Istoria genetică a Italiei
Acest articol sau secțiune ar trebui revizuit și actualizat cât mai curând posibil . |

Istoria genetică este ansamblul descoperirilor făcute prin intermediul geneticii populației , o ramură a geneticii care analizează constituția genetică a populațiilor mendeliene în termeni calitativi (variante alelice prezente într-o populație) și cantitative (frecvențe alelice și genotipice ). Aceste descoperiri au făcut posibilă, prin analiza înrudirii și a diferențelor genetice, atât la nivelul grupurilor etnice, cât și al populațiilor umane, reconstituirea fluxurilor migratorii, a traversărilor, a apariției sau a eliminării caracterelor care disting grupurile etnice actuale. și populațiile umane, atât în spațiu, cât și în timp, ajutând la reconstituirea istoriei omului de la apariția sa.
Note despre apariția omului modern și a doua ieșire din continentul african
Acum aproximativ șapte milioane de ani, stabilirea scoarței terestre a produs ridicarea Văii Riftului care traversează statele actuale din Etiopia, Kenya și Tanzania. Vânturile încărcate de ploaie din vest au fost prinse de ridicarea tectonică a unei bariere naturale, Riftul african, provocând un proces de uscare care a avut loc pe platoul cu același nume. Procesul de uscare a condus, de fapt, la formarea progresivă a savanei africane actuale pentru a înlocui pădurea, iar populația proto-maimuță africană s-a trezit geografic separată în două subpopulații de Rift: partea de vest a rămas luxuriantă și proto-maimuțele s-au așezat pe aceasta latură adaptată la un mediu împădurit, diferențiindu-se de celelalte și anticipând maimuțele antropomorfe moderne, în timp ce proto-maimuțele prinse pe platou, datorită uscării lente, adaptate la diferite condiții de mediu, cum ar fi dispariția pădurii înlocuită de cea africană savana . Acest tip de mediu a permis evoluția și afirmarea genului Homo [2] .
Afara din Africa
Procesul din afara Africii (literal: părăsirea Africii ) este primul proces de migrație cunoscut, chiar dacă, spre deosebire de fluxurile migratorii moderne, s-a datorat succesului primilor hominizi, care au putut să se extindă într-un mediu fără concurenți și deosebit de avantajos . Apartenența la genul Homo progenitor al speciei noastre, numită Homo ergaster , își are originea în Africa . Evoluția sa dă naștere la Homo erectus , iar colonizând Eurasia în valuri succesive se adaptează la diferite condiții de mediu, diferențându-se în specii Homo heidelbergensis și ulterior în Homo neanderthalensis cu caracteristici carnivore.
În afara Africii 2
Unii hominizi aparținând genului Homo, în loc să participe la procesul din afara Africii, s-au stabilit în Valea Riftului . Homo sapiens provine din aceste hominide, dintre care una dintre subspecii, definea Homo sapiens sapiens (acum aproximativ 200 000 de ani), a migrat pentru a se stabili ulterior ca singura specie aparținând genului Homo [3] , și înlocuindu-se, din motive încă pe scară largă. dezbătute, atât speciilor care au apărut în urma procesului din afara Africii, cât și celor co-evoluate în Africa.
Istoria populației (ADN-Y)
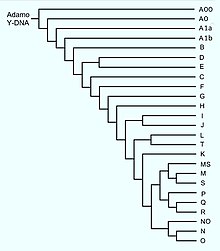
În genetică umană , cromozomul Y este împărțit în haplogrupuri definite pe baza mutației unui singur nucleotid [4] în secvența nerecombinantă a cromozomului Y numită NRY. [5] Fiecare mutație corespunde unui haplotip și cromozomul Y este moștenit de la tată la fiu. Deoarece mutația afectează o secvență non-recombinantă (adică nu suferă modificări atunci când este moștenită), este posibil să ne întoarcem, revenind din generație în generație, la descendența masculină.

Haplotipuri europene legate de Y-ADN
Studiul haplotipurilor sau combinația de variante alelice de -a lungul unui cromozom sau de-a lungul unui segment cromozomial care conține loci strâns asociați unul cu celălalt și care sunt în general moștenite împreună, ne-a permis identificarea a două haplotipuri europene definite Eu18 și Eu19 care au permis, prin metode de comparare a secvențelor, pentru a identifica urmele migrațiilor populațiilor europene care datează din epoca paleolitică, se crede că migrațiile populațiilor europene se datorează fenomenelor de mediu, cum ar fi glaciațiile, concurența dintre populații și căutarea hranei, fiind haplotipurile moștenite împreună, au fost identificate două nuclee de populație izolate, respectiv nucleele din Peninsula Iberică și nucleele din Ucraina, aceste haplotipuri reprezintă 50% din cromozomii Y europeni. Haplotipul Eu19 este de asemenea răspândit în nordul Pakistanului și în Asia centrală pentru a susține ipoteza că aceste două populații au migrat atât către Europa Centrală, cât și către Asia [6] .
Haplotipuri italiene legate de Y-ADN
Acest articol sau secțiune pe tema geneticii este considerat a fi verificat . |
Distribuția procentuală a haplotipurilor italiene ale cromozomului Y [7] .
| regiune | I1 | I2a | I2b | R1a | R1b | G2a | J2 | J1 | E1b1b | T. | Î |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Nordul Italiei | 7% | 1% | 3,5% | 4,5% | 49,5% | 7,5% | 10% | 1,5% | 11% | 2% | 0% |
| Toscana | 4% | 1,5% | 2,5% | 4% | 52,5% | 9% | 11,5% | 2% | 9% | 2% | 0% |
| centrul Italiei | 2,5% | 2% | 1,5% | 3% | 36% | 11% | 23% | 5% | 11,5% | 3% | 0% |
| sudul Italiei | 2,5% | 3,5% | 1% | 3% | 27,5% | 10,5% | 21,5% | 4% | 18,5% | 2,5% | 0% |
| Sicilia | 3,5% | 3% | 1% | 4,5% | 26% | 8,5% | 23% | 3,5% | 20,5% | 4% | 1% |
| Sardinia | 0% | 37% | 2% | 1% | 18,5% | 12% | 9% | 4% | 9,5% | 1,5% | 0% |
Istoricul populației (ADNmt)
În mod similar cu cromozomul Y, ADN-ul mitocondrial (ADNmt) conținut în mitocondrii este moștenit din partea maternă, permițându-ne astfel să urmărim calea patrilineală feminină , haplogrupul mitocondrial U5b3 ne-a permis să identificăm un efect fondator care a avut loc aproximativ 10.000 de ani acum în Italia, se crede că femelele purtătoare au migrat ulterior în Provence, probabil între 9000 și 7000 de ani în urmă, unde s-a dezvoltat varianta U5b3a1. Fenomenele de migrare ulterioare ar fi permis apoi introducerea haplotipului U5b3a1 din Provence în Sardinia, probabil în urma comerțului cu obsidian, astăzi aproximativ 4% din populația feminină din Sardinia aparține acestui haplotip. [8]
Haplotipuri legate de ADNmt
Cel mai frecvent haplogrup din Europa și Italia pare a fi haplogrupul H care a apărut probabil acum aproximativ 20.000 de ani în sudul Europei și în Orientul Apropiat, din nou în Europa în urmă cu aproximativ 15.000 de ani în Spania haplogrupul V diferă. Haplogrupul J se crede că a provenit din Orientul Apropiat sau din Caucaz, în timp ce în nord-est acum aproximativ 25.000 de ani este originar haplogrupul W, haplogrupul T provine din Mesopotamia acum aproximativ 17.000 de ani, haplogrupul U provine din Asia de Vest cu aproximativ 60.000 de ani în urmă, haplogrupul I cu aproximativ 30.000 de ani în urmă probabil în Europa, haplogrupul K cu aproximativ 16.000 de ani în urmă în Orientul Apropiat, haplogrupul X2 acum 30.000 de ani în Europa de Nord-Est [9] [10]
Tabel procentual de haplogrupuri femel ADNm prezente în Italia:
| H. | V. | J | T. | U | K. | THE | W | X2 | alții | |
|---|---|---|---|---|---|---|---|---|---|---|
| % | 33,5% | 4,5% | 7,5% | 12% | 12,5% | 7% | 2% | 2,5% | 2% | 16,5% |
Migrațiile umane detectate prin analiza ADN-ului Y
Paleolitic superior
Prin studii privind variația genetică a cromozomului Y în Europa, s-a găsit o perioadă timpurie de colonizare la sfârșitul erei glaciare, care a condus la diferențierea primelor haplotipuri europene asociate cu omul modern. Fiecare haplotip este de obicei asociat cu o populație care, după ce s-a stabilit pe teritoriu, a avut suficient timp pentru a se diferenția de populația inițială, lăsând loc propriului său haplotip caracteristic. Haplotipul I1a este mai prezent în Scandinavia, frecvența acestuia scade rapid deplasându-se spre câmpia de est și marginea Atlanticului, cu toate acestea diferențele constatate la nivelul microsateliților arată că probabil în sudul Franței este necesar să se caute zona de Originea populațiilor purtătoare de haplotipuri I1a și I1c. Haplotipul I1b se extinde de la estul Adriaticii până la estul Europei, scade în direcția sudului Balcanilor și dispare brusc în nordul Italiei. Haplotipul I și produsele secundare se presupune că au apărut în timpul ultimei glaciații din Balcani sau Europa de Est. Dimpotrivă, populația care transportă I1b2, cel mai probabil, își are originea în sudul Franței, suferind o expansiune în perioada post-glaciară și, ca urmare a fluxurilor migratorii, ar fi colonizat Sardinia acum aproximativ 9000 de ani [11] .
Mesolitic și Neolitic
În această perioadă există un grad apreciabil de expansiune a omului modern în peninsula italiană. Studiile actuale sunt foarte complexe datorită derivei genetice și a efectelor de fondare locală care au avut loc în toată peninsula în această perioadă de timp. Se crede că Haplogroup R1b (ADN-Y) este cea mai veche linie genetică prezentă pe continentul european [6] . Prezența sa este în mod obișnuit asociată cu un efect fondator care a avut loc în Europa Centrală și de Vest [12] [13] . Populațiile stabilite în Italia din mezolitic sunt caracterizate de frecvențe ridicate ale R1 * (xR1a1), o afecțiune care se găsește astăzi la populațiile basche, considerate cele mai similare genetic cu primii europeni. În timpul neoliticului, migranții au introdus variantele E3B1 și J2, 27% din variațiile genetice totale, pe baza analizei polimorfismelor, indică un gradient de distribuție clar al populației italiene pe axa nord-sud a peninsulei. Variațiile introduse în neolitic nu par a fi datorate fluxurilor migratorii din Spania, ci sunt configurate ca migrații din Asia sau Anatolia prin actuala zonă balcanică; mai mulți autori au sugerat că axa nord-sud de distribuție a diferențelor genetice între populațiile italiene se datorează evenimentelor colonizării grecești din sud. Cu toate acestea, noi studii sugerează că în epoca neolitică influența populațiilor din Anatolia a fost cauza principală a diferențelor din bazinul genetic italian, atribuind grecilor un rol de importanță secundară; în prezent se presupune că în timpul neoliticului haplotipul principal R1 * (xR1a1) a fost consolidat în timp ce haplotipurile HGS, E3B1 și J2 sunt absente sau prezente la frecvență scăzută. În special, în nordul Italiei, E3b2 de origine africană se găsește la o frecvență foarte mică [14] .
R1b-S28 (U152)
Distribuția variației STR a R1b în Europa scade de la est la vest, sugerând o intrare în Europa din vestul Asiei odată cu răspândirea agriculturii [15] . R1b este adesea citat ca exemplu de distribuție a valurilor haplogrupului, în acest caz, de la est la vest. [16] . Propunerea pentru o origine sud-estică a R1b își are originea în descoperirea primelor subclade ale R1b situate în Asia de Vest și cele mai recente din Europa de Vest [17] [18] . Cu toate acestea, nu există un acord cu privire la datarea migrației sau a migrațiilor responsabile de această distribuție, migrațiile înainte sau după neolitic nu sunt excluse [17] .
David K. Faux a emis ipoteza că mutația S28 (markerul U152) și subgrupurile înrudite pot fi asociate într-un mod neexclusiv populației celtice din arcul alpin [19] [20] .
Unele studii au identificat existența unei subclade la nivelul arcului alpin situat în nord-vestul Italiei numit S28 / U152 care sugerează existența unui nucleu de dispersie haplotipic tipic peninsulei italiene la nivelul Alpilor, inclusiv în nord -Italia de vest începând cu 1200 î.Hr. [21] , însă prima zonă geografică în care a apărut acest marker nu a fost încă identificată.
Această subclasă a haplogrupului R1b este cea mai comună în peninsula italiană (26,6% în nord și 10,5% în sud) și în insulele Sardinia și Corsica .
E1b1b (fost E3B)
E1b1b (E-M215), cunoscut anterior sub numele de E3B (sau "haplotipul V") [22] este definit de mutația M215. [23] [24] [25] face în prezent obiectul multor controverse.
În prezent, liniile de piese ale E-M215 și E-M35 par identice, se crede că haplogrupul a apărut în Africa de Est acum aproximativ 22400 de ani. [23] [26]
În Europa E-M35 este prezent, unii cercetători au emis ipoteza că această mutație ar putea reprezenta markerul unei migrații antice care a avut loc la sfârșitul Pleistocenului din Africa de Nord în Europa, Sinai și Egipt [27] .
În Italia predomină subclada E1b1b1b1 (E-M81).
Unii cercetători au avansat ipoteza conform căreia haplogrupul E3b1a2 a fost introdus în Marea Britanie începând de la populațiile grecești, împins pe măsură ce erau înrolați de armata romană pentru a coloniza insula britanică, atribuind dominației imperiale romane un rol indirect în difuzare. a acestui haplotip, aparținând Peloponezului, în insula britanică [28] .
J2a-M410 și J2b-M12
Răspândirea haplotipului J2 în bazinul mediteranean este adesea asociată cu expansiunea popoarelor agricole în perioada neolitică [29] . Aspectul J2 este estimat a fi în urmă cu aproximativ 18.500 de ani, cu o diferență de 3.500 de ani în urmă [30] . Unii cercetători îl plasează printre haplogrupurile din Asia de Vest și de Sud-Est, asociindu-l cu prezența descoperirilor arheologice neolitice, precum statuete și ceramică pictată [31], s -a avansat ipoteza că subclada J2a-M410 aparține primei fermieri [32] . Cu toate acestea, alți cercetători fac ipoteza unui posibil eveniment de dispersie în perioada post-neolitică, în special legat de dominația Greciei antice [33] . În Europa, frecvența haplogrupului J2 scade dramatic pe măsură ce ne deplasăm spre nord de la Marea Mediterană.
În Italia, J2 apare cu frecvențe regionale cuprinse între 9% și 36% [34] .
S-a propus că subclada J2a-M410 este legată de populațiile din Creta antică. Haplogrupul J2b-M12 a fost asociat cu Anatolia neolitică (aproximativ 8500 - 4300 î.Hr.) și a fost raportat în siturile cretane (3,1%) [35] .
J1 J-M267
În studiile mai vechi se numește UE10, acest haplogrup se găsește cu frecvențe importante în Orientul Mijlociu, Caucaz, Africa de Nord, Cornul Africii. De asemenea, se găsește mai rar, dar încă ocazional în cantități semnificative, în Europa și Orientul Îndepărtat, cum ar fi subcontinentul indian și Asia Centrală.
J1 este împărțit în mai multe sub-clade, dintre care unele au fost recunoscute chiar înainte de J1, de exemplu J-M62 [36] . Cu excepția J1c3, majoritatea subcladelor nu sunt comune [37] . Frecvența și diversitatea lui J1 (și, de asemenea, a lui J2) fac din acest haplotip unul dintre markerii candidați prin care se presupune că răspândirea tehnologiei agricole în timpul neoliticului poate fi reconstituită.
| Populația | Marime de mostra | J1 total | J1 în absența P58 | J-P58 (J1c3) | publicare |
| Nord-Estul Italiei | 67 | 0,0% | N / A | N / A | Battaglia și colab. (2008) [38] |
| Italieni | 915 | 0,7% | N / A | N / A | Capelli și colab. (2009) [39] |
| Sicilieni | 236 | 3,8% | N / A | N / A | Di Gaetano și colab. (2009) [40] |
G M201
Haplogrupul G al cromozomului Y apare la frecvențe joase în populații multiple, dar este distribuit în Europa Centrală și Asia de Vest, Africa de Nord, India și Sri Lanka.
În prezent, nu există un consens cu privire la datele și locurile de origine ale acestui haplogrup [41] [42] .
În Europa de Vest și zona Mării Negre, haplogrupul G se găsește în medie la aproximativ 5% din populație. Concentrația scade sub această medie în Scandinavia, Polonia, Islanda și Insulele Britanice.
În Tirol (și Tirolul de Sud ) procentul G ajunge la 8% sau mai mult; În zonele de nord și în Sardinia atinge vârfuri la 11% din populație [43] . Analizele genetice asupra rămășițelor mumiei Similaun , mai cunoscute sub numele de Ötzi, au arătat că aparținea unei subclade (G-L91) a acestui haplogrup [44] .
romanizare
Deși fenomenul romanizării, în Europa și în Italia, a adus contribuții importante la nivel cultural, politic și istoric al populațiilor locale, nu există un proces de omogenizare la nivel genetic [45] . Diferitele zone ale Italiei prezintă astăzi diferențe în ceea ce privește distribuția procentuală a haplotipurilor, legate de descendența Y în diferitele zone ale Italiei, care pare mai asemănătoare cu distribuția perioadei preromane.
Din punct de vedere genetic, peninsula italiană și insulele Mediteranei au interesat diverși autori: acest lucru se datorează prezenței simultane a popoarelor care au rămas relativ izolate, cum ar fi sardinii, și a populațiilor enorm eterogene, care au fost întotdeauna afectate de fenomene migratoare, cum ar fi de exemplu cele de coastă din sudul Italiei [45] .
Chiar și migrațiile care au avut loc în solul italic de lacăderea Imperiului Roman de Vest până în anul 1000 d.Hr. nu au modificat în mod semnificativ fondul de gene al italienilor de astăzi; se estimează că haplogrupul I este răspândit în nordul italienilor în ordinea de 2-3% și în sudul italienilor în 1-1,5% [46] . Unele subclade ale haplogrupului R1a, care ar fi putut pătrunde în Italia împreună cu invaziile din nord-vestul Europei, apar în procente cuprinse între 2,5% și în special R1b1c9 de care aparțin 3,5% din italieni [47] , 5,6% în nordul Italiei [12] [13] , deosebit de răspândită în rândul popoarelor din nord-vestul Europei, totuși, întrucât aceste subclade tocmai menționate sunt destul de răspândite și în alte zone europene, este dificil să se stabilească dacă au fost într-adevăr introduse în Italia prin migrațiile acestor popoare.
Arabi
Migrațiile ulterioare ale populațiilor, cum ar fi cele rezultate din raidurile arabe, nu au afectat distribuția haplotipică a Italiei în procente semnificative, cu excepția Siciliei, unde civilizația arabă a înflorit timp de aproximativ două secole și impactul colonizării arabe a avut loc mai intens. peninsula; cu toate acestea, prezența arabă nu a schimbat în mod semnificativ compoziția neolitică a insulei [48] . Contribuția arabă este estimată între 6% și 7,5% în Sicilia, 6,5% în Puglia de nord-vest, 4,8% în estul Campaniei ; trebuie subliniat faptul că studiile anterioare [ citația necesară ] efectuată în aceleași zone a dat procente mai mici. Celelalte regiuni prezintă procente mai mici, de la 0 la 2% [49] [50] .
Arborele filogenetic al haplogrupurilor cromozomului Y și X
Harta genetică a Europei
Mai mulți cercetători au contribuit la dezvoltarea unei hărți genetice a Europei, care arată un grad clar de similaritate structurală cu harta geografică. Principalele diferențe genetice au fost găsite între populațiile din nord și sud. O cercetare genetică publicată în Current Biology , care a eșantionat 24 de populații umane europene, confirmă faptul că diferențele genetice din Europa sunt direct proporționale cu distanța dintre popoarele cu diferențe mai mari pentru Italia și Finlanda. Populațiile care locuiesc în aceste două țări ar prezenta, prin urmare, o distanță genetică mai mare decât celelalte 22 de populații eșantionate din cauza barierelor geografice care le separă de alte populații [51] [52] .
În ceea ce privește Italia în special, un studiu din 2014 a constatat că „populațiile italiene sunt extrem de eterogene din punct de vedere genetic, atât de mult încât diversitatea lor poate fi comparată cu cea observată între grupurile care trăiesc în colțuri opuse ale țării.„ Europa ” [53] [54] . Un alt studiu confirmă ceea ce tocmai a fost raportat asupra eterogenității genetice a populației italiene, care este distribuită de-a lungul unei axe longitudinale vest-est; Sardinienii au în schimb modele specifice [55] [56] , precum și puncte de contact relevante cu primii locuitori istorici ai insulei [57] [58] .
Cercetătorii ipotezează trei evenimente majore de colonizare în Europa, începând din sud. Primii oameni moderni ar fi intrat în ea cu aproximativ 45.000 de ani în urmă, după care s-a produs o întrerupere a fluxurilor migratorii datorită unui maxim glaciar, cu aproximativ 20.000 de ani în urmă; a doua colonizare, care a avut loc când s-a retras gheața, datează de acum aproximativ 17.000 de ani, începând cu populațiile care se întorceau din zonele de refugiu spre sud [59] ; ultima colonizare a avut loc acum aproximativ 10.000 de ani odată cu extinderea agriculturii din Orientul Apropiat . [60] [61]
Notă
- ^ Caracterizarea proceselor biologice care modelează structura genetică a populației italiene , su bmcgenet.biomedcentral.com , BMC Genetics.
- ^ Chiar și astăzi unii biologi, inclusiv Jared Diamond , sunt de părere că distincția dintre genurile Homo și Pan este complet arbitrară și artificială, care poate fi rezolvată prin reclasificarea cimpanzeului comun ca Homo troglodytes și a bonobo ca Homo paniscus .
- ^ Omul modern conform studiilor genetice este originar din Africa. În timpul procesului de migrație numit Out-of-Africa 2 .
- ^ SNPs ( Polimorfisme cu nucleotide unice ).
- ^ Cromozomul Y care nu este recombinat .
- ^ a b Ornella Semino, G. Passarino; PIJAMALE. Oefner et al., The Genetic Legacy of Paleolithic Homo sapiens sapiens in Exant Europeans: AY Chromosome Perspective , în Science Magazine , nr. 5494, noiembrie 2000, pp. 1155-1159.
- ^ [1] Eupedia: Distribuția haplogrupurilor de ADN-cromozom Y european (Y-ADN) după regiune în procente. A se vedea nota nr. 4.
- ^ [2] American Journal of Human Genetics: Mitochondrial Haplogroup U5b3: A Distant Echo of the Epipaleolithic in Italy and the Legacy of the Early Sardinians
- ^ [3] Distribuția europeană a haplogrupurilor ADN mitocondrial (ADNmt) după regiune în procente
- ^ [4] Diferențierea cronologică a haplogrupurilor de ADNmt
- ^ Copie arhivată ( PDF ), la arheologija.ff.uni-lj.si . Adus la 19 martie 2008 (arhivat din original la 6 martie 2009) . Siiri Rootsi: Cromozomul Y haplogrupul I fluxul de gene preistorice în Europa
- ^ a b [5] Myres și colab. - Un haplogrup al cromozomului Y R1b Holocen a fost efectul fondator în Europa Centrală și de Vest (2010)
- ^ a b [6] Dienekes blogspot - Efectul fondatorului R1b în Europa Centrală și de Vest (25 august 2010)
- ^ Capelli C, Brisighelli F, Scarnicci F, Arredi B, Caglia 'A, Vetrugno G, Tofanelli S, Onofri V, Tagliabracci A, Paoli G, Pascali Variația genetică a cromozomului Y în peninsula italiană este clinală și susține un model de amestec pentru Întâlnire mezolitic-neolitic VL. Mol Phylogenet Evol. 2007 iulie; 44 (1): 228-39. Epub 2006 Dec 13. PMID 17275346
- ^ B. Furniture, ES Poloni și C. Tyler-Smith (2007). „Populația Europei”. În Crawford, Michael H .. Genetica antropologică: teorie, metode și aplicații. Cambridge, Marea Britanie: Cambridge University Press. p. 394. ISBN 0-521-54697-4 .
- ^ Chiaroni, J; Underhill, P; Cavalli-Sforza, LL (2009), „Diversitatea cromozomului Y, expansiunea umană, deriva și evoluția culturală”, PNAS 106 (48): 20174: 20179
- ^ a b Myres, Natalie (2010), "A major cromozom haplogroup R1b Holocene effect in Central and Western Europe", European Journal of Human Genetics, doi: 10.1038 / ejhg.2010.146
- ^ Cruciani și colab. (2010), "Diferențierea puternică intra și inter-continentală dezvăluită de SNPs cromozomului Y M269, U106 și U152", Forensic Science International: Genetics, doi: 10.1016 / j.fsigen.2010.07.006
- ^ David K. Faux Un semnal genetic al ascendenței celtice din Europa Centrală: cercetări preliminare privind marcatorul cromozomului Y U152 17 noiembrie 2008 [7]
- ^ David K. Faux Y-Chromosome Marker S28 / U152 Haplogroup R-U152
- ^ Harta dispersiei subcladei S28 / U152 [8] bazată pe reconstrucția liniei Y ISOGG [9]
- ^ Gérard și colab.; Berriche, S; Aouizérate, A; Diéterlen, F; Lucotte, G (2006), „Berberii nord-africani și influențele arabe în vestul Mediteranei dezvăluite de haplotipurile ADN-ului cromozomului Y”, Human Biology 78 (3): 307-316, doi: 10.1353 / hub. 2006.0045, PMID 17216803
- ^ a b Cruciani și colab., F.; La Fratta, R.; Trompetă, B.; Santolamazza, P.; Sellitto, D.; Colomb, EB; Dugoujon, J.-M.; Crivellaro, F. și colab. (2007), „Tracing Movements Human Human Movements in Northern / Eastern Africa and Western Eurasia: New Clues from Y-Chromosomal Haplogroups E-M78 and J-M12”, Molecular Biology and Evolution 24 (6): 1300–1311, doi: 10.1093 / molbev / msm049, PMID 17351267 A se vedea, de asemenea, Date suplimentare.
- ^ Adams și colab.; Bosch, Elena; Balaresque, Patricia L.; Ballereau, StéPhane J; Lee, Andrew C.; Arroyo, Eduardo; López-Parra, Ana M.; Aler, Mercedes și colab. (2008), „Moștenirea genetică a diversității religioase și a intoleranței: descendențe paterne ale creștinilor, evreilor și musulmanilor din Peninsula Iberică”, The American Journal of Human Genetics 83 (6): 725, doi: 10.1016 / j.ajhg. 2008.11.007, PMC 2668061, PMID 19061982
- ^ Silva și colab.; Carvalho, Elizeu; Costa, Guilherme; Tavares, Lygia; Amorim, António; Gusmão, Leonor (2006), "Variația genetică a cromozomului Y în populația Rio De Janeiro", American Journal of Human Biology 18 (6): 829-837 doi = 10.1002 / ajhb.20567, doi: 10.1002 / ajhb.20567, PMID 17039481
- ^ Cruciani și colab., Fulvio; La Fratta, Roberta; Santolamazza, Piero; Sellitto, Daniele; Pascone, Roberto; Moral, Pedro; Watson, Elizabeth; Ghid, Valentina și colab. (Mai 2004), „Analiza filogeografică a cromozomilor Haplogroup E3b (E-M215) Y dezvăluie evenimente migratorii multiple în și din Africa” (PDF), American Journal of Human Genetics 74 (5): 1014-1022, doi: 10.1086 / 386294, PMC 1181964, PMID 15042509
- ^ Underhill și Kivisild; Kivisild, Toomas (2007), "Utilizarea cromozomului Y și a structurii populației ADN-ului mitocondrial în urmărirea migrațiilor umane", Annu. Pr. Genet. 41: 539–64, doi: 10.1146 / annurev.genet.41.110306.130407, PMID 18076332
- ^ Steven C. Bird Haplogroup E3b1a2 ca posibil indicator al așezării în Marea Britanie de către soldații de origine balcanică http://www.jogg.info/32/bird.htm
- ^ ISOGG , pe isogg.org .
- ^ Ornella Semino și colab., "Originea, difuzia și diferențierea haplogrupurilor E și J ale cromozomului Y: inferențe asupra neolitizării Europei și evenimente migratorii ulterioare în zona mediteraneană", American Journal of Human Genetics 74: 1023-1034, 2004
- ^ R. King și PA Underhill (2002), Distribuție congruentă a ceramicii pictate în neolitic și figurine ceramice cu descendențe ale cromozomului Y, Antichitate 76: 704-714
- ^ J. Chiaroni și colab. (2008), Corelația precipitațiilor anuale cu diversitatea cromozomului Y uman și apariția economiilor agricole și pastorale neolitice în Semiluna Fertilă, Antichitate Volum: 82 Număr: 316 Pagină: 281–289
- ^ F. Di Giacomo și colab. (2004), haplogrupul cromozomial Y ca semnătură a colonizării postneolitice a Europei, Human Genetics 115 (5): 357-71
- ^ Hair C et. Variația genetică a cromozomului Y în peninsula italiană este clinală și susține un model de amestec pentru întâlnirea mezolitic-neolitic Mol Phylogenet Evol. 2007 iulie; 44 (1): 228-39. Epub 2006 13 decembrie
- ^ Regele, RJ; Ozcan, SS, Carter, T., Kalfoglu, E., Atasoy, S., Triantaphyllidis, C., Kouvatsi, A., Lin, AA, Chow, CE. T., Zhivotovsky, LA, Michalodimitrakis, M., Underhill, PA, (2008). „Influențele anatoliene ale cromozomului Y diferențial asupra neoliticului grec și cretan”. Annals of Human Genetics 72 Numărul 2 martie 2008 (Pt 2): 205-214. doi: 10.1111 / j.1469-1809.2007.00414.x. PMID 18269686
- ^ Y Chromosome Consortium "YCC" (2002), "A Nomenclature System for the Tree of Human Y-Chromosomal Binary Haplogroups", Genome Research 12 (2): 339–348, doi:10.1101/gr.217602, PMC 155271, PMID 11827954
- ^ Tofanelli, Sergio; Ferri, Gianmarco; Bulayeva, Kazima; Caciagli, Laura; Onofri, Valerio; Taglioli, Luca; Bulayev, Oleg; Boschi, Ilaria et al. (2009), "J1-M267 Y lineage marks climate-driven pre-historical human displacements", European Journal of Human Genetics 17: 1520–1524, doi:10.1038/ejhg.2009.58
- ^ Battaglia; Fornarino, Simona; Al-Zahery, Nadia; Olivieri, Anna; Pala, Maria; Myres, Natalie M; King, Roy J; Rootsi, Siiri et al. (2008), "Y-chromosomal evidence of the cultural diffusion of agriculture in southeast Europe", European Journal of Human Genetics 17 (6): 820–830, doi:10.1038/ejhg.2008.249, PMC 2947100, PMID 19107149
- ^ Capelli; Onofri, Valerio; Brisighelli, Francesca; Boschi, Ilaria; Scarnicci, Francesca; Masullo, Mara; Ferri, Gianmarco; Tofanelli, Sergio et al. (2009), "Moors and Saracens in Europe: estimating the medieval North African male legacy in southern Europe", European Journal of Human Genetics 17 (6): 848–852, doi:10.1038/ejhg.2008.258, PMC 2947089, PMID 19156170
- ^ Di Gaetano; Cerutti; Crobu; Robino (2009), " Differential Greek and northern African migrations to Sicily are supported by genetic evidence from the Y chromosome ", European Journal of Human Genetics 17 (1): 91–99, doi:10.1038/ejhg.2008.120, PMC 2985948, PMID 18685561
- ^ Semino O, Passarino G, Oefner PJ, et al. (November 2000). "The genetic legacy of Paleolithic Homo sapiens sapiens in extant Europeans: a Y chromosome perspective". Science 290 (5494): 1155–9. doi:10.1126/science.290.5494.1155. PMID 11073453
- ^ Cinnioğlu C, King R, Kivisild T, et al. (January 2004). "Excavating Y-chromosome haplotype strata in Anatolia". Hum. Genet. 114 (2): 127–48. doi:10.1007/s00439-003-1031-4. PMID 14586639
- ^ Zei, G. et al. (2003). "From surnames to the history of Y chromosomes: the Sardinian population as a paradign.". Eur J of Human Genetics 11 (10): 802–07. doi:10.1038/sj.ejhg.5201040. PMID 14512971
- ^ Andreas Keller, Angela Graefen, Markus Ball, Mark Matzas, Valesca Boisguerin, Frank Maixner, Petra Leidinger, Christina Backes, Rabab Khairat, Michael Forster, Björn Stade, Andre Franke, Jens Mayer, Jessica Spangler, Stephen McLaughlin, Minita Shah, Clarence Lee, Timothy T. Harkins, Alexander Sartori, Andres Moreno-Estrada, Brenna Henn, Martin Sikora, Ornella Semino, Jacques Chiaroni, Siiri Rootsi, Natalie M. Myres, Vicente M. Cabrera, Peter A. Underhill, Carlos D. Bustamante e Eduard Egarter Vigl, New insights into the Tyrolean Iceman's origin and phenotype as inferred by whole-genome sequencing , in Nature Communications , vol. 3, 28 febbraio 2012, p. 698, DOI : 10.1038/ncomms1701 , PMID 22426219 .
- ^ a b Cavalli Sforza Luigi L.; Menozzi Paolo; Piazza Alberto, Storia e geografia dei geni umani , Adelphi (collana Gli Adelphi), 2000(2 ed.), EAN 9788845915888.
- ^ [10] Archiviato il 24 giugno 2009 in Internet Archive . Phylogeography of Y-Chromosome Haplogroup I Reveals Distinct Domains of Prehistoric Gene Flow in Europe
- ^ [11] Y-chromosome Short Tandem Repeat DYS458.2 Non-consensus Alleles Occur Independently in Both Binary Haplogroups J1-M267 and R1b3-M405
- ^ Differential Greek and northern African migrations to Sicily are supported by genetic evidence from the Y chromosome , su ncbi.nlm.nih.gov , 2008.
- ^ European Journal of Human Genetics , https://www.nature.com/articles/ejhg2008258 .
- ^ Dienekes blogspot : North African male legacy in Southern Europe quantified , su dienekes.blogspot.com .
- ^ ( EN ) Nicholas Wade, Visual Science - The Genetic Map of Europe , in NY Times , The New York Times Co., 13 agosto 2008.
- ^ ( EN ) AA.VV., Correlation between Genetic and Geographic Structure in Europe , in Current Biology , Cell Press, 26 agosto 2008, DOI : 10.1016/j.cub.2008.07.049 .
- ^ Biodiversità umana, italiani al primo posto
- ^ Capocasa M. et al. Linguistic, geographic and genetic isolation: a collaborative study of Italian populations. Journal of Anthropological Sciences, volume 92, doi 10.4436/JASS.92001
- ^ Gli italiani non esistono. Siamo un grande mix genetico. Tranne i sardi , in Il Corriere della Sera , 2018.
- ^ Accademia delle Scienze di Torino , Alberto Piazza, I profili genetici degli italiani ( PDF ), su units.it .
- ^ Il Dna dei sardi è il più simile a quello degli europei preistorici , in Il Corriere della Sera , 2017.
- ^ Seimila anni di storia dal DNA degli antichi sardi , in La Repubblica , 2020.
- ^ Con il termine zone di rifugio, si intendono regioni geografiche non meglio identificate nelle quali i primi migranti giunti in europa si sarebbero concentrati in seguito all'avanzamento dei ghiacci, per poi fuoriuscirne ed espandersi al ritiro degli stessi, cioè al termine dell'ultimo massimo glaciale
- ^ Genes mirror geography within Europe , su ncbi.nlm.nih.gov .
- ^ Genes mirror geography within Europe - Figure 2
Bibliografia
- Luigi L. Cavalli Sforza, Paolo Menozzi e Alberto Piazza, Storia e geografia dei geni umani , 2ª ed., Milano, Adelphi, 2000, ISBN 978-88-459-1588-8 .
- L. e F. Cavalli-Sforza , Chi siamo , Milano, Mondadori, 1993, ISBN 88-04-36901-9 .
- Luigi L. Cavalli Sforza e Albert J. Ammerman, La transizione neolitica e la genetica di popolazione in Europa , Torino, Boringhieri, 1986, ISBN 88-339-0043-6 .
- Luigi L. Cavalli Sforza, Francesco Cavalli-Sforza e Ada Piazza, Razza o pregiudizio? L'evoluzione dell'uomo fra natura e storia , Milano, Einaudi scuola, 1996, ISBN 88-286-0297-X .
- ( EN ) Cultural transmission and evolution. A quantitative approach , con Marcus W. Feldman, Princeton, Princeton University Press, 1981.
- ( EN ) Luigi L. Cavalli-Sforza e AWF Edwards, Analysis of human evolution , in SJ Geerts (a cura di), Genetics Today. Proceedings of the XI International Congress of Genetics , The Hague , The Netherlands, September, 1963 , vol. 3, Oxford, Pergamon Press, 1965, pp. 923–933.
- ( EN ) The genetics of human population , con Walter F. Bodmer, San Francisco, WH Freeman and C., 1971.
Voci correlate
- Ipotesi genetiche sul popolamento dell'Europa
- Storia genetica dell'Europa
- Storia genetica della Sardegna
Altri progetti
-
 Wikimedia Commons contiene immagini o altri file su Storia genetica dell'Italia
Wikimedia Commons contiene immagini o altri file su Storia genetica dell'Italia

![[8]](http://www.eupedia.com/images/content/Haplogroup-R1b-S28.gif){kind=link}