Aphidoidea
| Afide | |
|---|---|
 | |
| Clasificare științifică | |
| Domeniu | Eukaryota |
| Regatul | Animalia |
| Sub-regat | Eumetazoa |
| Superphylum | Protostomie |
| Phylum | Arthropoda |
| Subfilum | Hexapoda |
| Clasă | Insecta |
| Subclasă | Pterygota |
| Cohortă | Exopterygota |
| Subcoorte | Neoptere |
| Superordine | Paraneoptere |
| Secțiune | Rhynchotoidea |
| Ordin | Rhynchota |
| Subordine | Homoptera |
| Secțiune | Sternorrhyncha |
| Superfamilie | Aphidoidea Geoffroy , 1762 |
| Denumiri comune | |
Afide | |
| Familii | |
Afidele sau păduchii de plante ( Aphidoidea Geoffroy , 1762 ) sunt o superfamilie de insecte fitomitice incluse în ordinea Rhynchota ( subordine Homoptera , secțiunea Sternorrhyncha ).
Morfologie
Afidele au un corp îndesat și mic, cu capul și toracele în general mai mici decât abdomenul; profilul își asumă o formă globulară și conică în față. Exoscheletul este moale și delicat; afidele moarte, de fapt, își păstrează cu greu forma corpului. Ele prezintă adesea un polimorfism intraspecific marcat care se încadrează în general în cazurile de peciloginie (polimorfism în sexul feminin), cu indivizi înaripați și înaripați. Mai rar, polimorfismul afectează și sexul masculin.
Capul este prevăzut cu trei ocelli ; ochii compuși sunt prevăzuți cu un proces lateral care transportă trei omatidii (numiți ochi juvenili ). Aparatul bucal este de tip aspirator înțepător , cu rostru , a cărui bază, ca în toate Sternorrinchi, este situată între coxurile picioarelor din față. Morfologia antenelor este un element de determinare sistematică. În general, acestea sunt subțiri și alungite, îndreptate în sus și înapoi, uneori atât cât întregul corp. Acestea sunt formate din 3-6 articole, dintre care cele două proximale sunt destul de scurte, celelalte, formând flagelul , sunt vizibil alungite. În general, formele aferente au trei articole și șase aripi. Antenomul terminal are cea mai mare dezvoltare și este compus dintr-o parte bazală, de același diametru ca și ceilalți antenomeri și o extensie terminală subțire și alungită.

În scopuri taxonomice, o importanță deosebită este rezervată prezenței și numărului de sensili prezenți pe antene: la vârful părții bazale a ultimului antennomer, sau înainte de extensia terminală, există un sensilus placoid numit rhinario primar , de formă circulară , uneori împodobit cu fire de păr; un al doilea rinariu primar este prezent la vârful penultimului antennomer (de obicei al cincilea). Al doilea rinariu este absent în schimb în antenele compuse din doar trei articole. În formele înaripate există și alți sensilli, de dimensiuni mai mici și număr și formă variabile (circulare, ovale sau transversale); acestea sunt numite rhinari secundari și sunt întotdeauna situate în ultimul segment, lângă rhinario primar, și adesea și în al patrulea și al cincilea segment. Formele apterice sunt în general lipsite de rinari secundari sau sunt prevăzute cu ele pe al treilea segment.
Toracele este redus și nu foarte diferențiat în formele apterice; segmentele sale își asumă un aspect substanțial similar cu cel al segmentelor abdominale, din care se disting prin dimensiunea mai mică și prin prezența picioarelor. În formele înaripate, protoraxul ia forma unui guler distinct de cap, în timp ce mezo și metatorax tind să se unească într-o singură regiune distinctă de protorax și abdomen. Picioarele sunt de tip ambulator, relativ lungi și subțiri, cu picioarele formate din două segmente.

C : costuri; Pt : pterostigma; R s : sector radial; M : ramuri ale mass-media; Cu : ramuri ale cotului.
Aripile , atunci când sunt prezente, sunt toate membranate și au o venulație simplificată, uniformă în generalitatea afidelor. În aripa anterioară există o singură coastă longitudinală mare, interpretată ca fuziunea subcostei, a razei, a mediei și a cubitului. Această coastă se desfășoară paralel cu marginea costală și se termină distal într-un pterostigma mai mult sau mai puțin alungit. În spate, din această nervură, ramurile unice se ramifică, interpretate după cum urmează începând de la ramura terminală până la cea proximală: sector radial (R s ), mediu (M), prima ramură a cotului (Cu 1 ) și a doua ramura cotului (Cu 2 ). Sectorul radial este scurt și începe de la pterostigma. În schimb, media suferă două bifurcații: de la prima bifurcație are loc o ramură care la rândul ei se bifurcă în R 1 și R 2 , în timp ce cea posterioară rămâne nedivizată și, prin urmare, interpretată ca o fuziune a ramurilor R 3 și R 4 . Pterostigma este o areolă sclerificată, constând din fuziunea subcostei și a ramurii de rază (R 1 ). Aripa posterioară reia venulația celei anterioare dar substanțial mai simplificată, cu absența pterostigmei și prezența a doar trei ramuri ale coastei principale (sectorul radial, mijlocul și cotul).
Poziția aripilor în faza de repaus este caracteristică: de fapt, acestea sunt pliate înapoi și ținute vertical și împerechere, ridicate deasupra abdomenului. În aparență, se pare că afidele își pliază aripile ca fluturii diurni, în realitate marginea costală este poziționată în partea de jos. În esență, este, prin urmare, o pliere care poate fi urmărită înapoi la cea a majorității homopterelor, în care aripile sunt în general pliate peste abdomen.
Abdomenul este ghemuit, format din 9 urite. Ultima urită are un proces posterior, numit codicul , utilizat pentru îndepărtarea mierii . Pe partea dorsal-laterală a abdomenului, între a cincea și a șasea urită, există două tuburi mai mult sau mai puțin alungite, numite sifoane sau cornicule . În unele afide sifoanele sunt absente sau reduse la găuri simple. Sifoanele sunt folosite ca mijloc de apărare: în situații de pericol secretă o ceară fluidă care se solidifică rapid, încâlcind gura sau picioarele oricărui prădător . Sifoanele emit, de asemenea, un feromon care acționează ca un semnal de alarmă chimică.
Ovipozitorul este în general absent.
Anatomie și fiziologie
Cel mai important aspect anatomo-fiziologic este absența trompelor Malpighiene , prin urmare Afidele, cu puține excepții, nu au propriul sistem excretor care să prevadă eliminarea cataboliților de amoniac derivați din metabolismul proteinelor. Funcția de detoxifiere este realizată de bacteriile simbiotice adăpostite într-un micetom .
Nutriția are loc în detrimentul sucurilor celulelor parenchimatoase individuale sau în detrimentul limfei procesate. În acest caz, stiletele sunt capabile să străpungă țesuturile plantei pătrunzând adânc până la vasele floemului și să injecteze saliva necesară pentru a face limfa mai fluidă. Aceasta trece de-a lungul canalului alimentar printr-o simplă ascensiune capilară , prin urmare afidul nu trebuie să exercite nicio forță de aspirație. Acest aspect este uneori exploatat de cercetători pentru a preleva probe de sevă procesate folosind afide decapitate.
Genetică și reproducere

Partenogeneza apomictică este deosebit de răspândită în Afide (stabilitatea caracterelor din descendenți datorită absenței meiozei ). Reproducerea amfigonică alternează, în general, după un număr nespecificat de generații partenogenetice. Amfigonia nu explică singura variabilitatea genetică considerabilă a afidelor. Pe baza observațiilor cariologice, COGNETTI ( 1961 ), a descris o formă singulară de recombinare a genelor , numită endomeioză , care ar explica variabilitatea genetică a acestor insecte: înainte de segmentarea nucleului oului partenogenetic, împerecherea cromozomilor omologi cu apariția de trecere peste fenomene [1] . Deși mecanismele nu sunt pe deplin clare, alte experiențe au evidențiat existența mecanismelor de variabilitate genetică în cadrul unei partenogeneze apomictice [2] .
Determinarea sexului are loc în prima diviziune a oului partenogenetic. Setul cromozomial al femelelor include doi cromozomi sexuali, cel al masculului doar unul. Pierderea unui cromozom sexual în timpul primei diviziuni are ca rezultat dezvoltarea unui bărbat partenogenetic. Sperma conține întotdeauna cromozomul sexual, de aceea ouăle obținute din reproducerea amfigonică sunt întotdeauna femele.
Un alt aspect particular al potențialului reproductiv al multor specii de afide este viviparitatea . Acest comportament se găsește în familia Aphididae , niciodată în cele din Adelgidae și Phylloxeridae (familii incluse în mod tradițional în grupul Aphidoidea ovipara ). La speciile vivipare, femelele partenogenetice sunt numite virginopare și embrionii a trei generații cuibărite se dezvoltă în interiorul lor. Acest comportament necesită o aprovizionare considerabilă cu energie, garantată de capacitatea excepțională de a se hrăni cu aceste insecte și explică numărul mare de generații care caracterizează adesea ciclul afidelor.
Dezvoltare post-embrionară și polimorfism

Dezvoltarea post-embrionară trece prin patru etape, cu diferențe marcate între indivizii înaripați și fără aripi: în cele două etape succesive aripate ale neanidului , unul al preninfei și al unei nimfe ; în acele ateros se trece prin patru etape ale neanidei. Diferențele morfologice dintre stadiile juvenile și cele adulte, la indivizii fără aripi, privesc numărul de articole ale antenelor și forma codiculului și a sifonelor.
Polimorfismul este o trăsătură recurentă la afide și este asociat cu posibila complexitate a ciclului lor. Tipul normal, prevăzut cu aripi, este flancat de forme apterice care trebuie interpretate ca adaptare la o fază specifică a ciclului. Formele apterice derivă din prezența unei cantități excesive de neotenină , hormonul juvenil; lipsei dezvoltării aripilor sunt asociate alte caractere secundare care se referă în cea mai mare parte la dezvoltarea ochilor compuși, a antenelor și a prezenței ocelilor.
În afară de gradele de diferențiere polimorfă, uneori duse la extrem la unele specii, unele tipuri polimorfe fundamentale se pot distinge prin rigoare schematică. Trebuie remarcat faptul că între aceste tipuri pot fi inserate polimorfisme intermediare care complică și mai mult, în sens morfologic și etologic, caracterizarea anumitor specii.
- Fondatori . Femelele aeriene au ieșit dintr-un ou amfigonic. Se reproduc prin partenogeneză.
- Virginopare . Femele pterale sau înaripate, născute prin partenogeneză. Cei înaripați, numiți și migranți, sunt responsabili pentru propagarea speciei pe diferite gazde. Indiferent de gradul de meiotterism , toate virginoparele se reproduc prin partenogeneză. Terminologiile specifice sunt utilizate pentru a diferenția funcțional virginoparii născuți pe gazda primară și pe același aparat ca și fondatoarea ( fondatrigenie ) sau pe gazda secundară sau pe aparate diferite de cea a fondatorului ( virginogenii sau exilați).
- Sex . Femele pterale sau înaripate, născute prin partenogeneză. Diferențierea morfologică și etologică a sesupării este extrem de variabilă, în funcție de specie, astfel încât în această categorie există diferite tipuri:
- Gynoparous : femele cu aripi, se reproduc prin partenogeneză generând femele amfigonice anfigonice.
- Andropare : femele atere , se reproduc prin partenogeneză generând masculi înaripați (amfigoni).
- Amfipare : femelele apterice sau înaripate se reproduc prin partenogeneză generând atât bărbați, cât și femele, atât amfigonice, cât și fără aripi.
- Femele amfigonice . Femele ateroase, născute prin partenogeneză. Se reproduc prin amfigonie prin împerecherea cu masculii.
- Bărbați. Indivizi cu aripi sau fără aripi, născuți prin partenogeneză.
După cum sa menționat anterior, pot apărea forme intermediare între tipurile descrise care, pe lângă prezentarea detaliilor morfologice și funcționale care le identifică, au o etologie derivată mai ales dintr-o adaptare la gazde specifice sau condiții de mediu în general. Trebuie remarcată și apariția polimorfismelor intermediare generate de relația trofică cu unele endoparazitoide .
Sinteza carotenoidelor
Carotenoizii sunt fundamentali în multe procese biologice; membrii regnului animal , însă, nu îi produc și îi iau cu hrana lor, precum și vitamine și aminoacizi, prin urmare trebuie să fi existat, la nivel ancestral, o pierdere a genelor sintetizatoare. Cu toate acestea, s-a descoperit că unele specii de afide au dobândit gene de origine fungică prin transfer orizontal de gene , datorită cărora sunt capabile să producă caroten prin procesul de fotosinteză anaerobă. [3]
Aceste insecte sunt recunoscute prin culoarea lor roșie în loc de verde; afidele sunt singurii reprezentanți ai regnului animal capabili să sintetizeze carotenoizi. [4]
Ciclu
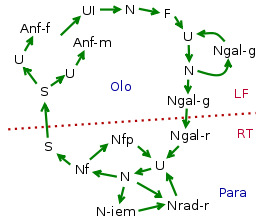 |
| Filoxera viței de vie ( Daktulosphaira vitifoliae ) este un exemplu de ciclu complex, monoic și heterotopic cu coexistența unui holociclu și a unui paracic și o tendință spre anolociclu în Europa . Legendă
|
Datorită complexității sale, ciclul biologic este un aspect interesant care evidențiază gradul considerabil de specializare a afidelor și adaptarea lor la anumite condiții de mediu (gazde ale plantelor) și condiții climatice (curs sezonier). Rezultatul este un potențial biologic ridicat, care exploatează pe de o parte amfigonia ca mijloc de reproducere și pe de altă parte partenogeneza ca mijloc de multiplicare: cu amfigonia, specia se perpetuează de la an la an, depășind rigorile iernii., Cu partenogeneză populația se înmulțește rapid colonizând mediul. Multiplicitatea stimulilor de mediu este responsabilă pentru diferențierea ciclurilor în diferitele specii de afide.
În general, ciclurile de afide văd succesiunea alternativă între o generație amfigonică și un număr nedeterminat de generații partenogenetice. Pe baza frecvenței generației amfigonice, se disting trei tipuri diferite de cicluri:
- holociclu : în fiecare an se formează o generație amfigonică din care provine forma de iernare;
- paraciclo : generația amfigonică apare cu o frecvență de mai mulți ani, în orice caz prevalența generațiilor partenogenetice este clară;
- anolociclu : generația amfigonică dispare complet și specia se perpetuează numai prin partenogeneză.
În raport cu gazda plantei, se disting două tipuri de cicluri:
- heteroic : ciclul are loc pe diferite tipuri de gazde distincte, respectiv, în primar și secundar. Foarte des gazda primară este reprezentată de o specie arbore sau arbustivă , cea secundară de o plantă erbacee . În general, există o specificitate a relației cu gazda primară și o polifagie în relația cu oaspeții secundari;
- monoice : ciclul are loc pe o singură specie de plantă.
În cele din urmă, în raport cu organele afectate, se disting două tipuri diferite de cicluri:
- heterotop : ciclul se dezvoltă pe diferite părți ale plantei;
- homotop : ciclul se dezvoltă pe aceeași parte a plantei.
Din cele spuse, este clar că diferitele specii de afide pot prezenta cicluri complet diferite ca o adaptare la condiții climatice și de mediu specifice.
Ciclul heteroic și homotop
Este cel mai complex tip, în general sub forma unui anolociclu, care denotă o adaptare a speciei la un curs sezonier caracterizat prin alternanța dintre un sezon cald și unul rece, tipic regiunilor temperate. Ciclul vede alternanța dintre câteva generații de primăvară, asociate cu o specie de plante arborice și un număr nedeterminat de generații de vară asociate cu o plantă erbacee anuală. Pe măsură ce toamna se apropie, specia revine la gazda sa principală pentru iernare. Etapa de iernare este reprezentată de ou, numit ou durabil , depus în general între interstițiile scoarței gazdei primare.

Un ciclu heteroic și homotop tipic se dezvoltă în conformitate cu următoarea schemă. Din ouul durabil, obținut prin amfigonie, se naște o femelă, numită întemeietoare . Două sau trei generații de păsări virgine derivă de la fondator, numit fondatrigenie , întotdeauna asociat cu gazda primară. Fondatrigeniile sunt fundamental abere, totuși în aceste generații apar și indivizi înaripați, cu un procent crescător care trece de la a doua la a treia. Aripile sunt păsări virgine migratoare și se mută la speciile de plante care acționează ca gazde secundare.
Gazda secundară este urmată de un număr nedeterminat de generații de virginogenii, numite exilați , din care se nasc și alte virginopare, atât atere, cât și înaripate. Aceștia din urmă sunt responsabili pentru propagarea pe alte gazde secundare aparținând sau nu aceleiași specii ca gazda din care provin.
Aproape de toamnă, apar sesupare, generația partenogenetică de tranziție din care va proveni amfigonicul. În general, sesupara este de două tipuri: ginoparii sunt înaripați și migrează către gazda primară, pe care se reproduc generând femelele amfigonice; androparele sunt pe de altă parte anterioase și generează masculi înaripați pe gazda secundară care la rândul lor migrează la gazda primară pentru reproducere. Pe de altă parte, la multe specii sessupare sunt nediferențiate din punct de vedere etologic și sunt toate înaripate. În acest caz se comportă ca amfipare și după ce au migrat la gazda primară se reproduc indiferent generând amfigoni fără aripi de ambele sexe. În cele din urmă, la alte specii, apar forme intermediare între sesupare și virginogenii, din care se pot naște alte femele partenogenetice sau indivizi amfigonici, complicând și mai mult scanarea ciclului.
În cele din urmă, această aparentă complicație comportamentală a sesupării are un scop biologic fundamental derivat dintr-o nevoie de adaptare: gazda secundară este, în general, o plantă erbacee care va fi deshidratată la sfârșitul ciclului său; mecanismele de migrație diferențială au scopul de a face ca reproducerea amfigonică să aibă loc pe gazda primară, singura capabilă să permită iernarea și supraviețuirea speciei. Indiferent de modul în care sunt generate, prin urmare, amfigonienii se împerechează pe gazda primară, iar femelele depun oul durabil în puncte capabile să-l protejeze de adversitățile climatice și de prădători.
Ciclul monoic și homotop

Schematic, este mai simplu decât precedentul tocmai datorită faptului că nu există un invitat secundar. Planta gazdă este reprezentată de o specie de plantă lemnoasă, capabilă să garanteze, pe de o parte, supraviețuirea formelor de iarnă și dezvoltarea generațiilor de primăvară și, pe de altă parte, multiplicarea speciilor afidice. Aparent, schema retrage etapele celei anterioare cu o simplificare terminologică: în acest caz, de exemplu, distincția dintre fondatrigenie și exilați în lumea virginopară nu are sens, deoarece toate generațiile partenogenetice se dezvoltă pe gazde ale aceleiași specii.
Virginogeniile înaripate au sarcina de a propaga linii paralele cu cele ale fondatorului pe oaspeții aceleiași specii. Pe măsură ce se apropie toamna, apar sesupare, care în acest tip de ciclu sunt practic amfipare și aptere.
Paracicluri și anolocicluri
Cele descrise mai sus pot fi considerate două cazuri extreme între care pot apărea forme intermediare de cicluri care denotă versatilitatea remarcabilă a liniilor evolutive din această superfamilie. Ca o consecință a factorilor legați de specie, cum ar fi adaptarea evolutivă, sau la anumite condiții de mediu și climatice, cum ar fi adaptarea contextuală, pot fi găsite divergențe semnificative față de cele două scheme anterioare, cum ar fi efectuarea unui ciclu monoic pe gazdă primară sau secundară paralelă cu un ciclu heteroic sau dezvoltarea paraciclurilor pe oaspeții secundari intercalate, cu intervale de mai mulți ani, la reproduceri amfigonice ocazionale pe oaspeții primari, până la cazul extrem în care reproducerea amfigonică dispare definitiv odată cu efectuarea anolocicluri pe oaspeții primari sau secundari.
În multe cazuri, aceste diferențieri sunt rezultatul unei adaptări la condițiile climatice și de mediu favorabile care transformă alternanța forțată a anolociclilor în alternanțe opționale. Aceste condiții apar adesea în regiunile climatice cu ierni blânde, unde dispare necesitatea de a garanta iernarea, sub forma unui ou durabil.
Determinismul polimorfismului

Polimorfismul afidelor este determinat de interacțiunea complexă dintre factorii endogeni, climatici și de mediu. Această interacțiune se manifestă în special prin apariția virginogeniilor sesupare și aripioase. De fapt, în condiții de mediu favorabile, potențialul biologic al afidelor este realizat în tot potențialul său cu bacterialism și reproducere partenogenetică.
Apariția sesupare este, în general, un răspuns la stimulii de mediu de natură climatică, deoarece această fază este fundamentală pentru iernarea speciei în regiunile reci. Stimulii de mediu sunt reprezentați de scăderea fotoperiodei și scăderea temperaturii în regiunile reci și doar scăderea temperaturii în regiunile blânde de iarnă [5] . Din acest motiv, în nordul Italiei, sessupare apar de obicei la sfârșitul verii, în timp ce în sudul Italiei apar frecvent la sfârșitul toamnei. Comportamentul speciilor de rădăcină este indus în schimb de modificări ale fiziologiei sistemului radicular al gazdei [5] . Factorii endogeni, verificați în condiții experimentale, interferează cu stimulii derivați din climă sau din fiziologia plantei gazdă, care au scopul biologic de a reduce frecvența generației amfigonice: sesuparele se formează de fapt mai ușor cu cât sunt partenogenetice. generația este îndepărtată de cea a fondatorului. Cazul extrem este cel al fondatorilor, care nu pot genera sesupare chiar dacă sunt reproduse condițiile climatice-de mediu menționate mai sus. Acest comportament se regăsește, cu intensitate mai mică, și în 3-4 generații succesive de virginogenii [5] .

Apariția virginogeniilor înaripate trebuie considerată un eveniment necesar pentru propagarea speciei către condiții de mediu mai favorabile: potențialul reproductiv este, de fapt, redus în cele înaripate, care sunt în mod fundamental mai puțin fertile decât apotecarii. Apariția este deci determinată mai ales de factori nutriționali și sociali [5] : colonii de virginogenii apterice se dezvoltă pe lăstarii sensibili și apariția celor înaripate apare doar în condiții de supraaglomerare; la cei aflați în procesul de lignificare, aspectul celor înaripate ar fi în schimb determinat de conținutul diferit de aminoacizi din limfa procesată, prin urmare comportamentul răspunde la un stimul nutrițional.
În cele din urmă, un caz particular se găsește în relațiile dintre furnici și afide [5] . Coloniile protejate de furnici sunt de fapt formate aproape exclusiv de virginogenii apterice: furnicile beneficiază de creșterea virginogeniilor apterice, deoarece acestea se hrănesc cu o voracitate mai mare și, prin urmare, produc cantități mai mari de mieră . În acest caz, inhibarea apariției aripilor este puțin cunoscută și este probabil atribuibilă mecanismelor fiziologice care induc persistența hormonului juvenil.
Afidele și agricultura

Afidele se numără printre cele mai cunoscute insecte fitofage din cauza pagubelor extinse cauzate culturilor. Potențialul lor reproductiv este atât de mare încât populația lor atinge rapid niveluri care determină decăderea gazdei atacate. Asocierea recombinării genelor pentru endomeioză cu partenogeneză este, de asemenea, un factor biologic de succes care îngreunează controlul chimic: insecticidele sistemice sunt de fapt cele mai eficiente în lupta împotriva afidelor, cu toate acestea specificitatea lor structurală este cauza mutațiilor care generează rezistență., Care , datorită partenogenezei, se răspândește rapid în populație. La fel ca acarienii și ciupercile patogene, afidele sunt, prin urmare, printre adversitățile de natură biologică care prezintă frecvent rezistență la agenții chimici.
Daunele directe cauzate de afide sunt următoarele:
- Scăderea limfei. Aceasta este dauna directă care fură resursele de energie din centrală.
- Modificarea fiziologiei. Saliva injectată de stiletele maxilare interferează cu fiziologia plantei, inducând o activitate respiratorie mai mare și, prin urmare, o cheltuială mai mare de resurse energetice.
- Alterarea structurii morfoanoatomice. Saliva multor specii conține substanțe asemănătoare auxinelor care induc fenomene de alterare a dezvoltării cu formarea de galele , pseudogale și deformări (rulare, curlare, curbare a frunzelor). Aceste modificări implică, în general, o reducere a indicelui zonei frunzelor și, prin urmare, o reducere a eficienței asimilative a plantelor.

Pagubele directe au repercusiuni economice imediate care au ca rezultat o reducere mai mult sau mai puțin accentuată a producției și cu o posibilă depreciere a valorii produsului datorită deformărilor, decolorărilor și murdăriei din mierea de miere, exuvie , insecte. La acestea trebuie adăugate și daunele secundare ale vitalității reduse: reducerea resurselor energetice pentru plantă duce la o deteriorare progresivă care reduce rezistența sa naturală la alte adversități. În cele din urmă, plantele slăbite de atacuri intense și repetate ale afidelor sunt mai expuse la intrarea ciupercilor patogene și a insectelor xilofage .
Pagubele indirecte sunt după cum urmează:
- Propagarea virozei . Afidele sunt adesea vectori de viruși de tip purtat de stilet și, mai rar, de tip circulator. Posibilitatea de transmitere a virozelor este adesea o posibilitate mai înfricoșătoare decât daunele directe cauzate de afide, deoarece multe viroze pot provoca distrugerea totală a unei culturi sau a unei plantații.
- Producția de miere. La produzione di abbondanti secrezioni di melata rappresenta un danno indiretto sotto due differenti aspetti: da un lato la melata può indurre un deprezzamento del prodotto, che diventa appiccicoso e poco presentabile, da un altro favorisce l'insediamento dei funghi agenti della fumaggine . Pur trattandosi di funghi saprofiti, che non hanno perciò un rapporto diretto con la pianta, sono da considerarsi dannosi perché lo sviluppo del micelio riduce l'efficienza fotosintetica della pianta e produce un deprezzamento del prodotto.
Relazioni tra afidi e virus

Come si è detto in precedenza, gli afidi sono spesso vettori di virus fitopatogeni. L'importanza di questa relazione è notevole, al punto che la normativa fitosanitaria in materia di vivaismo impone la lotta preventiva agli afidi come misura di profilassi contro la diffusione delle virosi. Alcune specie polifaghe possono trasmettere anche oltre un centinaio di specie virali diverse [6] . Il meccanismo di trasmissione dipende dal tipo di virus: è persistente nel caso dei virus circolativi, non persistente nel caso di virus stylet-borne .
I virus stylet-borne basano la loro infettività su un meccanismo puramente meccanico e coinvolge gli afidi con nutrizione parenchimatica. Le particelle virali presenti in una pianta infetta imbrattano l'esterno degli stiletti boccali dell'afide che può quindi trasmetterlo per contatto meccanico nel momento in cui pungono una pianta sana. L'infettività è di tipo non persistente: infatti gli afidi perdono il virus nell'arco di poche ore o in occasione della muta . La propagazione da una pianta all'altra è affidata alle migranti su brevi distanze.
I virus circolativi basano la loro infettività su un meccanismo fisiologico e coinvolge gli afidi con nutrizione floematica. Le particelle virali presenti in una pianta infetta vengono assunte dal floema con la suzione della linfa; all'interno del corpo dell'insetto ed entrano nel circolo sanguigno passando nelle ghiandole salivari. La trasmissione alle piante sane avviene perciò mediante l'iniezione della saliva. La trasmissibilità varia in relazione al virus e al vettore ma è generalmente più corta che nei virus propagativi. La propagazione da una pianta all'altra può interessare anche grandi distanze.
Per quanto riguarda i virus propagativi: si tratta sempre di virus persistenti ma questi ultimi a differenza dei circolativi sono in grado di replicarsi all'interno degli afidi; questo porta ad un allungamento della lunghezza della trasmissibilità che dura per tutta la vita dell'insetto; in alcuni casi i virus entrano nelle gonadi rendendo infetta anche la progenie dei vettori, è facile comprendere come virus di questo tipo in presenza del giusto ambiente e dei giusti vettori portino quasi sempre allo sviluppo di epidemie.
Il rapporto tra virus e afide si può considerare di tipo mutualistico : è stato riscontrato che l'infezione virale provoca alterazioni di tipo fisiologico fra le quali è significativo un aumento della concentrazione di amminoacidi . Poiché la nutrizione azotata è strettamente correlata alla fecondità delle virginogenie, la presenza di un virus in una colonia ne stimola la moltiplicazione e la sovrappopolazione e la comparsa con maggiore frequenza delle virginogenie alate. Questo rapporto determina perciò un incremento del potenziale biologico dell'afide e della sua capacità di diffusione, favorendo nel frattempo la propagazione spaziale del virus [6] . Un altro aspetto che denota un mutualismo è l'innocuità del virus nei confronti dell'afide, al contrario di quanto avviene nel rapporto fra i virus circolativi ei Cicadellidi [6] .
Fattori di controllo

Come tutti gli organismi partenogenetici, dal punto di vista ecologico gli afidi sono un esempio classico di specie a strategia di tipo r , in grado cioè di raggiungere rapidamente livelli di popolazione elevati (pullulazioni) con crescita di tipo esponenziale. Le specie di questo tipo sono però facilmente controllate dal concorso di molteplici fattori intrinseci ed estrinseci. Questi ultimi possono distinguersi in fattori biotici e abiotici.
Tra i fattori interni hanno una particolare incidenza la competizione intraspecifica , causata dal sovraffollamento, e la comparsa delle virginogenie alate. In entrambi i casi il meccanismo di controllo è rappresentato da una riduzione della fecondità, alla quale si aggiunge l'elevata mortalità che colpisce le alate nelle fasi di migrazione.
Tra i fattori esterni di natura abiotica sono di particolare importanza quelli climatici. La temperatura è il fattore principale che provoca il drastico crollo delle popolazioni nelle regioni temperate, specie in quelle a inverno rigido. Nelle regioni con estati calde e asciutte il controllo sulle popolazioni è esercitato dalla riduzione della disponibilità alimentare. Per questi motivi le pullulazioni degli afidi sono particolarmente temibili, nelle nostre condizioni ambientali, soprattutto in primavera, in quanto l'elevato numero di germogli teneri e l'elevato tenore in azoto solubile nella linfa hanno un effetto stimolante sulla fecondità delle virginogenie. Tra i fattori climatici hanno un ruolo non trascurabile anche le precipitazioni ei forti venti, che interferiscono con le migrazioni delle alate. In ogni modo va considerato che gli agrosistemi deviano spesso dalle condizioni ambientali naturali e possono costituire microambienti favorevoli alle infestazioni degli afidi anche in piena estate. In tal senso ha un ruolo non trascurabile la duplice azione positiva delle concimazioni azotate e dell'irrigazione perché, stimolando il rigoglio vegetativo, hanno un effetto favorevole anche per le infestazioni degli afidi.

Tra i fattori biologici hanno un ruolo fondamentale gli insetti entomofagi . Fra i predatori sono particolarmente attivi sugli afidi i Coleotteri Coccinellidi , i Ditteri Sirfidi , i Neurotteri Crisopidi e diversi Rincoti Eterotteri ( Antocoridi , Miridi , Nabidi ). I predatori sono spesso ostacolati dalle Formiche, che tendono a proteggere le colonie allontanando i predatori. Fra i parassitoidi sono particolarmente importanti gli Imenotteri Braconidi ( Aphidiinae ) e Calcidoidi ( Aphelinidae ) ei Ditteri Cecidomidi .
Le dinamiche di popolazione degli Afidi e degli insetti ausiliari sono sfasate e gli entomofagi, pur essendo efficacissimi nel controllo biologico, fanno sentire il loro peso con uno spiccato ritardo (fine primavera, inizio estate), perciò da soli non sono in grado di controllare le infestazioni primaverili. In questo contesto ha un ruolo fondamentale la lotta integrata in quanto permette di intervenire in una fase critica senza pregiudicare l'evoluzione dei fattori naturali. La lotta a calendario , specie se condotta con principi attivi poco selettivi, ha invece effetti deleteri per i seguenti motivi:
- è efficace sulle prime infestazioni, ma può causare la comparsa di fenomeni di resistenza, rendendo difficile il controllo delle infestazioni successive, quando si abusa con trattamenti ripetuti a base di aficidi specifici;
- abbattendo le prime popolazioni sotto la soglia d'intervento causa un ritardo della dinamica di popolazione degli entomofagi;
- ha un impatto devastante sull'entomofauna utile quando s'impiegano prodotti a largo spettro d'azione.
In merito alla vegetazione spontanea, il comportamento va ponderato caso per caso in quanto da un lato si deve ostacolare lo sviluppo delle piante che possono rappresentare focolai d'infestazione ma nello stesso tempo si devono preservare quelle specie alle quali sono infeudati gli ospiti o le vittime secondarie dei parassitoidi e dei predatori.
Utilità

Nel panorama degli aspetti negativi legati agli Afidi, va citata anche un'utilità diretta nei confronti dell' apicoltura nei boschi montani di aghifoglie. In queste formazioni forestali la fonte principale di zuccheri per le api è rappresentata proprio dalla melata prodotta da alcuni afidi infeudati alle Conifere . Le popolazioni di questi afidi, in ambienti in cui è abbastanza tutelata la biodiversità , non raggiungono proporzioni tali da provocare danni economici ma rappresentano invece un'occasione d'integrazione dei redditi prodotti dalle forestazioni con la produzione di miele di melata . Parimenti, va specificato che, per le sue proprietà organolettiche (colore, gusto), questo prodotto dell'apicoltura non è particolarmente apprezzato dal mercato pertanto l'interesse in ambito nazionale è marginale.
Sistematica
Non c'è ancora una convergenza sulla sistematica interna degli Aphidoidea. Nelle vecchie classificazioni gli Afidi erano distinti in due raggruppamenti artificiali:
- Aphidoidea ovipara : comprendeva specie esclusivamente ovipare che fanno capo alle famiglie degli Adelgidae e dei Phylloxeridae .
- Aphidoidea vivi-ovipara : comprendeva specie vivipare o ovipare che fanno capo a diverse famiglie: Anoecidae , Aphididae , Chaitophoridae , Callaphididae , Hormaphididae , Lachnidae , Mindaridae , Pemphigidae (syn. Eriosomatidae ), Phloeomyzidae , Thelaxidae .
Il precedente schema è ritenuto da molti superato e ingloba nell'ambito degli Aphididae tutti gli Aphidoidea vivi-ovipara ripartendola in più sottofamiglie in linea di massima corrispondenti alle famiglie citate sopra.
- Famiglia Adelgidae
- Famiglia Aphididae
- Famiglia Phylloxeridae
Una classificazione ancora più recente scorpora il raggruppamento in due superfamiglie, Aphidoidea sensu strictu , comprendente la sola famiglia degli Aphididae, e Phylloxeroidea, comprendente gli Adelgidae ei Phylloxeridae.
Note
- ^ G. Cognetti, Endomeiosis in parthenogenetic lines of Aphids , in Cellular and Molecular Life Sciences , vol. 17, n. 4, 1961, pp. 168-169, DOI : 10.1007/BF02160362 , ISSN 1420-682X . URL consultato il 1º dicembre 2020 (archiviato dall' url originale il 29 aprile 2016) . Abstract.
- ^ KA Shufran, DC Peters, JA Webster, Generation of clonal diversity by sexual reproduction in the greenbug, Schizaphis graminum , in Insect Molecular Biology , vol. 6, n. 3, 2003, pp. 203-209, DOI : 10.1046/j.1365-2583.1997.00174.x .
- ^ Animali: scoperto negli afidi processo simile alla fotosintesi [ collegamento interrotto ] , in AGI.it , 23 agosto 2012. URL consultato il 24 agosto 2012 . ; Afidi: gli unici animali che sintetizzano carotenoidi , in Lescienze.it , 28 aprile 2010. URL consultato il 24 agosto 2012 .
- ^ Nancy A. Moran & Tyler Jarvik, Lateral transfer of genes from fungi underlies carotenoid production in aphids , in Science , vol. 328, n. 5978, 2010, pp. 624–627, DOI : 10.1126/science.1187113 , PMID 20431015 .
- ^ a b c d e Tremblay , pp. 111-112 .
- ^ a b c Tremblay , pp. 114-115 .
Bibliografia
- Ermenegildo Tremblay. Entomologia applicata . Volume II Parte I. 1ª ed. Napoli, Liguori Editore, 1981. ISBN 978-88-207-1025-5 .
- Antonio Servadei, Sergio Zangheri, Luigi Masutti. Entomologia generale ed applicata . Padova, CEDAM, 1972.
- Aldo Pollini. Manuale di entomologia applicata . Bologna, Edagricole, 2002. ISBN 88-506-3954-6 .
- Ângelo Moreira da Costa Lima.XXIII. Homópteros in Insetos do Brasil . Tomo 2. Escola Nacional de Agronomia, 1940. ( in portoghese ).
- Sebastiano Barbagallo (coordinamento). Gli afidi e le colture agrarie . REDA, 1985.
Altri progetti
-
 Wikizionario contiene il lemma di dizionario « Afidi »
Wikizionario contiene il lemma di dizionario « Afidi » -
 Wikimedia Commons contiene immagini o altri file su Afidi
Wikimedia Commons contiene immagini o altri file su Afidi -
 Wikispecies contiene informazioni su Afidi
Wikispecies contiene informazioni su Afidi
-
 Wikinotizie contiene l'articolo Venduto su eBay un insetto di 50 milioni di anni , 24 agosto 2008
Wikinotizie contiene l'articolo Venduto su eBay un insetto di 50 milioni di anni , 24 agosto 2008
Collegamenti esterni
- ( EN ) Aphidoidea , su Fossilworks.org .
- ( EN ) The Aphidoidea Web Page , su sel.barc.usda.gov , Systematic Entomology Laboratory, US Department of Agriculture. URL consultato il 7 ottobre 2008 (archiviato dall' url originale il 2 ottobre 2008) .
- ( EN ) David Stern, Aphidomorpha. Aphids, green flies, plant lice, adelgids, phylloxerids , su tolweb.org , The Tree of Life Web Project. URL consultato il 7 ottobre 2008 .
- ( EN ) Superfamily Aphidoidea , in Australian Faunal Directory , Australian Government, Department of the Environment, Water, Heritage and the Arts. URL consultato il 19 dicembre 2009 .
- ( EN ) Superfamily: Aphidoidea , su BugGuide . URL consultato il 7 ottobre 2008 .
- ( EN ) Superfamily: Phylloxeroidea , su BugGuide . URL consultato il 7 ottobre 2008 .
- ( EN ) Taxon details: Aphidoidea , in Fauna Europaea version 2.6.2 , Fauna Europaea Web Service, 2013. URL consultato il 07-10-2008 .
- ( EN ) Taxon details: Phylloxeroidea , in Fauna Europaea version 2.6.2 , Fauna Europaea Web Service, 2013. URL consultato il 07-10-2008 .
- ( EN ) ITIS Standard Report Page: Aphidoidea , in Integrated Taxonomic Information System . URL consultato il 07-10-2008 .
- ( EN ) ITIS Standard Report Page: Adelgoidea , in Integrated Taxonomic Information System . URL consultato il 07-10-2008 .
| Controllo di autorità | Thesaurus BNCF 33251 · LCCN ( EN ) sh85005957 · BNF ( FR ) cb12000650n (data) · NDL ( EN , JA ) 00560051 |
|---|