Pteranodon
| Pteranodon | |
|---|---|
 | |
| Gama geologică | |
| Starea de conservare | |
Fosil | |
| Clasificare științifică | |
| Domeniu | Eukaryota |
| Regatul | Animalia |
| Phylum | Chordata |
| Clasă | Sauropsida |
| Ordin | † Pterosaurie |
| Subordine | † Pterodactyloidea |
| Familie | † Pteranodontidae |
| Subfamilie | † Pteranodontinae Marsh , 1876 |
| Tip | † Pteranodon Marsh, 1876 |
| Nomenclatura binominala | |
| † Pteranodon longiceps Marsh, 1876 | |
| Sinonime | |
| |
Pteranodon (al cărui nume înseamnă „aripă fără dinți“) este un general pe cale de disparitie pterosaur pterodactyloide pteranodontide trăit în Late Cretacicului , acum aproximativ 86-84,500,000 ani ( în Santonian ), în actualul Kansas , Alabama , Nebraska , Wyoming si Dakota de Sud , Statele Unite ale Americii . Specia tip a genului, P. longiceps , este unul dintre cei mai mari pterozauri cunoscuți, cu o anvergură a aripilor de peste 6 metri. Pteranodonul este universal cel mai cunoscut pterosaur, de asemenea datorită colecției remarcabile de fosile, superioară oricărui alt pterosaur, cu aproximativ 1.200 de exemplare cunoscute, dintre care multe au fost bine conservate cu cranii aproape complete și schelete articulate: abundența sa indică faptul că acest animal a fost o parte importantă a comunității de animale din Marea Interioară de Vest . [1]
Deși este prezentat în mod repetat în cultura de masă ca dinozaur , Pteranodonul a fost un pterosaur . Prin definiție, toți dinozaurii aparțin unuia dintre cele două grupuri din Dinosauria , și anume Saurischia sau Ornithischia . Ca atare, acest lucru exclude pterosaurii. Deși nu sunt dinozauri, pterosaurii formează o cladă cu dinozauri în cadrul cladei Avemetatarsalia . [2]
Descriere


Speciile de pteranodoni sunt extrem de bine reprezentate în înregistrarea fosilelor, permițând descrieri detaliate ale anatomiei și ontogenezei lor. În prezent, au fost identificate peste 1.000 de exemplare, deși mai puțin de jumătate sunt suficient de complete pentru a oferi cercetătorilor informații anatomice valide. Cu toate acestea, numărul exemplarelor colectate este mai mare decât orice altă specie de pterosauri și include atât exemplare masculine, cât și femele, cu diferite grupe de vârstă și, eventual, mai multe specii. [2]
Exemplarele adulte de Pteranodon din speciile principale pot fi împărțite în două clase de mărime distincte. Cea mai mică clasă sunt exemplarele cu cea mai mică și mai rotunjită crestă craniană și canale pelviene foarte largi. Mărimea canalului pelvian a permis probabil acestor indivizi să depună ouă , fiind identificați ca indicând exemplare femele. Clasa mai mare, reprezentând masculii, are șolduri înguste și creste craniene foarte mari, care au fost probabil folosite în recunoașterea intraspecifică și curte.
Un bărbat adult Pteranodon a fost printre cei mai mari pterosauri care au trăit vreodată, precum și unul dintre cele mai mari animale zburătoare cunoscute până la sfârșitul secolului XX, când au fost descoperiți pterosaurii uriași azhdarhi . [2] Anvergura aripilor unui bărbat adult mediu Pteranodon a fost de 5,6 metri. Femelele adulte au fost mult mai mici, având o medie de 3,8 metri într-o anvergură a aripilor. Cel mai mare exemplar de Pteranodon longiceps , găsit în cadrul formațiunii Niobrara , măsura 6,25 metri în anvergură. Un exemplar și mai mare, cunoscut din formația Pierre Shale, are o anvergură a aripilor de 7,25 metri, deși acest exemplar poate aparține genului și speciei distincte Geosternbergia maysei . [2] Deși majoritatea exemplarelor fosile se găsesc zdrobite, există suficiente fosile în trei dimensiuni pentru a pune împreună o descriere detaliată a animalului.
Metodele folosite pentru estimarea masei corporale a exemplarelor mari de Pteranodon (cele cu anvergura aripilor de aproximativ 7 metri) sunt notoriu nesigure, rezultând o gamă largă de estimări de la 20 de kilograme până la 93 de kilograme. Într-o revizuire a estimărilor mărimii pterosaurilor publicată în 2010, cercetătorii Mark Witton și Mike Habib au arătat că estimările mai mari sunt aproape sigur greșite, având în vedere volumul total al corpului lui Pteranodon , și ar putea fi corecte doar dacă animalul. „Era format în principal din aluminiu ”. [3] Witton și Habib au luat în considerare metodele utilizate de cercetătorii care au obținut estimări de masă mai mici, dar la fel de defectuoase. Cele mai multe au fost produse prin scalarea animalelor moderne, cum ar fi liliecii și păsările, la dimensiunea Pteranodonului , în ciuda faptului că pterosaurii au proporții corporale și anatomice diferite față de orice alt animal viu. [3]
Alte trăsături distinctive care diferențiază Pteranodonul de alți pterosauri sunt coloanele vertebrale subțiri ale vertebrelor, ligamentele osoase asemănătoare plăcilor care întăresc vertebrele deasupra șoldului și o coadă relativ lungă. Pteranodon , de fapt, avea o vertebră caudală „sacralizată” (adică o parte a sacului sinusal ) și 11 sau mai multe vertebre caudale libere făcute foarte deosebite de prezența elementelor distale care formează o pereche de centre (porțiunea centrală a vertebrelor) topite și alungite, pe care le-au separat distal, formând două „tije caudale” paralele. Această structură a fost considerată inițial o formă de proiecție caudală pentru vertebrele posterioare, dar în schimb pare a fi o serie de vertebre mici fuzionate cu o canelură în centru, aparent parte a arcului neuronal. [4] Lungimea acestei structuri a dat lui Pteranodon o coadă neobișnuit de lungă pentru un pterodactiloid, cu o lungime a cozii de aproximativ 3,5% din anvergura aripilor sau până la 25 de centimetri la masculii mai mari. [4] Cu toate acestea, funcția acestei structuri curioase rămâne neclară: Bennett (2001) a propus că a susținut și a controlat o mică uropatie întinsă între coapsele animalului, dar acest lucru este în contrast cu fosilele de pterosauri care sugerează că coada nu era conectat la uropatie. Este posibil ca Pteranodon să fi fost excepția de la regulă, dar alte funcții, cum ar fi un suport pentru înot, ancorarea pentru structurile de afișare a țesuturilor moi sau o paletă, nu pot fi excluse (Witton, 2013).
Craniul și ciocul

Spre deosebire de pterozaurii primitivi, cum ar fi Rhamphorhynchus și Pterodactylus , Pteranodon poseda un cioc fără dinți, similar cu cel al păsărilor. Ciocul Pteranodonului era format din margini osoase solide care ieșeau din baza maxilarelor. Ciocul era lung, subțire și se termina într-un vârf ascuțit și subțire. Maxilarul, care era mai lung decât mandibula, era curbat în sus; în timp ce acest lucru a fost atribuit în mod normal numai ciocului care se îndoaie în sus, un exemplar (UALVP 24238) are o curbură corespunzătoare lățimii ciocului spre vârf. Deși vârful ciocului nu este cunoscut la acest specimen, nivelul de curbură sugerează că ciocul acestui specimen trebuie să fi fost extrem de lung. Forma unică de cioc din acest specimen l-a determinat pe Alexander Kellner să o atribuie unui gen distinct, Dawndraco , în 2010. [5] Cu toate acestea, studiile ulterioare ale Elizabeth Martin-Silverstone și colab. (2017) au arătat că diferențele de cioc între indivizi erau probabil o caracteristică comună la pteranodontide . Prin urmare, au ajuns la concluzia că „ Dawndraco ” a fost pur și simplu un mascul de Geosternbergia sternbergi . [6]
Mai mult, mulți cercetători cred că, după toate probabilitățile, Pteranodonul deținea un sac mic sub cioc, similar ca funcție cu cel al pelicanului de astăzi, iar anumite urme fosile ar părea să indice acest lucru. [7]
Cea mai distinctivă caracteristică a Pteranodonului este voluminosul său crestet cranian. Aceste creste erau formate din osul frontal care ieșea în spatele craniului și în sus. Mărimea și forma acestor creste variază în funcție de o serie de factori, inclusiv vârsta, sexul și speciile. Masculii din Pteranodon sternbergi , cea mai veche specie descrisă până în prezent (și uneori plasate în genul distinct Geosternbergia ), posedau o creastă verticală cu o proiecție largă înainte, în timp ce descendenții lor, Pteranodon longiceps , posedau o creastă mai îngustă și se așezau înapoi în cap . [1] Femelele ambelor specii, pe de altă parte, aveau creste mici și rotunjite. [8] Crestele erau probabil structuri expoziționale și comunicative, deși puteau avea și alte funcții. [9] Se crede că Pteranodonii ar putea folosi creasta craniană pentru a schimba poziția în timpul zborului ca un fel de cârmă.
Clasificare
Evoluţie

Majoritatea fosilelor Pteranodon provin din formațiunea Niobrara , din centrul Statelor Unite . Pteranodonul a trăit mai mult de 4 milioane de ani, între Coniac târziu și Campanian timpuriu, în Cretacicul superior . [8] Genul este prezent în majoritatea straturilor formațiunii niobrare, cu excepția celor două superioare; în 2003 Kenneth Carpenter a examinat distribuția și datarea fosilelor din această formațiune, arătând că speciile presupuse P. sternbergi au trăit cu 88-85 milioane de ani în urmă, în timp ce speciile ulterioare P. longiceps au trăit între 86-84, 5 milioane de ani în urmă. O posibilă a treia specie, pe care Kellner a numit-o Geosternbergia maysei , în 2010, a fost găsită în Sharon Springs din Formația Pierre Shale, din Kansas , Wyoming și Dakota de Sud și pare să fi trăit între 81,5-80,5 milioane de ani. [10]
La începutul anilor 1990, Bennett a remarcat faptul că cele două forme principale de Pteranodon prezente în formațiunea Niobrara au fost separate cu exactitate în timp, cu puține suprapuneri, dacă nu există. Din această cauză și a asemănării lor generale, Bennett a sugerat că cele două forme reprezentau cronospecii într-o singură linie evolutivă care durează aproximativ 4 milioane de ani. Cu alte cuvinte, doar o singură specie de Pteranodon ar fi fost prezentă la un moment dat, iar P. sternbergi (sau Geosternbergia ) a fost, probabil, strămoșul direct al P. longiceps . [2]
Specii valabile

Mulți cercetători susțin că există cel puțin două specii de Pteranodon . Cu toate acestea, în afară de diferențele dintre bărbați și femele descrise mai sus, scheletele post-craniene ale Pteranodonului prezintă variații mici sau deloc între specii sau specimene, iar corpul și aripile tuturor Pteranodonilor au fost în esență identice. [8]
În mod tradițional, două specii de Pteranodon sunt recunoscute ca fiind valabile: specia tip Pteranodon longiceps și Pteranodon sternbergi . Cele două specii diferă doar prin forma creastei la masculii adulți (descrisă mai sus) și posibil prin unghiul unor oase ale craniului. [8] Deoarece fosilele bine conservate ale craniului Pteranodon sunt extrem de rare, cercetătorii folosesc stratigrafia (adică ce strat de rocă din formațiunea geologică în care se găsește o fosilă) pentru a determina identitatea speciei în majoritatea cazurilor.
Pteranodon sternbergi este singura specie cunoscută de Pteranodon cu creastă verticală. Maxilarul inferior al P. sternbergi avea 1,25 metri lungime. [11] Specia a fost recuperată de George F. Sternberg în 1952 și descrisă de John Christian Harksen în 1966 din partea de jos a formațiunii Niobrara. Specia este mai veche decât P. longiceps și este considerată de Bennett drept strămoșul direct al speciilor ulterioare. [8]
Deoarece fosilele identificabile ca P. sternbergi se găsesc exclusiv în straturile inferioare ale formațiunii Niobrara, iar fosilele P. longiceps se găsesc exclusiv în straturile superioare, o fosilă fără sculele poate fi identificată pe baza locației sale în coloana geologică (deși de mulți descoperirile timpurii ale fosilelor, nu au fost înregistrate date precise privind localizarea lor, ceea ce face multe fosile neidentificabile). [5]
Mai jos este o cladogramă care prezintă rezultatele unei analize filogenetice prezentate pentru prima dată de Andres și Myers (2013) [12] și actualizate cu date suplimentare de la Andres și colab. (2018): [13]
| Ornithocheiroidea |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Clasificări alternative

Datorită variațiilor subtile dintre exemplarele pteranodontide din cadrul formațiunii Niobrara, mulți cercetători au atribuit toate materialele găsite genului Pteranodon , la cel puțin două specii ( P. longiceps și P. sternbergi ) care se disting în principal prin forma creastei . Cu toate acestea, clasificarea acestor două forme variază de la cercetător la cercetător. În 1972, Halsey Wilkinson Miller a publicat un articol în care susținea că diferitele forme ale Pteranodonului erau suficient de diferite pentru a fi plasate în subgenuri distincte. El a numit aceste exemplare drept Pteranodon ( Occidentalia ) occidentalis (pentru specia dezafectată acum P. occidentalis ) și Pteranodon ( Sternbergia ) sternbergi . Cu toate acestea, numele Sternbergia era deja utilizat, iar în 1978 Miller a redenumit specia Pteranodon ( Geosternbergia ) sternbergi , denumind o a treia combinație subgen / specie pentru P. longiceps , ca Pteranodon ( Longicepia ) longiceps . Cu toate acestea, cercetătorii principali de pterosauri de la sfârșitul secolului al XX-lea, inclusiv S. Christopher Bennett și Peter Wellnhofer, nu au adoptat aceste nume subgenerice și au continuat să plaseze toate exemplarele pteranodontide din genul unic Pteranodon .
În 2010, cercetătorul pterosaur Alexander Kellner a revizuit clasificarea HW Miller. Kellner, urmând ideile lui Miller că diferențele dintre speciile Pteranodon au fost suficiente pentru a le clasifica în diferite genuri, plasate P. sternbergi în genul Miller-numit, Geosternbergia , împreună cu specimenul Pierre Shale craniu , care Bennett a avut anterior. Considerat un mare P de sex masculin longiceps . Kellner a susținut că creasta acestui exemplar, deși nu a fost complet conservată, era foarte asemănătoare cu cea a Geosternbergiei . Deoarece specimenul era cu milioane de ani mai tânăr decât orice Geosternbergia cunoscută, Kellner l-a atribuit noii specii Geosternbergia maysei . Numeroase alte exemplare de pteranodont sunt cunoscute în aceeași formațiune, iar Kellner a sugerat că ar putea aparține aceleiași specii ca G. maysei , dar pentru că le lipsește craniile, el nu a putut să le identifice cu încredere. [5]
Specii renegate

Mai multe specii de Pteranodon au fost denumite încă din 1870, deși majoritatea sunt considerate acum sinonime junior ale a două sau trei specii valide. Cea mai cunoscută specie este specia tip , P. longiceps , bazată pe un exemplar bine conservat, inclusiv cel mai vechi craniu cunoscut din SW Williston. Acest individ avea o anvergură a aripilor de 7 metri (23 de picioare). [14] Alte specii valabile includ posibila specie P. sternbergi , cu o anvergură a aripilor estimată inițial de 9 metri. [14] Speciile P. occidentalis , P. velox , P. umbrosus , P. harpyia și P. comptus sunt considerate nomina dubia de Bennett (1994), iar alți cercetători pun la îndoială validitatea lor. Probabil că toate reprezintă sinonime ale celor mai cunoscute specii.
Deoarece trăsătura distinctivă cheie pe care Marsh a remarcat-o în Pteranodon a fost lipsa de dinți, orice fragmente de maxilar fără pterosaur, oriunde s-au găsit în lume, au avut tendința de a fi atribuite lui Pteranodon , la sfârșitul secolului al XIX-lea și începutul secolului al XIX-lea. Acest lucru a dus la numirea unei multitudini de specii noi și la o mare confuzie taxonomică. Numele Pteranodon a devenit refugium peccatorum , cam așa cum s-a întâmplat pentru dinozaurul Megalosaurus , pentru a eticheta toate rămășițele pterosaurilor care nu se puteau distinge decât prin absența dinților. Speciile (adesea cu reputație dubioasă bazate pe variații sexuale sau trăsături juvenile) au fost reclasificate de mai multe ori, iar mai multe subgenuri au fost ridicate în anii 1970 de către Halsey Wilkinson Miller pentru a le menține în diferite combinații, confuzând în continuare taxonomia ( subgenele includ Longicepia , Occidentalia și Geosternbergia ). Autori de seamă care au discutat despre diferitele aspecte ale lui Pteranodon includ Bennett, Padian, Unwin, Kellner și Wellnhofer. Două specii, P. orogensis și P. orientalis , nu erau nici măcar pteranodontide și au fost redenumite Bennettazhia oregonensis și , respectiv, Bogolubovia orientalis .
Lista speciilor și sinonimelor
Starea numelor enumerate mai jos se bazează pe un sondaj realizat de Bennett, 1994, dacă nu se indică altfel. [8]
| Nume | Autor | Ani | Stat | Notă |
|---|---|---|---|---|
| Pterodactylus occidentalis | Mlaştină | 1872 | Reclasificat ca Pteranodon occidentalis | Reclasificat ca Pterodactylus oweni Marsh 1871 (preocupat de Seeley 1864) |
| Pterodactylus ingens | Mlaştină | 1872 | Reclasificat ca Pteranodon ingens | |
| Pterodactylus velox | Mlaştină | 1872 | Nomen dubium | Reclasificat ca Pteranodon velox |
| Ornithochirus umbrosus | Face față | 1872 | Nomen dubium | |
| Ornithochirus harpyia | Face față | 1872 | Nomen dubium | |
| Pterodactylus umbrosus | (Cope) Cope | ( 1872 ) 1874 | Reclasificat ca Ornithochirus umbrosus | |
| Pteranodon longiceps | Mlaştină | 1876 | Valabil | Specii tip |
| Pteranodon ingens | (Marsh) Williston | ( 1872 ) 1876 | Nomen dubium | Reclasificat ca Pterodactylus ingens |
| Pteranodon occidentalis | Mlaştină | ( 1872 ) 1876 | Nomen dubium | Reclasificat din Pterodactylus occidentalis |
| Pteranodon velox | Mlaştină | ( 1872 ) 1876 | Nomen dubium | Reclasificat din Pterodactylus velox , pe baza unui exemplar tânăr |
| Pteranodon gracilis | Mlaştină | 1876 | Reclasificat ca Nyctosaurus gracilis | |
| Pteranodon comptus | Mlaştină | 1876 | Nomen dubium | |
| Pteranodon nanus | Mlaştină | 1876 | Reclasificat ca Nyctosaurus nanus | |
| Ornithocheirus umbrosus | (Cope) Newton | ( 1872 ) 1888 | Reclasificat ca Pteranodon umbrosus | Corectarea ortografică a Ornithochirus umbrosus |
| Ornithocheirus harpyia | (Cope) Newton | ( 1872 ) 1888 | Reclasificat ca Pteranodon harpyia | Corectarea ortografică a Ornithochirus harpyia |
| Pteranodon umbrosus | (Cope) Williston | ( 1872 ) 1892 | Nomen dubium | Reclasificat ca Ornithochirus umbrosus |
| Ornithostoma ingens | (Marsh) Williston | ( 1872 ) 1893 | Sinonim al lui Pteranodon ingens | Reclasificat din Pteranodon ingens |
| Ornithostoma umbrosum | (Cope) Williston | ( 1872 ) 1897 | Sinonim al lui Pteranodon umbrosus | Reclasificat din Pteranodon umbrosus |
| Pteranodon oregonensis | Gilmore | 1928 | Reclasificat ca Bennettazhia oregonensis | |
| Pteranodon sternbergi | Harksen | 1966 | Valabil | |
| Pteranodon marshi | Miller | 1972 | Sinonim al lui Pteranodon longiceps | |
| Pteranodon bonneri | Miller | 1972 | Reclasificat ca Nyctosaurus bonneri | |
| Pteranodon walkeri | Miller | 1972 | Sinonim al lui Pteranodon longiceps | |
| Pteranodon (Occidentalia) eatoni | (Miller) Miller | ( 1972 ) 1972 | Sinonim al lui Pteranodon sternbergi | |
| Pteranodon eatoni | (Miller) Miller | ( 1972 ) 1972 | Sinonim al lui Pteranodon sternbergi | Reclasificat din Pteranodon (Occidentalia) eatoni |
| Pteranodon (Longicepia) longicps | (Marsh) Miller | ( 1872 ) 1972 | Sinonim al lui Pteranodon longiceps | Reclasificat din Pteranodon longiceps |
| Pteranodon (Longicepia) marshi | (Miller) Miller | ( 1972 ) 1972 | Sinonim al lui Pteranodon longiceps | Reclasificat din Pteranodon marshi |
| Pteranodon (Sternbergia) sternbergi | (Harksen) Miller | ( 1966 ) 1972 | Reclasificat ca Pteranodon (Geosternbergia) sternbergi | Reclasificat din Pteranodon sternbergi |
| Pteranodon (Sternbergia) walkeri | (Miller) Miller | ( 1972 ) 1972 | Reclasificat ca Pteranodon (Geosternbergia) walkeri | Reclasificat din Pteranodon walkeri |
| Pteranodon (Pteranodon) marshi | (Miller) Miller | ( 1972 ) 1973 | Sinonim al lui Pteranodon longiceps | Reclasificat din Pteranodon marshi |
| Pteranodon (Occidentalia) occidentalis | (Mlaștină) Olshevsky | ( 1872 ) 1978 | Sinonim al lui Pteranodon occidentalis | Reclasificat din Pteranodon occidentalis |
| Pteranodon (Longicepia) ingens | (Mlaștină) Olshevsky | ( 1872 ) 1978 | Sinonim al lui Pteranodon ingens | Reclasificat din Pteranodon ingens |
| Pteranodon (Pteranodon) ingens | (Mlaștină) Olshevsky | ( 1872 ) 1978 | Sinonim al lui Pteranodon ingens | Reclasificat din Pteranodon ingens |
| Pteranodon (Geosternbergia) walkeri | (Miller) Miller | ( 1972 ) 1978 | Sinonim al lui Pteranodon longiceps | Reclasificat din Pteranodon walkeri |
| Pteranodon (Geosternbergia) sternbergi | (Harksen) Miller | ( 1966 ) 1978 | Sinonim al lui Pteranodon sternbergi | Reclasificat din Pteranodon (Sternbergia) sternbergi |
| Pteranodon orientalis | (Bogolubov) Nesov și Yarkov | ( 1914 ) 1989 | Reclasificat ca Bogolubovia orientalis | Reclasificat din Ornithostoma orientalis |
| Geosternbergia walkeri | (Miller) Olshevsky | ( 1872 ) 1991 | Sinonim al lui Pteranodon sternbergi | Reclasificat din Pteranodon (Sternbergia) walkeri |
| Geosternbergia sternbergi | (Harksen) Olshevsky | ( 1966 ) 1991 | Sinonim al lui Pteranodon sternbergi | Reclasificat din Pteranodon (Geosternbergia) sternbergi |
Istoria descoperirii

Primele descoperiri
Pteranodonul a fost primul pterosaur găsit în afara Europei . Fosilele sale au fost descoperite pentru prima dată de Othniel Charles Marsh în 1870, în depozitele de calcar din Cretacicul superior de pe Smoky Hill din vestul Kansasului. Aceste paturi de gips s-au așezat pe fundul a ceea ce a fost odată Marea Interioară de Vest , o mare mare de mică adâncime care se întindea în ceea ce este acum partea centrală a continentului nord-american. Aceste exemplare timpurii, YPM 1160 și YPM 1161, constau din oase parțiale ale aripilor, precum și dinți ai peștilor preistorici Xiphactinus , pe care Marsh i-a atribuit din greșeală acestui nou pterosaur (toți pterosaurii cunoscuți până atunci aveau dinți). În 1871, Marsh a numit descoperirea „ Pterodactylus oweni ”, atribuindu-l binecunoscutului (dar mult mai mic) gen european Pterodactylus . În 1871, Marsh a adunat oase noi de pe aripile acestui pterosaur mare. Dându-și seama că numele ales de el fusese deja folosit pentru o altă specie europeană de pterosaur, Pterodactylus oweni , numit de Harry Seeley în 1864, Marsh și-a redenumit pterosaurul gigant din America de Nord drept Pterodactylus occidentalis , denumirea „deget cu aripi occidentale”, în descrierea noului exemplar din 1872. De asemenea, el a numit alte două specii, pe baza diferențelor de mărime: Pterodactylus ingens (cel mai mare exemplar colectat până acum) și Pterodactylus velox (cel mai mic). [8]
Între timp, rivalul lui Marsh, Edward Drinker Cope , a dezgropat și câteva exemplare ale marelui pterosaur nord-american. Pe baza acestor exemplare, Cope a numit două specii noi, Ornithochirus umbrosus și Ornithochirus harpyia , în încercarea de a le atribui marelui gen european Ornithocheirus , deși a scris greșit numele (uitând „e”). [8] Lucrarea lui Cope numind specia sa a fost publicată în 1872, la doar cinci zile după lucrarea lui Marsh. Acest lucru a provocat o dispută, luptată în literatura publicată, asupra cărui nume a luat prioritate față de care, evident, erau aceleași specii. [8] Cope a recunoscut în 1875 că numele lui Marsh au luat prioritate față de ale sale, dar a susținut că Pterodactylus umbrosus era o specie distinctă (dar nu un gen) față de ceea ce Marsh numise anterior. [15] Reevaluarea de către oamenii de știință ulteriori a susținut cazul lui Marsh și a constatat că afirmația lui Cope că P. umbrosus era o specie mai mare și mai distinctă era incorectă. [8]
Un pterosaur fără dinți

În timp ce primele oase de aripă Pteranodon au fost colectate de Marsh și Cope la începutul anilor 1870, primul craniu Pteranodon a fost găsit la 2 mai 1876, de-a lungul râului Smoky Hill, în județul Wallace (acum județul Logan), Kansas , SUA, de Samuel Wendell Williston, un colecționar de fosile care a lucrat pentru Marsh. [1] Un al doilea craniu mai mic a fost descoperit curând. Aceste cranii au arătat că pterozaurii nord-americani erau diferiți de orice specie europeană prin faptul că nu aveau dinți și aveau creste osoase mari pe cap. Marsh a recunoscut această mare diferență, descriind exemplarele ca fiind „distincte de toate genurile cunoscute anterior din ordinul Pterosauriei prin întreaga absență a dinților”. Marsh a recunoscut că această caracteristică garantează un nou gen și, astfel, a inventat numele Pteranodon (care înseamnă „aripă fără dinți”) în 1876. Marsh a reclasificat toate speciile nord-americane denumite anterior ca Pterodactylus în Pteranodon . Marsh a atribuit cel mai mic craniu speciei Pteranodon occidentalis , în funcție de mărime, și a atribuit cel mai mare craniu, YPM 1117, noii specii Pteranodon longiceps , despre care credea că este o specie de dimensiuni medii între P. occidentalis mic și P mare ingeni . [16] [8] Mlaștina a numit alte câteva specii: Pteranodon comptus și Pteranodon nanus au fost numite pe baza scheletelor fragmentare ale indivizilor mici, în timp ce Pteranodon gracilis se baza pe un os de aripă pe care îl confundase cu un os pelvian. Curând și-a dat seama de greșeala sa și a reclasificat exemplarul înapoi într-un gen separat, pe care l-a numit Nyctosaurus . P. nanus a fost ulterior recunoscut și ca specimen de Nyctosaurus . [17] [8]
În 1892, Samuel Williston a examinat problema clasificării lui Pteranodon . El a menționat că, în 1871, Seeley menționase existența unui set parțial de fălci de pterosaur fără dinți din Cambridge Greensand din Anglia , pe care el le-a numit „Ornithostoma”. Întrucât principala caracteristică pe care Marsh o folosise pentru a separa Pteranodon de alți pterosauri era lipsa de dinți, Williston a concluzionat că „ Ornithostoma ” trebuie considerat sinonimul principal pentru Pteranodon . Tuttavia, nel 1901, Pleininger fece notare che "Ornithostoma" non era mai stato descritto scientificamente né aveva nemmeno assegnato un nome specifico fino al lavoro di Williston, e quindi era stato un nomen nudum e non poteva battere Pteranodon per priorità di denominazione. Williston accettò questa conclusione e tornò a chiamare il genere Pteranodon . [8] Tuttavia, sia Williston che Pleininger erano errati, poiché inosservato di entrambi era il fatto che, nel 1891, lo stesso Seeley aveva finalmente descritto e correttamente nominato Ornithostoma , assegnandolo alla specie O. sedgwicki . Nel 2010, ulteriori ricerche sull'identità di Ornithostoma hanno dimostrato che probabilmente Ornithostoma non era uno Pteranodon e non era neppure un suo parente stretto, ma potrebbe essere stato in realtà un azhdarchoide , un diverso tipo di pterosauro sdentato. [18]
Revisione delle specie
Williston è stato anche il primo scienziato a valutare criticamente tutte le specie di Pteranodon classificate da Cope e Marsh. Williston concordò con la maggior parte della classificazione di Marsh, con alcune eccezioni. In primo luogo, non credeva che P. ingens e P. umbrosus potessero essere considerati sinonimi, cosa che persino Cope credeva. Considerava specie dubbie sia P. velox che P. longiceps ; il primo era basato su frammenti non diagnostici e il secondo, sebbene noto da un cranio completo, probabilmente apparteneva a un'altra delle specie precedentemente nominate. Nel 1903, Williston rivisitò la questione della classificazione di Pteranodon , e rivise la sua precedente conclusione, portando il numero di specie da sette a tre. Considerò sia P. comptus che P. nanus esemplari di Nyctosaurus , e divise gli altri in specie piccole ( P. velox ), medie ( P. occidentalis ) e grandi ( P. ingens ), basate principalmente sulla forma delle ossa del braccio. Pensava che P. longiceps , l'unic< specie conosciuta da un cranio, potesse essere sinonimo di P. velox o P. occidentalis , in base alle dimensioni. [8]
Nel 1910, Eaton divenne il primo scienziato a pubblicare una descrizione dettagliata dell'intero scheletro di Pteranodon , come era noto all'epoca. Eaton usò i suoi risultati per rivedere la classificazione del genere ancora una volta sulla base di una migliore comprensione delle differenze nell'anatomia di Pteranodon , conducendo esperimenti usando modelli di ossa di argilla per aiutarsi a determinare gli effetti di schiacciamento e appiattimento sulle forme delle ossa del braccio che Williston aveva usato nella sua stessa classificazione. Eaton scoprì che la maggior parte delle differenze nelle forme ossee potrebbe essere facilmente spiegata dalle pressioni della fossilizzazione e concluso che nessuno degli scheletri di Pteranodon mostrava differenze significative tra loro oltre alle dimensioni. Pertanto, Eaton ideò un nuovo schema di classificazione che si basava sulle sole differenze dei crani, che assegnò alle varie specie, proprio come fece Marsh in base alle dimensioni. Alla fine, Eaton riconobbe solo tre specie valide: P. occidentalis , P. ingens e P. longiceps . [8]
La scoperta di esemplari con creste verticali, classificate da Harksen nel 1966 come la nuova specie Pteranodon sternbergi , complicò ulteriormente la situazione, spingendo un'altra revisione del genere da parte di Miller nel 1972. Perché era impossibile determinare la forma della cresta per tutte le specie sulla base di scheletri privi di cranio, Miller concluse che tutte le specie di Pteranodon , tranne le due basate su teschi ( P. longiceps e P. sternbergi ), dovevano essere considerate nomena dubia e abbandonate. Il cranio che Eaton pensava appartenesse a P. ingens fu collocato nella nuova specie Pteranodon marshi , e il cranio che Eaton aveva assegnato a P. occidentalis fu rinominato Pteranodon eatoni . Miller riconobbe anche un'altra specie basata su un cranio con una cresta simile a quella di P. sternbergi ; Miller chiamò questa specie Pteranodon walkeri . Per aiutare a mettere ordine in questo groviglio di nomi, Miller creò tre categorie o "sottogeneri" per loro. P. marshi e P. longiceps sono stati collocati nel sottogenere Longicepia , anche se questo è stato successivamente cambiato semplicemente in Pteranodon a causa delle regole di priorità. P. sternbergi e P. walkeri , le specie a cresta verticale, ricevettero il sottogenere Sternbergia , che in seguito fu cambiato in Geosternbergia perché Sternbergia era un nome già in uso ("preoccupato"). Infine, Miller nominò il sottogenere Occidentalia per P. eatoni , il cranio precedentemente associato a P. occidentalis . Miller ha ulteriormente ampliato il concetto di Pteranodon per includere Nyctosaurus come quarto sottogenere. Miller li considerava una progressione evolutiva, con il primitivo Nyctosaurus , all'epoca ritenuto un genere senza cresta, che aveva dato origine agli Occidentalia (dalla cresta più piccola), che a sua volta diede origine a Pteranodon con la sua lunga cresta all'indietro, portando infine a Geosternbergia con la sua grande cresta verticale. Tuttavia, Miller fece diversi errori nel suo studio riguardo a quali esemplari Marsh aveva assegnato a quali specie, e la maggior parte degli scienziati ignorò il suo lavoro sull'argomento nelle loro ricerche successive, sebbene Wellnhofer (1978) seguì l'elenco delle specie di Miller, e Schoch (1984) pubblicò in qualche modo un'altra revisione che essenzialmente è tornata allo schema di classificazione originale di Marsh, in particolare affondando P. longiceps come sinonimo di P. ingens . [8]
Riconoscimento delle variazioni
All'inizio degli anni '90, S. Christopher Bennett pubblicò diversi importanti articoli che analizzavano l'anatomia, la tassonomia e la storia della vita di Pteranodon . [5]
Fossili frammentari assegnati a Pteranodon sono stati scoperti anche a Scania , in Svezia . [19]
Paleobiologia
Volo

La forma dell'ala dello Pteranodon suggerisce che questo animale avesse uno stile di volo simile a quello di un moderno albatros . Ciò si basa sul fatto che lo Pteranodon aveva un rapporto di aspetto elevato (apertura alare per la lunghezza della corda alare) molto simile a quello di un albatros, circa 9:1 per lo Pteranodon , rispetto all' 8:1 dell'albatros. Gli albatros trascorrono lunghi periodi in mare per pescare ed usano un modello di volo chiamato "veleggiamento dinamico" che sfrutta il gradiente verticale della velocità del vento vicino alla superficie dell'oceano per percorrere lunghe distanze senza battere le ali, e senza l'ausilio di correnti termiche , assenti in oceano aperto. [20] Anche se gran parte del volo dell'animale si sarebbe basato sul veleggiamento, come avviene per gli uccelli marini con ali lunghe, il volo dello Pteranodon richiedeva comunque un occasionale battito d'ali attivo, e studi sul carico alare (la forza delle ali rispetto al peso del corpo) indicano che l'animale era in grado di volare in modo sostanziale sbattendo attivamente le ali, contrariamente ad alcune ipotesi precedenti secondo cui l'animale era così grande che avrebbe potuto solamente planare. [21]
Come molti altri pterosauri, Pteranodon , probabilmente, decollava in una posizione quadrupede, usando i loro lunghi arti anteriori come leva, per lanciarsi in aria con un balzo in avanti seguito da un rapido battito d'ali per prendere quota. Quasi tutta l'energia sarebbe stata generata dagli arti anteriori. Le ali si sarebbero spiegate solo quando l'animale si fosse sollevato abbastanza dal terreno e avesse completato il lancio in aria. [21]
Locomozione terrestre

Storicamente, la discussione sulla locomozione terrestre di Pteranodon , in particolare se l'animale fosse bipede o quadrupede , così come la locomozione terrestre dagli pterosauri in generale, è stato oggetto di dibattito.
Attualmente la maggior parte dei ricercatori sostiene che gli pterosauri fossero animali quadrupedi, spostandosi utilizzando tutti e quattro gli arti, e ripiegando le ali dietro il corpo, durante la locomozione a terra. Ciò è stato confermato anche dal ritrovamento di piste di impronte attribuite a pterosauri. [22]
La possibilità di una locomozione acquatica è stata discussa in diverse pubblicazioni, in particolare negli studi effettuati dal paleontologo Christopher Bennett (2001), che evidenziarono la netta somiglianza degli arti posteriori di Pteranodon con quelli dell' ibis rosso (somiglianze trovate nelle dita I, III e IV) e dei gabbiani (somiglianze trovate nelle dita II e IV). [23]
Al contrario gli arti di Pteranodon risultavano diversi nella morfologia da quelli di altre specie prese in esame, come quella del pavone , dell'estinta alca impenne e del coccodrillo marino . [23]
Bennett ritenne fortemente probabile che lo Pteranodon fosse capace di nuotare e che la sua struttura fisica fosse idonea perfino a brevi immersioni subacquee, in modo analogo allo stile di caccia delle moderne sule e pellicani . [24] .
Dieta e caccia

È noto che la dieta di Pteranodon fosse costituita in gran parte da pesce ; diverse ossa di pesce fossilizzate sono state ritrovate nella regione dello stomaco di uno Pteranodon , e un bolo di pesce fossilizzato è ristato trovato tra le fauci dell'esemplare AMNH 5098. Numerosi altri esemplari conservano frammenti di squame e vertebre nella zona del torso, indicando che il pesce costituiva la maggior parte della dieta di questo animale, sebbene sia possibile si possa essere cibato anche di molluschi o altri vertebrati marini [25]
Tradizionalmente, la maggior parte dei ricercatori ipotizzava che lo Pteranodon catturasse il pesce immergendo il becco in acqua mentre era in volo radente sul mare. Tuttavia, questa si ipotesi si basato sul presupposto che questi animali non potessero decollare dalla superficie dell'acqua. [26] Tuttavia, è molto probabile che Pteranodon fosse in grado di decollare dall'acqua, e che potesse immergersi per pescare. Anche un piccolo Pteranodon femmina avrebbe potuto raggiungere una profondità di almeno 80 centimetri (31 pollici) con il becco e il collo mentre il resto del corpo galleggiava sulla superficie, e potrebbe aver raggiunto profondità anche maggiori tuffandosi in acqua dall'aria come alcuni moderni uccelli marini dalle ali lunghe. [25] Nel 1994, Bennett evidenziò che la testa, il collo e le spalle di Pteranodon avevano una costituzione robusta, proprio molti uccelli subacquei. Ciò suggerisce che l'animale era in grado di tuffarsi da grandi altezze e raggiungere anche grandi profondità, ripiegando le ali come le moderne sule . [24]
Funzione della cresta
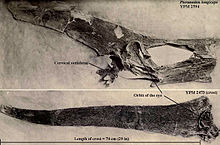
Una delle caratteristiche più note di Pteranodon è la sua iconica cresta cranica, sebbene la sua funzione sia ancora oggetto di dibattito. La maggior parte dei dibattiti si è concentrata sulla forma a lama della cresta, con la punta all'indietro nei maschi di P. longiceps , ignorando però l'ampia gamma di variazioni individuali di età e sesso. Il fatto che le creste varino così tanto tra individui della stessa specie esclude la maggior parte delle funzioni pratiche diverse da quelle utilizzate in display d'accoppiamento. [27] Pertanto, la funzione principale della cresta era probabilmente quella di display per attrarre il partner, mentre tutte le altre funzioni erano secondarie. [9]
Le possibili interpretazioni scientifiche della funzione della cresta iniziarono nel 1910, quando George Francis Eaton propose due possibilità: un contrappeso aerodinamico e/o un punto di attacco muscolare. Eaton ipotizzò che la cresta potesse essere un punto di ancoraggio per grandi e lunghi muscoli mascellari, ma ammise che questa funzione da sola non poteva spiegare il perché delle grandi dimensioni di alcune creste. [28] Bennett (1992) concordò con la stessa valutazione di Eaton che la cresta era troppo grande e variabile per essere un sito di attacco muscolare. [9] Eaton aveva ipotizzato che una possibile funzione secondaria della cresta era di contrappeso al lungo becco, riducendo la necessità di pesanti muscoli del collo per controllare l'orientamento della testa. [28] Alcuni test effettuati nella galleria del vento hanno dimostrato che la cresta funzionava effettivamente in qualche modo come un efficace contrappeso, ma Bennett notò che, ancora una volta, l'ipotesi si concentra solo sulle lunghe creste dei maschi di P. longiceps , e non sulle più grandi creste di P. sternbergi o su quelle più piccole delle femmine. Bennett scoprì che le creste delle femmine non avevano alcun effetto di controbilanciamento e che le creste dei maschio di P. sternbergi avrebbero, da sole, avuto un effetto negativo sull'equilibrio della testa. In effetti, il movimento laterale delle creste avrebbe richiesto un'elevata muscolatura del collo per controllare l'equilibrio della testa. [9]
Nel 1943, Dominik von Kripp ipotizzò che le cresta avrebbero potuto avere la funzione di timone , un'idea abbracciata da numerosi ricercatori successivi. [9] [29] Ross S. Stein ha persino suggerito che la cresta potesse essere sostenuta da una membrana di pelle che collegava la cresta rivolta all'indietro al collo e alla schiena, aumentando la sua superficie ed efficacia come un timone. [30] L'ipotesi del timone, ancora una volta, non tiene conto degli individui femminili né di P. sternbergi , che aveva una cresta rivolta verso l'alto, non rivolta all'indietro. Bennett scoprì anche che, anche nella sua qualità di timone, la cresta non avrebbe fornito la stessa forza direzionale della semplice manovra delle ali. L'ipotesi che la cresta fosse un freno ad aria e che gli animali volgessero la testa di lato per rallentare, soffre di un problema simile. [31] Inoltre, le ipotesi del timone e del freno ad aria non spiegano perché esista una variazione così grande nella dimensione della cresta anche tra gli adulti. [9]
Alexander Kellner suggerì che le grandi creste dello pterosauro Tupandactylus , così come altre specie, potrebbero essere state utilizzate per lo scambio di calore, consentendo a questi pterosauri di assorbire o disperdere calore e regolare la propria temperatura corporea, il che spiegherebbe anche la correlazione tra dimensione della cresta e taglia corporea. Tuttavia, non ci sono prove di vasi sanguigni nella cresta per questo scopo, tuttavia, e le grandi ali membranose piene di vasi sanguigni avrebbero svolto tale compito in modo molto più efficace. [9]
Con queste ipotesi escluse, l'ipotesi meglio supportata per la funzione della cresta sembra essere quella di display sessuale. Ciò è coerente con la variazione dimensionale osservata negli esemplari fossili, dove femmine e giovani hanno creste piccole o addirittura assenti ei maschi hanno creste grandi, elaborate e variabili. [9]
Variazione sessuale

Gli esemplari adulti di Pteranodon possono essere divisi in due classi di dimensioni distinte, piccola e grande, con la classe degli esemplari più grandi, che è circa una volta e mezza più grande della classe più piccola, e la classe degli esemplari più piccoli, che è circa due volte più comune della classe degli esemplari più grandi. Entrambe le classi di dimensioni vivevano fianco a fianco e sebbene inizialmente i ricercatori avevano ipotizzato che si trattassero di specie diverse, Christopher Bennett ha dimostrato che le differenze tra loro sono coerenti con il concetto che rappresentino esemplari femmine e maschi e che le specie di Pteranodon erano sessualmente dimorfiche . I crani della classe di dimensioni maggiori possiedono creste grandi, rivolte verso l'alto e/o all'indietro, mentre le creste della classe di dimensioni più piccole sono piccole e triangolari. Alcuni crani più grandi mostrano anche la prova di una seconda cresta che si estendeva in lungo e in basso, verso la punta del becco, che non si trova in esemplari più piccoli. [9]
Il sesso delle diverse classi di dimensioni era determinato non dai crani, ma dalle ossa pelviche. Contrariamente a quanto ci si potrebbe aspettare, la classe di dimensioni più piccole aveva ossa pelviche sproporzionatamente grandi e larghe. Bennett interpretò ciò come un canale del parto più spazioso, attraverso il quale passavano le uova. Bennett concluse quindi che la classe di piccole dimensioni con creste piccole e triangolari rappresentino esemplari femmine e gli esemplari più grandi con cresta grandi ed elaborate rappresentano esemplari maschi. [9]
Si noti anche che le dimensioni complessive e le dimensioni della cresta corrispondono anche all'età dell'individuo. Sono noti esemplari giovani sia femmine che maschi, ei maschi più giovani presentano spesso piccole creste simili a quelle delle femmine adulte. Pertanto, sembra che le grandi creste si siano sviluppate nei maschi solo quando raggiungevano le loro dimensioni adulte, rendendo difficile stabilire il sesso di esemplari immaturi da resti parziali. [32]
Il fatto che il numero delle femmine superi quello dei maschi, di due a uno, suggerisce che, come negli animali moderni con dimorfismo sessuale correlato alle dimensioni, come i leoni marini e altri pinnipedi , Pteranodon era probabilmente poligamo , con alcuni maschi che competevano per assicurarsi il dominio su un gran numero di femmine. In modo simile ai pinnipedi moderni, i maschi Pteranodon potrebbero aver gareggiato per stabilire un proprio territorio sulle scogliere o sulle spiagge, con i maschi più grandi e con la cresta più grande che occupavano un maggior territorio e avevano più possibilità di accoppiandosi con le femmine. Le creste dei maschio di Pteranodon non sarebbe state usate in combattimento, ma piuttosto come "simboli visivi di rango dominante", con rituali di esibizione che sostituivano la competizione fisica con altri maschi. Se questa ipotesi è corretta, è anche probabile che i maschi di Pteranodon non prendevano parte nell'allevamento dei piccoli; un simile comportamento non si riscontra nei maschi dei moderni animali poligami che generano più prole allo stesso tempo. [9] Tuttavia, i giovani pterosauri sono noti per precoci, ed essere indipendenti e capaci di volare sin dalla schiusa, pertanto non era necessario l'aiuto di nessuno dei genitori. [33]
Paleoecologia

Gli esemplari fossili assegnati a Pteranodon sono stati ritrovati sia nei depositi di Smoky Hill Chalk della Formazione Niobrara, sia nei depositi di Sharon Springs leggermente più recenti della Formazione di Pierre Shale. All'epoca di Pteranodon , questa zona era coperta da un grande mare interno, noto come Mare interno occidentale . Famose per i fossili raccolti dal 1870, queste formazioni si estendono dal sud del Kansas , negli Stati Uniti, fino a Manitoba , in Canada. Tuttavia, gli esemplari di Pteranodon (o eventuali esemplari di altri pterosauri) sono stati ritrovati solo nella metà meridionale della formazione, in Kansas , Wyoming e South Dakota . Nonostante il fatto che numerosi fossili siano stati trovati nelle parti contemporanee della formazione in Canada, nessun esemplare di pterosauro è mai stato trovato lì. Ciò suggerisce che la gamma geografica naturale di Pteranodon copriva solo la parte meridionale della Formazione Niobrara e che il suo habitat non si estendeva più a nord del South Dakota. [8]
Alcuni fossili molto frammentari appartenenti ai pterosauri pteranodonti, e forse allo stesso Pteranodon , sono stati ritrovati anche sulla Costa del Golfo e sulla costa orientale degli Stati Uniti . Ad esempio, alcuni frammenti di ossa della Formazione Mooreville, in Alabama , e la Formazione Merchantville del Delaware potrebbero appartenere a Pteranodon , anche se questi fossili sono troppo frammentari per dare un'identificazione precisa. [8] Anche alcuni resti dal Giappone sono stati provvisoriamente attribuiti a Pteranodon , ma la loro distanza geografica dall'habitat dell'animale rende improbabile questa identificazione. [8]

Pteranodon longiceps condivideva il cielo con lo pterosauro Nyctosaurus , dalla grande cresta. Rispetto a P. longiceps , che era una specie molto comune, Nyctosaurus era più raro, costituendo solo il 3% dei fossili di pterosauro dalla formazione. Anche meno comune era l'uccello dentato, Ichthyornis . [10]
È probabile che, come in altri animali poligami (in cui i maschi competono per il maggior numero di femmine nel proprio harem), Pteranodon viveva principalmente sulle scogliere o isole rocciose al largo, dove poteva nidificare lontano dai predatori terrestri e nutrirsi lontano dalla costa; la maggior parte dei fossili di Pteranodon si trovano in luoghi che all'epoca erano a centinaia di chilometri dalla costa. [9]
Sotto la superficie, il mare era popolato principalmente da invertebrati come ammoniti e calamari . I vertebrati marini erano estremamente comuni ed, oltre ai pesci, comprendevano tartarughe marine , come Toxochelys , il plesiosauro Styxosaurus e l'uccello marino incapace di volare Parahesperornis . I mosasauri erano i rettili marini più comuni, con generi tra cui Clidastes e Tylosaurus . [1] Alcuni fossili eccezionali ci dimostrano che almeno alcuni di questi rettili marini si sono nutriti di Pteranodon . Barnum Brown , nel 1904, riportò lo scoperta del contenuto dello stomaco di un plesiosauro che conteneva ossa di "pterosauro", molto probabilmente uno Pteranodon . [36]
All'interno dei gessi della Formazione Niobrara sono stati ritrovati anche i resti di dinosauri terrestri, suggerendo che gli animali morti sulla riva venivano portati via dal mare o trascinati dai fiumi, dove le loro carcasse venivano consumate da organismi marini (come nel caso di un esemplare di adrosauro che mostra segni di morsi di uno squalo ). [37]
Periodo di vita ed evoluzione
In termini generali, lo Pteranodon sarebbe vissuto nella fine degli anni del Coniaciano ai primi anni delle fasi del Cretaceo . Però, gli anni precisi in cui è vissuto non sono ancora stati del tutto confermati; nel 2003 , Kenneth Carpenter ha esaminato la distribuzione e la datazione dei fossili, dimostrando che lo Pteranodon esisteva 88-85 milioni di anni fa, mentre Pteranodon Longiceps esisteva tra i 86 e 84,5 milioni di anni fa. Invece nel 2010 , la formazione Pierre Shale in Kansas , Wyoming e Dakota del Sud , ha ipotizzato che il noto rettile preistorico sia risalente tra 81,5 e 80,5 milioni di anni fa.
Nei primi anni del 1990 , sono stati notati che il principale Polimorfismo dello Pteranodon si è separata in poco tempo. Ciò ha suggerito, insieme alla loro somiglianza globale, che probabilmente rappresentano all'interno un unico ceppo evolutivo della durata di circa 4 milioni di anni. In altre parole, una sola specie di Pteranodon sarebbero stati presenti in un dato momento, e lo Pteranodon sternbergi con ogni probabilità è stato l'antenato diretto dello Pteranodon longiceps .
Nella cultura di massa
- Nel film King Kong uno Pteranodon rapisce Ann Darrow e viene affrontato e ucciso da Kong. Lo pteranodonte compare con ruoli simili anche in Un milione di anni fa dove si scontra con un Rhamphorhynchus gigantesco, e La vendetta di Gwangi .
- Nei film animati della saga " Alla ricerca della Valle Incantata " uno Pteranodon di nome Petrie è uno dei personaggi principali.
- Nella serie animata Il treno dei dinosauri i protagonisti sono pteranodonti.
- Nel film Rodan, il mostro alato fa il suo debutto Rodan, uno Pteranodon modificato dalle radiazioni, che in seguito diventerà uno dei più famosi mostri del cinema giapponese.
- Gli pteranodonti appaiono in Il mondo perduto - Jurassic Park , Jurassic Park III , Jurassic World e Jurassic World - Il regno distrutto .
- Il Pokémon Aerodactyl è basato sul pteranodonte.
- King , personaggio del manga e anime One Piece , ha la capacità di trasformarsi in uno Pteranodon grazie ai poteri del suo frutto del diavolo.
- Degli pteranodonti appaiono in Fantasia .
- Appare come mostro nelle serie animate The Scooby-Doo Show e Be Cool, Scooby-Doo! e nel film Scooby-Doo 2 - Mostri scatenati
- Alcune ricostruzioni di pteranodonte sono presenti in diversi parchi tematici tra cui: il Parco della Preistoria di Rivolta d'Adda ( CR ), il Parco dei Dinosauri di Castellana Grotte ( BA ), il Parco Natura Viva di Bussolengo ( VR ) e il parco "Il Mondo della Preistoria" di Simbario ( VV )
Note
- ^ a b c d Bennett, SC (2000). "Inferring stratigraphic position of fossil vertebrates from the Niobrara Chalk of western Kansas." Current Research in Earth Sciences: Kansas Geological Survey Bulletin , 244 (Part 1): 26 pp.
- ^ a b c d e SC Bennett, The Pterosaurs of the Niobrara Chalk , in The Earth Scientist , vol. 11, n. 1, 1994, pp. 22-25.
- ^ a b MP Witton e MB Habib, On the Size and Flight Diversity of Giant Pterosaurs, the Use of Birds as Pterosaur Analogues and Comments on Pterosaur Flightlessness , in PLoS ONE , vol. 5, n. 11, 2010, p. e13982, Bibcode : 2010PLoSO...513982W , DOI : 10.1371/journal.pone.0013982 , PMC 2981443 , PMID 21085624 .
- ^ a b Bennett, SC (1987). "New evidence on the tail of the pterosaur Pteranodon (Archosauria: Pterosauria)." Pp. 18–23 in Currie, PJ and EH Koster (eds.), Fourth Symposium on Mesozoic Terrestrial Ecosystems, Short Papers. Occasional Papers of the Tyrrell Museum of Paleontology, #3.
- ^ a b c d Kellner, AWA, Comments on the Pteranodontidae (Pterosauria, Pterodactyloidea) with the description of two new species ( PDF ), in Anais da Academia Brasileira de Ciências , vol. 82, n. 4, 2010, pp. 1063-1084, DOI :10.1590/S0001-37652010000400025 .
- ^ Martin-Silverstone, E., Glasier, J., Acorn, J., Mohr, S., and Currie, P. 2017. Reassesment of Dawndraco kanzai Kellner, 2010 and reassignment of the type specimen to Pteranodon sternbergi Harksen, 1966. Vertebrate Anatomy Morphology Palaeontology , 3 :47-59. DOI : 10.18435/B5059J
- ^ Piero e Alberto Angela, Rettili Volanti , in "Il pianeta dei dinosauri" , Arnoldo Mondadori Editore, 1993, p. 111, ISBN 88-04-41235-6 .
- ^ a b c d e f g h i j k l m n o p q r s Bennett, SC, Taxonomy and systematics of the Late Cretaceous pterosaur Pteranodon (Pterosauria, Pterodactyloida) , in Occasional Papers of the Natural History Museum, University of Kansas , vol. 169, 1994, pp. 1-70.
- ^ a b c d e f g h i j k l Bennett, SC, Sexual dimorphism of Pteranodon and other pterosaurs, with comments on cranial crests , in Journal of Vertebrate Paleontology , vol. 12, n. 4, 1992, pp. 422-434, DOI : 10.1080/02724634.1992.10011472 .
- ^ a b K Carpenter, Vertebrate Biostratigraphy of the Smoky Hill Chalk (Niobrara Formation) and the Sharon Springs Member (Pierre Shale) , in High-Resolution Approaches in Stratigraphic Paleontology , vol. 21, 2003, pp. 421-437, DOI : 10.1007/978-1-4020-9053-0 .
- ^ Zimmerman, H., Preiss, B., and Sovak, J. (2001). Beyond the Dinosaurs!: sky dragons, sea monsters, mega-mammals, and other prehistoric beasts , Simon and Schuster. ISBN 0-689-84113-2 .
- ^ B. Andres e TS Myers, Lone Star Pterosaurs , in Earth and Environmental Science Transactions of the Royal Society of Edinburgh , vol. 103, 2013, p. 1, DOI : 10.1017/S1755691013000303 .
- ^ B. Andres, J. Clark e X. Xu, The Earliest Pterodactyloid and the Origin of the Group , in Current Biology , vol. 24, 2014, pp. 1011-6, DOI : 10.1016/j.cub.2014.03.030 , PMID 24768054 .
- ^ a b Peter Wellnhofer, The Illustrated Encyclopedia of Pterosaurs , New York, Barnes and Noble Books, 1996 [1991] , p. 139, ISBN 0-7607-0154-7 .
- ^ Cope, ED (1875). "The Vertebrata of the Cretaceous formations of the West." Report, US Geological Survey of the Territories (Hayden) , 2 : 302 pp., 57 pls.
- ^ Marsh, OC, Notice of a new sub-order of Pterosauria , in American Journal of Science , Series 3, vol. 11, n. 65, 1876a, pp. 507-509, DOI : 10.2475/ajs.s3-11.66.507 .
- ^ Marsh, OC, Principal characters of American pterodactyls , in American Journal of Science , Series 3, vol. 12, n. 72, 1876b, pp. 479-480, DOI : 10.2475/ajs.s3-12.72.479 .
- ^ AO Averianov, Ornithostoma sedgwicki – valid taxon of azhdarchoid pterosaurs , in Proceedings of the Zoological Institute RAS , vol. 316, n. 1, 2012, pp. 40-49.
- ^ Allt om Vetenskap, 10-2012, p.72
- ^ Padian, K. (1983), "A functional analysis of flying and walking in pterosaurs". , in Paleobiology. , 9 (3), 218–239.
- ^ a b Witton, MP; Habib, MB (2010). "On the Size and Flight Diversity of Giant Pterosaurs, the Use of Birds as Pterosaur Analogues and Comments on Pterosaur Flightlessness" , su journals.plos.org .
- ^ Unwin, David M., The Pterosaurs: From Deep Time , New York: Pi Press, 2006, pp. 210 -222, ISBN 978-0-13-146308-0 .
- ^ a b CLAW MORPHOLOGY OF PTERANODON AND POSSIBLE AQUATIC LOCOMOTION , su gsa.confex.com . URL consultato il 15 aprile 2019 (archiviato dall' url originale il 7 gennaio 2011) .
- ^ a b Bennett, SC (1994), "The Pterosaurs of the Niobrara Chalk" , in The Earth Scientist. , vol. 11, (1): 22–25.
- ^ a b Bennett, SC (1994), "The Pterosaurs of the Niobrara Chalk" , in The Earth Scientist. , vol. 11, (1): 22-25.
- ^ Bennett, SC (1994), "The Pterosaurs of the Niobrara Chalk" , in The Earth Scientist , vol. 11, (1): 22-25.
- ^ JL Tomkins, NR Lebas, MP Witton, DM Martill e S. Humphries, Positive Allometry and the Prehistory of Sexual Selection ( PDF ), in The American Naturalist , vol. 176, n. 2, 2010, pp. 141-148, DOI : 10.1086/653001 , PMID 20565262 .
- ^ a b Eaton, GF (1910). "Osteology of Pteranodon ." Memoirs of the Connecticut Academy of Arts and Sciences , 2 :1–38, pls. i–xxxi.
- ^ von Kripp, D. (1943). "Ein Lebensbild von Pteranodon ingens auf flugtechnischer Grundlage." Nova Acta Leopoldina, NF , 12 (83): 16–32 [in German].
- ^ Stein, RS, Dynamic analysis of Pteranodon ingens : a reptilian adaptation to flight , in Journal of Paleontology , vol. 49, 1975, pp. 534-548.
- ^ Bramwell, CD and Whitfield, GR (1974). "Biomechanics of Pteranodon." Philosophical Transactions Royal Society B , 267 .
- ^ Bennett, SC, The osteology and functional morphology of the Late Cretaceous pterosaur Pteranodon . General description of osteology , in Palaeontographica, Abteilung A , vol. 260, 2001, pp. 1-112.
- ^ Xiaolin Wang, Alexander WA Kellner, Shunxing Jiang, Qiang Wang, Yingxia Ma, Yahefujiang Paidoula, Xin Cheng, Taissa Rodrigues, Xi Meng, Jialiang Zhang, Ning Li e Zhonghe Zhou, Sexually dimorphic tridimensionally preserved pterosaurs and their eggs from China , in Current Biology , vol. 24, n. 12, 2014, pp. 1323-1330, DOI : 10.1016/j.cub.2014.04.054 , PMID 24909325 .
- ^ vertebre cervicali di un pteradonte con un dente di squalo conficcato in una di esse
- ^ http://markwitton-com.blogspot.com/2018/12/no-cretaceous-sharks-did-not-leap-from.html
- ^ Brown, B., Stomach stones and the food of plesiosaurs , in Science , vol. 20, n. 501, 1904, pp. 184-185, Bibcode : 1904Sci....20..184B , DOI : 10.1126/science.20.501.184 , PMID 17737868 .
- ^ MJ Everhart e K. Ewell, [27:sdhcvf 2.0.co;2 Shark-bitten dinosaur (Hadrosauridae) vertebrae from the Niobrara Chalk (Upper Coniacian) of western Kansas ], in Transactions of the Kansas Academy of Science , vol. 109, 1–2, 2006, pp. 27-35, DOI : 10.1660/0022-8443(2006)109[27:sdhcvf]2.0.co;2 .
Altri progetti
-
 Wikimedia Commons contiene immagini o altri file su Pteranodon
Wikimedia Commons contiene immagini o altri file su Pteranodon -
 Wikispecies contiene informazioni su Pteranodon
Wikispecies contiene informazioni su Pteranodon
Collegamenti esterni
- ( EN ) Pteranodon , su Enciclopedia Britannica , Encyclopædia Britannica, Inc.
- ( EN ) Pteranodon , su Fossilworks.org .
| Controllo di autorità | Thesaurus BNCF 59790 · LCCN ( EN ) sh85108544 · BNF ( FR ) cb125455988 (data) |
|---|