mai multe celule piramidale mari decât alte tipuri de cortex. Este răspândit în zonele motorii ale cortexului, cum ar fi sediul girului precentral (zona 4) a cortexului motor primar, cortexul premotor (zona 6), dar și girusul paraolfactiv sau zona subcalosală, parte a sistemului limbic. Este cel mai gros tip de scoarță.
- Cortexul granular are un strat granular extern deosebit de dezvoltat, cu numeroase celule spinoase nepiramidale în interior și celule mici piramidale. Lamina III (stratul piramidal) este slab dezvoltată. Este un tip de cortex asociat cu zone la care ajung multe fibre aferente, de fapt compune cortexul somatosenzorial primar (girul postcentral), girul temporal superior (o zonă cu aferențe acustice), zona striată (aferente vizuale). După cortexul polar, este cel mai subțire cortex.
- Cortexul frontal are lamine II și IV slab dezvoltate, asemănătoare cu cortexul agranular, dar în lamina sa III (stratul piramidal) și V (stratul ganglionar) există foarte multe celule piramidale de dimensiuni medii-mici. Este cea mai groasă scoarță după cea agranulară. Este răspândit în majoritatea lobului frontal, cu excepția zonelor 10 și 46 și pentru girusul rectus, în lobul parietal din zona 7 și în girusul temporal inferior al lobului temporal.
- Cortexul parietal are lamine II și IV mai dezvoltate decât frontalul, similar cu acesta are un număr mare de celule piramidale de dimensiuni mici sau medii și posedă un număr mare de celule stelate. Împreună cu frontalul, este cel mai frecvent tip de cortex din telencefal. Acesta include girul unghiular, girul supramarginal, girul temporal mediu și superior și zonele 10 și 46 ale lobului frontal. Coaja sa are o grosime intermediară între cele cinci tipuri.
- Cortexul polar are o lamina subțire III (strat piramidal) în comparație cu alte tipuri de cortex, are un număr bun de celule stelate, dar într-o măsură mai mică decât stratul granular. Este cel mai subțire și cel mai puțin reprezentat tip de scoarță. Include cortexul cel mai apropiat de polii frontali și occipitali și zona parastriată sau zona vizuală secundară (zona 18).
Citoarhitectonică
Cortexul telencephalic poate fi filogenetic împărțit în trei porțiuni, în funcție de ordinea apariției în evoluția animalelor: de la cel mai vechi la cel mai recent, aceste porțiuni sunt archipallium, paleopallium și neopallium [7] . Archipallium și paleopallium, antecedente filogenetic și, prin urmare, prezente și la animalele mai puțin evoluate, au un citoarhitectonic relativ simplu și uniform, care ia denumirea de alocortex sau cortex heterogenetic; pe de altă parte, cortexul neopallium, care este apanajul animalelor superioare și, prin urmare, mai complex, ia numele de isocortex sau cortex omogenetic.
La rândul său, izocortexul poate fi împărțit în două structuri diferite, care alternează în diferite zone ale creierului, în funcție de funcția căreia îi sunt atribuite:
- Un cortexul homotipica, în cazul în care diferite tipuri de celule sunt reprezentate în mai mult sau mai puțin în aceeași măsură
- Un cortexul heterotipica, unde în loc să observăm prevalența [8] a unui tip neuronal peste toate celelalte.
Neocortexul este alcătuit din trei mari familii de neuroni: celule piramidale, celule spinoase nepiramidale și celule nepiramidale nepinoase.
- Celulele piramidale sunt cel mai frecvent tip neuronal al cortexului cerebral. Acestea se numesc astfel deoarece pirenoforul lor ia adesea o formă triunghiulară sau de balon și are un diametru de 10-80 µm. Corpurile lor celulare sunt localizate în special în lamina III (stratul piramidal) și V (stratul ganglionar) al cortexului cerebral. Celulele piramidale au un dendrit apical deosebit de lung și dezvoltat care urcă în straturile cortexului cu ramificații rare, scăzând progresiv în diametru până când devine arborizat difuz doar în straturile cele mai superficiale. În plus față de dendrita apicală, fiecare celulă piramidală are numeroase dendrite bazale mai scurte, care au un curs diagonal sau orizontal care variază de la câteva zeci de micrometri până la 1 mm, prin urmare tind să rămână în același strat al cortexului în pe care pirenoforul celulei. Dendritele bazale se ramifică difuz chiar și la o distanță mică de pirenofor. Atât dendrita apicală, cât și dendritele bazale ale celulelor piramidale sunt bogate în spini dendritici și cu cât este mai mare distanța de pirenofor, cu atât numărul lor tinde să fie mai mare. Axonul celulelor piramidale își are originea în general în porțiunea bazală a pirenoforului, adesea opusă dendritei apicale. Este subțire, nu are ramificații, merge profund cu un curs adesea perpendicular pe suprafața cortexului, mai întâi în straturile profunde ale cortexului, apoi în substanța albă unde continuă sub formă de fibre mielinizate. Celulele piramidale sunt neuroni de proiecție care utilizează neurotransmițători de excitare în sinapsele lor, în principal glutamat și aspartat.
- Celulele nepiramidale spinoase, numite și celule stelate spinoase, sunt cea mai abundentă familie de neuroni multipolari din cortexul cerebral după celulele piramidale. Corpurile lor celulare sunt mici cu un diametru de 5-10 µm și sunt localizate în special în lamina IV (stratul granular interior). Au numeroase dendrite primare cu spini care radiază de la pirenofor și parcurg o distanță variabilă în interiorul cortexului cerebral, ramificându-se difuz. Axonii lor sunt foarte des limitați la cortex și nu mielinizați, au un curs tendențial vertical și se îndreaptă spre lamina mai superficială a cortexului. Cel mai frecvent neurotransmițător din sinapsele lor este glutamatul.
- Celulele nepiramidale nepiramidale, numite și celule stelate non-spinoase, sunt cea mai puțin răspândită, dar cea mai eterogenă familie de neuroni din cortexul cerebral. De fapt, în ciuda numelui, unii dintre ei au spini dendritici, dar într-o măsură mai mică decât cele două familii anterioare. Sunt celule cu o formă foarte variabilă, unite prin axoni limitați la substanța cenușie și, prin urmare, tendențial nemelinizate sau ușor mielinizate. Cursul dendritelor sau axonilor lor poate fi orizontal, radial sau vertical, în funcție de tipul de celulă. Toți împărtășesc faptul că sunt interneuroni inhibitori, astfel încât neurotransmițătorul lor principal este GABA, rar glicina. Există o răspândire largă a veziculelor dense care conțin neuropeptide (chiar mai mult de un tip pe celulă), cum ar fi neuropeptida Y, substanța P, VIP, colecistochinina și somatostatina. În cadrul acestei clase de neuroni, numeroase celule se disting pe baza formei piroforilor sau a ramurilor dendritice sau axonale. Cele mai cunoscute sunt celulele orizontale, celulele coșului, celulele sfeșnicului, celulele cu buchet dublu, celulele Martinotti, celulele fusului, celulele neurogliaforme, celulele bipolare.
- Celulele orizontale sunt neuroni cu un pirenofor fusiform, cu axa majoră dispusă orizontal, foarte frecventă în lamina I (stratul molecular) și II (stratul granular exterior) al cortexului cerebral. Au dendrite lungi (comparativ cu alte celule nepiramidale) care se ramifică orizontal din corpul celulei, adesea ramificându-se în același strat în care se găsește pirenoforul. Axonii lor la scurt timp după ce provin din corpul celulei se împart în două ramuri care rulează orizontal în direcții opuse în același strat; adesea axonul provine dintr-un dendrit.
- Le cellule a canestro sono neuroni dal pirenoforo di forma irregolare e variabile per forma e dimensioni, diffuse nella lamina IV (strato granulare interno). Possiedono numerosi e corti dendriti che decorrono radialmente rispetto al pirenoforo e si ramificano in prossimità di esso. Gli assoni sono brevi, hanno decorso verticale ed in prossimità della loro cellula bersaglio (di norma una cellula piramidale) si ramificano orizzontalmente avviluppandola in una rete che ricorda un canestro, da qui il nome di queste cellule. Formano sinapsi assondendritiche o assosomatiche.
- Le cellule a candelabro possiedono un pirenoforo di forma variabile, spesso tondeggiante o ovalare posto nella lamina III (strato piramidale) della corteccia cerebrale, più raramente nella parte profonda della lamina II (strato granulare esterno) e in quella superficiale della lamina IV (strato granulare interno). L'albero dendritico origina o dal polo superiore o da quello inferiore del corpo. I loro assoni hanno decorso ascendente qualora queste cellule si trovino nella suddivisione IIIa dello strato piramidale, la più superficiale, in quella intermedia, la IIIb, possono avere decorso ascendente o discendente, mentre nella profonda, la IIIc, hanno decorso discendente. Gli assoni si ramificano poco dopo l'origine e si dirigono in prossimità della cellula piramidale bersaglio dove emettono rami verticali che seguono i coni d'emergenza degli assoni di questa.
- Le cellule a doppio bouquet oa doppio pennacchio sono neuroni dal pirenoforo ovalare o fusiforme, posto nelle lamine II (strato granulare esterno) e III (strato piramidale). Possiedono almeno due alberi dendritici, uno ascendente e uno discendente, ampiamente ramificati. Il loro assone poco dopo l'origine si divide in due rami, uno ascendente verso la lamina II e uno discendente verso la lamina V che emettono a loro volta dei rami collaterali.
- Le cellule di Martinotti sono una tipologia delle cellule a doppio pennacchio di cui fanno parte anche le cellule a doppio bouquet, i loro pirenofori, piccoli e irregolari, si trovano nelle lamine V (strato ganglionare) o VI (strato multiforme). I loro dendriti si dipartono da tutto il pirenoforo tranne che nella porzione basale, decorrono radialmente e sono piuttosto corti e ramificati. La loro caratteristica è un lungo assone ascendente originante dal polo superiore della cellula che si porta sino alla lamina I (strato molecolare) e qui si arborizza diffusamente.
- Le cellule fusiformi o bipolari, sono neuroni dal pirenoforo ovoidale situato nella lamina IV (strato granulare interno) della corteccia cerebrale. Possiedono un dendrite ascendente e un dendrite discendente con scarse ma lunghe ramificazioni che possono raggiungere rispettivamente lo strato II (strato granulare esterno) e lo strato VI (strato multiforme). Nel complesso l'albero dendritico che si forma è molto stretto. Il loro assone origina autonomamente o da un dendrite e si dirige verticalmente poco dopo l'origine, seguendo la conformazione di uno degli alberi dendritici.
- Le cellule neurogliaformi o "a ragnatela", sono neuroni dal pirenoforo sferico dal diametro di circa 10 µm, diffuse all'interno delle lamine II (strato granulare esterno), III (strato piramidale), o IV (strato granulare interno) con una posizione dipendente dalla tipologia di corteccia cerebrale. Possiedono un numero variabile di corti dendriti principali che decorrono radialmente rispetto al pirenoforo e si ramificano ampiamente, da qui la rassomiglianza con una ragnatela. L'albero dendritico che si viene a formare è pressoché sferico. Il loro assone decorre spesso orizzontalmente o trasversalmente, è più lungo dei dendriti (mediamente il triplo) e si ramifica distalmente.
Le cellule della neocorteccia sopraelencate possono essere ulteriormente divise in base alla direzione dei loro assoni, si distinguono perciò neuroni corticifughi (i cui assoni si dirigono verso la periferia) e neuroni corticicoli (i cui assoni si dirigono verso altri strati della corteccia, hanno funzioni associative). Di seguito è riportata una tabella riassuntiva.
| Neuroni corticifughi | Neuroni corticicoli |
|---|---|
| Cellule piramidali | Cellule a doppio pennacchio dendritico |
| Cellule polimorfe | Cellule del Martinotti |
| Granuli | Cellule orizzontali |
| Cellule marginali | |
| Cellule del 2º tipo di Golgi |
Sostanza bianca
La sostanza bianca [9] del telencefalo costituisce lo strato interposto fra il pallium ei nuclei della base: in una sezione condotta al di sopra del corpo calloso essa si presenta molto espansa e costituisce pertanto il cosiddetto centro semiovale. Questo è composto di diversi tipi di fibre, ossia:
- fibre di proiezione , che si distinguono a loro volta in:
- corticifughe (cioè originate da neuroni della corteccia e dirette a centri sottocorticali)
- corticipete (cioè a provenienza prevalentemente talamica, ottica e acustica e dirette verso neuroni corticali)
- fibre di associazione , che si distinguono a loro volta in:
- interemisferiche (cioè che collegano territori omologhi dei due emisferi), prevalentemente dirette o provenienti dal corpo calloso
- intraemisferiche (cioè che collegano circonvoluzioni più o meno distanti dello stesso emisfero), denominate anche fibre arcuate .

Più in profondità rispetto al centro semiovale, la sostanza bianca si organizza ad avvolgere con lamine i nuclei della base ei nuclei del talamo , formando tre capsule: la capsula interna , la capsula esterna e la capsula estrema .
- La capsula interna è una formazione bianca laminare interposta fra:
- medialmente, il talamo e il nucleo caudato
- lateralmente, il nucleo lenticolare.
- La maggior parte della capsula interna è costituita da fibre corticipete di provenienza talamica, mentre solo nella porzione più ventrale decorrono fibre corticifughe dirette al tronco encefalico e al midollo spinale . In una sezione orizzontale passante circa a livello dei tubercoli quadrigemelli superiori , si distinguono nella capsula interna un braccio posteriore, un ginocchio e un braccio anteriore, in cui decorrono le seguenti fibre:
- braccio posteriore : fibre cortico-spinali (per il midollo spinale), talamo-corticali (radiazione talamica superiore), in parte anche fibre cortico-tettali (per la lamina quadrigemina ), cortico-rubre (per il nucleo rosso ) e cortico-reticolari (per la formazione reticolare )
- ginocchio : fibre cortico-nucleari per i nuclei somatomotori dei nervi encefalici , fibre cortico-reticolari
- braccio anteriore : fibre talamo-corticali (radiazione talamica anteriore), fibre cortico-pontine (per il ponte , fibre cortico-ipotalamiche (per l' ipotalamo )
- La capsula esterna è una lamina sottile interposta fra:
- medialmente, la faccia laterale del putamen
- lateralmente, il claustro.
- Essa è attraversata principalmente da fibre cortico-tegmentali, dirette cioè verso il mesencefalo .
- La capsula estrema, molto esile, è interposta fra:
- medialmente, il claustro
- lateralmente, la corteccia del lobo dell'insula .
- Le fibre che la compongono sono per la maggior parte di associazione tra i lobi insulare, parietale e temporale.
Alla sostanza bianca del telencefalo appartengono anche alcuni sistemi commissurali , ossia di collegamento tra i due emisferi, che hanno la caratteristica di essere impari e mediani. Questi sistemi sono il corpo calloso, il setto pellucido, il fornice e la commessura anteriore.
Nuclei della base

I nuclei della base o gangli della base [10] sono formazioni grigie situate profondamente nella sostanza bianca telencefalica, in stretto rapporto con il talamo . Essi comprendono [11] :
- il claustro
- l' amigdala
- il corpo striato , comprendente a sua volta:
- il nucleo caudato
- il nucleo lenticolare , definizione che viene usata per indicare due nuclei strettamente connessi, il pallido ed il putamen .

Come tutte le formazioni encefaliche, anche i nuclei della base presentano differente origine filogenetica , che si può riassumere in una divisione in tre sezioni:
- l' archistriatum , la porzione più antica, è rappresentato dall'amigdala
- il paleostriatum è rappresentato dal pallido
- il neostriatum , la porzione più recente, comprende il caudato ed il putamen.
Amigdala
L' amigdala è una formazione grigia di forma globosa, situata in profondità rispetto al pallido, da cui è separata per l'interposizione delle fibre della capsula interna , e connessa con l'apice della coda del nucleo caudato; caudalmente, essa continua con l' uncus della circonvoluzione dell' ippocampo . Anatomicamente l'amigdala può essere assegnata al lobo temporale .
I nuclei che compongono l'amigdala possono essere distinti in due gruppi, ossia:
- nuclei corticomediali , comprendenti i nuclei amigdaloidei mediale, corticale e centrale, l'area amigdaloidea anteriore ed il nucleo della stria olfattiva laterale
- nuclei basilaterali , comprendenti i nuclei amigdaloidei laterale, basale e basale accessorio.
Il nucleo centrale dell'amigdala è implicato in fenomeni di aggressività. L'ablazione bilaterale dell'amigdala produce una sindrome comportamentale peculiare che si manifesta con abolizione dell'aggressività, in modo per certi aspetti simile a quanto avviene nella lobotomia frontale. Le proiezioni dell'amigdala si organizzano in due principali sistemi: la stria terminale e la via amigdalofuga.
Claustro
Il claustro è una sottile lamina di sostanza grigia, in continuità anteriormente con l' amigdala ed addossata alla corteccia insulare , dalla quale è separata soltanto per l'interposizione della capsula estrema ; medialmente, la capsula esterna lo separa invece dal putamen. Il suo significato funzionale non è del tutto noto.
Corpo striato

Il corpo striato è un voluminoso insieme di nuclei che deve il nome alla sua particolare organizzazione strutturale: esso è infatti composto da un'alternanza di formazioni grigie intersecate da fasci di sostanza bianca, che conferiscono alla struttura quel particolare aspetto "striato". Il corpo striato viene anatomicamente assegnato al lobo frontale .
La suddivisione in nuclei del corpo striato è operata dal sistema delle capsule di sostanza bianca del telencefalo. La capsula interna distingue così:
- il nucleo caudato , in posizione superomediale
- il nucleo lenticolare , in posizione inferolaterale.
Ulteriori suddivisioni vengono poi fatte all'interno di questi nuclei dagli altri tralci di sostanza bianca.
- Il nucleo caudato ha la forma di una grossa virgola a direzione sagittale che contorna il margine superiore del talamo , da cui è separato per mezzo del solco optostriato . Esso si dispone a ricoprire la capsula interna, di cui forma il limite superiore. In esso si possono distinguere:
- una testa arrotondata, che costituisce l'estremità anteriore del nucleo e che, seguendo il decorso dei due talami, in avanti converge fin quasi a toccare la sua corrispondente dell'altro emisfero, da cui rimane separata solo per l'interposizione del setto pellucido
- un corpo allungato, che sormonta il talamo per tutta la sua lunghezza
- una coda , che curva sotto la capsula interna e si assottiglia fino a terminare nell' amigdala .
- Il nucleo lenticolare è una formazione triangolare, con la punta rivolta verso il centro dell'encefalo, separato dal claustro per l'interposizione della capsula esterna. Esso viene poi ulteriormente suddiviso in due porzioni per l'interposizione di una sottile lamina di sostanza bianca, la lamina midollare laterale , che individua:
- il putamen , lateralmente ad essa, che costituisce la base del triangolo
- il globo pallido (o globus pallidus ), medialmente ad essa. Il pallido può essere poi suddiviso in due ulteriori sezioni per l'interposizione di una seconda lamina bianca, detta lamina midollare mediale , che individua un pallido esterno ed un pallido interno : quest'ultimo costituisce l'apice del triangolo.
Funzioni
I nuclei della base svolgono un ruolo importante nel controllo dei movimenti, volontari e meno, ma anche di alcune funzioni cognitive, tanto è vero che le delezioni o l'isolamento di queste strutture del telencefalo creano difficoltà nelle attività intellettive superiori. Il controllo sulle attività motorie viene svolto grazie a notevoli connessioni con le vie discendenti (fasci di fibre direttamente connesse con interneuroni o motoneuroni sinaptati con effettori motori); queste connessioni non sono dirette con i fasci discendenti come si potrebbe pensare ma indirettamente tramite l'area 4 della corteccia (area motoria primaria). I nuclei (o gangli) della base, rielaborano quindi informazioni provenienti dalla corteccia e le riproiettano ad essa costituendo delle vere e proprie vie. Si possono quindi distinguere quattro circuiti:
- Circuito Scheletromotorio: controlla la messa in atto dei movimenti volontari e involontari (per es. la postura). Raccoglie informazioni provenienti dalle aree motorie e somestetiche, per poi proiettare verso le aree motorie frontali.
- Circuito Oculomotorio: controlla i movimenti saccadici degli occhi. Raccoglie informazioni provenienti dalle aree visive frontali (campi oculari frontali, cortecce orbitofrontali laterale e mediale, campi oculari supplementari) per poi proiettare verso il mesencefalo (dove si trovano i nuclei dei nervi cranici che innervano la muscolatura orbitaria).
- Circuito Prefrontale: è coinvolto nella pianificazione dei movimenti. Raccoglie informazioni dalle regioni frontali e parietali per poi proiettare verso le aree frontali.
- Circuito Limbico: presenta connessioni con il sistema limbico ed è quella parte deputata all'influenza emozionale, mnemonica ed esperienziale (apprendimento) sul movimento.
Possiamo inoltre individuare tre vie per l'elaborazione dell'informazione:
- Nella via diretta le informazioni provenienti dalla corteccia fanno sinapsi nello striato, da qui connessioni inibitorie (GABAergiche) raggiungono il complesso costituito da SNr-GPi (Sostanza nera del Sömmering e Globus pallidus interno); da qui sempre con proiezioni inibitorie si giunge al talamo (nuclei interessati: ventrale anteriore, ventrale laterale, dorsale mediale) il quale proietta con modalità eccitatoria verso la corteccia motoria. In questo caso la corteccia risulta essere attivata.
- Nella via indiretta le informazioni provenienti dalla corteccia raggiungono lo striato, da qui connessioni inibitorie (GABAergiche) raggiungono il globus pallidus esterno (GPe), il quale a sua volta proietta in modo inibitorio al nucleo subtalamico di Luys, che risulta quindi disinibito e può scaricare sul complesso SNr-GPi eccitandoli. SNr-GPi inibiscono quindi il talamo e di conseguenza, la corteccia risulta essere inibita perché il talamo stesso è inibito.
- La via iperdiretta raccoglie le informazioni provenienti dalla corteccia e le trasmette al SNr-GPi, passando per il nucleo subtalamico; ovviamente da SNr-GPi raggiungono talamo e corteccia.
Anatomia comparata
Il telencefalo è una vescicola che dimostra avere con l'evoluzione la maggior plasticità mostrando peculiarità maggiori da una classe evolutiva all'altra perché, di fatto, l'evoluzione di questa vescicola coincide perfettamente con l'evoluzione dei Vertebrati. Il telencefalo è formato da due vescicole che possono evolversi o per eversione o per inversione; nel primo caso daranno origine a un telencefalo unico mentre nel secondo caso daranno origine a un telencefalo a due emisferi separati. La differenza principale fra l'everso e l'inverso è la posizione degli elementi costitutivi che lo compongono i quali rimangono costanti in tutte le classi evolutive; questa vescicola infatti è composta sempre da una archicorteccia dorsomediale, una paleocorteccia dorsolaterale, lo striato ventrolaterale ei nuclei del setto ventromediali.
Patologia
Nel seguito è un elenco di alcune famiglie di malattie pertinenti gli emisferi cerebrali.
- malattie traumatiche
- malattie infiammatorie
- malattie circolatorie
- malattie tumorali
- malattie degenerative primarie
- malattie dello sviluppo
- malattie ereditarie
- malattie congenite
- malattie infettive
- malattie rare
- malattie genetiche
Nel seguito è un elenco di alcune malattie del telencefalo.
Note
- ^ Bear, Neuroscienze. Esplorando il cervello , pag. 173
- ^ ( EN ) Giuseppe C. Balboni, et al. , Formazione e sviluppo delle vescicole encefaliche , in Anatomia Umana, Vol. 3 , Ristampa 2000, Milano, Edi. Ermes srl, 1976, pp. 20-21, ISBN 88-7051-078-6 .
- ^ ( EN ) Giuseppe C. Balboni, et al. , Configurazione esterna , in Anatomia Umana, Vol. 3 , Ristampa 2000, Milano, Edi. Ermes srl, 1976, p. 135, ISBN 88-7051-078-6 .
- ^ ( EN ) Giuseppe C. Balboni, et al. , Scissure primarie , in Anatomia Umana, Vol. 3 , Ristampa 2000, Milano, Edi. Ermes srl, 1976, pp. 136-138, ISBN 88-7051-078-6 .
- ^ ( EN ) Giuseppe C. Balboni, et al. , Lobi e circonvoluzioni , in Anatomia Umana, Vol. 3 , Ristampa 2000, Milano, Edi. Ermes srl, 1976, pp. 139-145, ISBN 88-7051-078-6 .
- ^ ( EN ) Giuseppe C. Balboni, et al. , Aree corticali , in Anatomia Umana, Vol. 3 , Ristampa 2000, Milano, Edi. Ermes srl, 1976, pp. 176-180, ISBN 88-7051-078-6 .
- ^ ( EN ) Giuseppe C. Balboni, et al. , Corteccia telencefalica , in Anatomia Umana, Vol. 3 , Ristampa 2000, Milano, Edi. Ermes srl, 1976, p. 166, ISBN 88-7051-078-6 .
- ^ "Prevalenza" non vuol dire "esclusività": nella corteccia cerebrale umana, infatti, non si osserva mai una zona caratterizzata dalla presenza di cellule di un tipo soltanto, ma solo aree dove un certo tipo neuronale è più rappresentato degli altri, che sono comunque presenti.
- ^ ( EN ) Giuseppe C. Balboni, et al. , Organizzazione della sostanza bianca del telencefalo. , in Anatomia Umana, Vol. 3 , Ristampa 2000, Milano, Edi. Ermes srl, 1976, p. 157, ISBN 88-7051-078-6 .
- ^ ( EN ) Giuseppe C. Balboni, et al. , Nuclei del telencefalo , in Anatomia Umana, Vol. 3 , Ristampa 2000, Milano, Edi. Ermes srl, 1976, p. 147, ISBN 88-7051-078-6 .
- ^ Per gli stretti rapporti funzionali che contraggono con i nuclei della base, vengono talvolta inclusi in questo gruppo anche il nucleo subtalamico e la sostanza nera . Tuttavia, sebbene dal punto di vista fisiologico possano entrare a pieno diritto nel circuito che coinvolge i nuclei della base, dal punto di vista anatomico queste due formazioni appartengono rispettivamente al subtalamo ed al mesencefalo , dove saranno descritte.
Bibliografia
- Giuseppe C. Balboni, et al. , Anatomia Umana, Vol. 3. , Ristampa 2000, Milano, Edi. Ermes srl, 1976, ISBN 88-7051-078-6 .
- Eric R. Kandel, Schwartz James H.; Jessell Thomas M., Principi di neuroscienze , a cura di Perri V., Spidalieri G., 3ª edizione, CEA, 2003, ISBN 88-408-1256-3 .
- Mark F. Bear, Connors Barry W.; Paradiso Michael A., Neuroscienze. Esplorando il cervello. , a cura di Casco C., Petrosini L., Oliveri M., 3ª edizione, Elsevier, 2007, ISBN 88-214-2943-1 .
Voci correlate
- Encefalo
- Tronco encefalico
- Cervelletto
- Midollo spinale
- Poli degli emisferi cerebrali
- Solchi orbitali
Altri progetti
-
 Wikimedia Commons contiene immagini o altri file su telencefalo
Wikimedia Commons contiene immagini o altri file su telencefalo
Collegamenti esterni
- ( EN ) Telencefalo , su Enciclopedia Britannica , Encyclopædia Britannica, Inc.
- Atlante Anatomico del Sistema Nervoso Centrale - Telencefalo (dal sito dell'Università di Urbino "Carlo Bo")
| Controllo di autorità | Thesaurus BNCF 37784 · LCCN ( EN ) sh85133359 · GND ( DE ) 4121188-1 · BNF ( FR ) cb12261508d (data) · NDL ( EN , JA ) 01087188 |
|---|