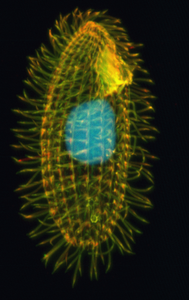
Tetrahimena
| Tetrahimena | |
|---|---|
 | |
| Clasificare științifică | |
| Domeniu | Eukaryota |
| Regatul | Protist |
| ( cladă ) | Chromalveolata |
| ( cladă ) | Alveolat |
| Phylum | Ciliophora |
| Clasă | Oligohimenoforeea |
| Ordin | Himenostomatida |
| Familie | Tetrahymenidae |
| Tip | Tetrahimena Furgason , 1940 |
Tetrahymena este un gen de protiști ciliați obișnuiți în apa dulce. Cele două specii tetrahymena thermophila și tetrahymena pyriformis sunt utilizate ca organisme model în cercetarea biomedicală. [1]
Spre deosebire de alte eucariote și la fel ca alte ciliate, tetrahimena are două tipuri diferite de nuclei ( dimorfism nuclear ), unul mic ( micronucleus ), un diploid utilizat pentru reproducere și unul mai mare ( macronucleus ) poliploid pentru funcționarea celulei. Macronucleul este produs de micronucleu prin mărirea genomului. Diviziunea sa are loc prin amitoză . Periodic macronucleul trebuie regenerat de micronucleus. În multe cazuri, acest lucru se întâmplă prin reproducerea sexuală care apare de obicei prin conjugare .

Organism model în biologia experimentală

Ca protozoar ciliate, Tetrahymena thermophila prezintă dimorfism nuclear : două tipuri de nuclee celulare. Au un macronucleu mai mare, fără linie germinativă și un micronucleu mai mic în fiecare celulă în același timp și ambele îndeplinesc funcții diferite cu proprietăți citologice și biologice distincte. Această versatilitate unică permite oamenilor de știință să utilizeze Tetrahymena pentru a identifica mai mulți factori cheie care privesc expresia genei și integritatea genomică. În plus, Tetrahymena posedă zeci de cili și are structuri microtubulare complicate, făcându-l un model optim pentru ilustrarea diversității și funcțiilor aranjamentelor microtubulare.
Deoarece Tetrahymena poate fi cultivată cu ușurință în cantități mari în laborator, a fost o sursă excelentă pentru analize biochimice de ani de zile, în special pentru activități enzimatice și pentru purificarea componentelor sub-celulare. Mai mult, odată cu avansarea tehnicilor genetice, a devenit un model excelent pentru studierea funcției genice in vivo . Secvențierea recentă a genomului macronucleului ar trebui să asigure că Tetrahymena va continua să fie utilizată ca sistem model.
Tetrahymena thermophila poate exista la 7 sexe diferite care se pot reproduce în 21 de combinații diferite, iar o singură tetrahymena nu se poate reproduce sexual cu ea însăși.
Studiile asupra Tetrahimenei au contribuit la mai multe etape științifice, inclusiv:
- Prima celulă care prezintă diviziune sincronizată, ceea ce a condus la primele înțelegeri ale existenței mecanismelor care controlează ciclul celular . [2]
- Identificarea și purificarea primei proteine motorii bazate pe citoschelet , cum ar fi dinina . [2]
- Ajutați la descoperirea lizozomilor și a peroxizomilor . [2]
- Prima identificare moleculară a rearanjării genomice somatice. [2]
- Descoperirea structurii moleculare a telomerilor , a enzimei telomerazei , a rolului telomerazei ARN ca șablon și a rolurilor acestora în senescența celulară și vindecarea cromozomială (pentru care a fost câștigat un premiu Nobel). [2]
- Co-descoperire acordată cu Premiul Nobel (1989, pentru chimie) a acidului ribonucleic catalitic ( ribozimă ). [2]
- Descoperirea funcției „ acetilării ” histonei . [2]
- Demonstrarea rolurilor modificării post-translaționale, cum ar fi acetilarea și glicilarea pe tubuline și descoperirea enzimelor responsabile de unele dintre aceste modificări (glutamilare).
- Structura cristalină a ribozomului 40S împreună cu factorul său de inițiere eIF1.
- Prima demonstrație că doi dintre codonii de oprire „universali”, UAA și UAG, vor codifica glutamina în unele eucariote , lăsând UGA ca singurul codon care se termină în aceste organisme. [3]
- Descoperirea autosplicării ARN . [4]
Notă
- ^ Alfred M. Elliott, Biology of Tetrahymena , Dowen, Hutchinson and Ross Inc., 1973, ISBN 0-87933-013-9 .
- ^ a b c d e f g Tetrahymena Genome Sequencing Paper White
- ^ Horowitz S și Gorovsky M, Un cod genetic neobișnuit în genele nucleare ale Tetrahimenei , în Proc. Natl. Acad. Sci. SUA , vol. 82, nr. 8, aprilie 1985, pp. 2452-2455, DOI : 10.1073 / pnas.82.8.2452 , PMC 397576 , PMID 3921962 .
- ^ K. Kruger, PJ Grabowski, AJ Zaug, J. Sands, DE Gottschling și TR Cech, ARN auto-splicing: Autoexcizie și autociclizare a ARN ribozomal intervenind secvența de tetrahimene , în Cell , vol. 31, 1982, p. 147, DOI : 10.1016 / 0092-8674 (82) 90414-7 .
Bibliografie
- ( EN ) JA Eisen și colab. , Macronuclear Genome Sequence of the Ciliate Tetrahymena thermophila, a Model Eukaryote , în PLoS Biology , vol. 4, nr. 9, 2006, pp. 1620-1642, DOI : 10.1371 / journal.pbio.0040286 .
- (EN) Methods in Cell Biology Volume 62: Tetrahymena thermophila, Editat de David J. Asai și James D. Forney. (2000). De Academic Press
- (EN) Collins, K. și Gorovsky, MA (2005). Tetrahymena thermophila Curr Biol. 15 : R317-8 [ conexiune întreruptă ] .
- ( EN ) Eisen JA, Coyne RS, Wu M, Wu D, Thiagarajan M, și colab. (2006) Secvența genomului macronuclear al ciliatei Tetrahymena thermophila, un model eucariot. PLoS Biol 4 (9): e286 , pe biology.plosjournals.org .
Alte proiecte
-
 Wikimedia Commons conține imagini sau alte fișiere despre Tetrahymena
Wikimedia Commons conține imagini sau alte fișiere despre Tetrahymena -
 Wikispeciile conțin informații despre Tetrahymena
Wikispeciile conțin informații despre Tetrahymena