Neuroștiințe și orientare sexuală

Relația dintre neuroștiințe și orientarea sexuală a devenit un obiect de studiu în rândul cercetătorilor încă dintr-o perioadă relativ recentă, ultimul deceniu al secolului al XX-lea.
Orientarea sexuală este un model durabil de iubire romantică sau atracție sexuală (sau o combinație a acestora) față de persoane de același sex sau sex , de sex opus sau, în cele din urmă, către ambele sexe sau mai multe sexe [1] [2] .
Cauzele și mecanismele ultime ale dezvoltării sale în specia umană continuă să fie în mare parte neclare și multe dintre abordările teoretice avansate rămân în stadiul pur speculativ și nu încetează să devină subiectul controverselor în rândul experților.
Cu toate acestea, progresele și progresele notabile realizate de neuroștiințe încearcă să explice și să ilustreze și caracteristicile legate de orientarea sexuală individuală; studiile și cercetările efectuate până acum au explorat conexiunile neuronale structurale pestrițe, relațiile funcționale și / sau cognitive și în cele din urmă teoriile evolutive în raport cu orientarea sexuală umană.
Neurobiologia dezvoltării
Multe dintre abordările teoretice referitoare la dezvoltarea orientării sexuale implică dezvoltarea neuronală precoce a fătului , modelele propuse ilustrând expunerea la hormoni prenatali, imunitatea maternă și instabilitatea dezvoltării psiho-organice în sine. Alți dintre factorii propuși includ controlul orientării de către gene .
În orice caz, nicio propunere concludentă nu a ieșit la iveală sau nu a demonstrat faptul că efectele externe asupra mediului sau, în orice caz, învățate sunt singurul responsabil pentru orientarea non-heterosexuală [3] .
Modelul androgen prenatal
Dimorfismul sexual în creier și comportamentul în rândul vertebratelor se explică prin influența androgenilor steroizi emiși de gonade , așa cum s-a demonstrat pentru mai multe modele animale în ultimele decenii. Modelul prenatal al „androgenilor homosexualității ” vrea să descrie efectele neuro-evolutive ale expunerii fetale la această clasă de hormoni [3] .
Deja la mijlocul anilor 1980, cercetătorii Norman Geschwind și Albert Galaburda au adus ipoteza Geschwind-Galaburda în atenția mediului științific, potrivit căreia bărbații homosexuali au fost expuși la concentrații mari de androgeni în etapele foarte timpurii ale gestației, explicând, de asemenea, tendința lor de a fi mai susceptibile de a fi stânga mână , și , prin extensie , la toate aceste trasaturi hiper-masculinizare observate în cadrul acestui grup de populație [3] .
Mai recent s-a argumentat că mai presus de toate variațiile temporale și locale ale expunerii androgenilor la creier aflate încă în procesul de formare a fătului ar fi un factor decisiv în căile care într-o fază mai avansată determină sau nu identitatea de gen în ansamblu și în special homosexualitatea. Am trecut apoi la căutarea „markerilor somatici” decisivi pentru expunerea hormonală prenatală, care pot fi explorate cu ușurință și într-un mod complet neinvaziv la populațiile altfel în normă în ceea ce privește endocrinologia [3] .
Deoarece se știe că astfel de markeri sunt puternic influențați de hormonii sexuali prenatali, demonstrarea variațiilor reale ale acestor caracteristici între homosexuali și heterosexuali poate oferi o „fereastră” către neuro-dezvoltare timpurie a orientării sub influență hormonală. S-au găsit diverși indicatori somatici (incluzând „rapoartele degetelor 2D: 4D”, potențialele evocate auditiv, modelul d ’ amprentă digitală și genele de tip beat) care demonstrează orientarea bazată pe variațiile intervenite la persoanele adulte sănătoase [3] [4] .
Alte dovezi care susțin rolul testosteronului și al hormonilor prenatali includ observații ale subiecților masculini cu boala congenitală a exstrofiei cloacale care au fost repartizați sexului feminin la naștere, dar care au fost declarați mai târziu a fi bărbați. Acest fapt susține teoria conform căreia doza crescută de testosteron prenatal este crucială pentru dezvoltarea identității de gen. Mai mult, femeile - ale căror mame au fost expuse la dietilstilbestrol (DES) în timpul sarcinii au prezentat rate mai mari atât de bisexualitate, cât și de homosexualitate [5] .
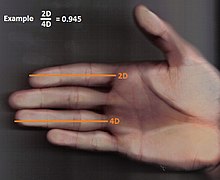
Raport 2D: 4D cifre
Cel mai bun indicator neinvaziv al expunerii hormonului prenatal este raportul cifrelor dintre a doua și a patra degetă a mâinii (prin urmare numit raport cifră 2D: 4D) [6] [7] , o măsură cunoscută a dimorfismului sexual: bărbații arată raporturi semnificativ mai mici decât femelele.
Pacienții cu supraexpunere la androgeni (cum ar fi hiperplazia suprarenală congenitală ) prezintă raporturi 2D: 4D mai mici, ceea ce oferă dovezi că expunerea la androgen prenatal este un factor cheie în această caracteristică.
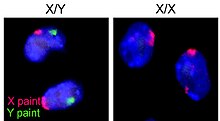
Indivizii XY cu sindrom de insensibilitate androgenă datorită unei gene disfuncționale a receptorilor androgeni - prezentă în identitățile de gen feminine și cu un raport numeric feminin - există exact așa cum s-ar putea prezice dacă hormonii androgeni afectează efectiv raporturile numerice. Această constatare demonstrează, de asemenea, că relația de diferențiere sexuală „în cifre” nu este de fapt strâns legată de cromozomul Y în sine [8] .
De asemenea, s-a arătat că raportul „2D: 4D” este influențat de variația genei receptorilor androgeni la bărbați [9] . Raportul dintre testosteron și estrogen în Amnios sa dovedit a fi corelat negativ cu raportul „2D: 4D” [3] .
Numeroase alte studii independente indică faptul că femeile lesbiene au „raporturi de cifre” masculinizate [10] [11] [12] [13] [14] [15] [16] [17] [18] [19] și că homosexualii în general cazurile relevă o relație care poate fi fie hiper-masculinizată, fie feminizată. Aceste rezultate par să întărească modelul de androgen prenatal: că expunerea netipică a hormonului este corelată cu dezvoltarea ulterioară a homosexualității umane [3] .
Potențiale evocative auditive
Studiile efectuate asupra procesării sistemului nervos central al senzației auditive și ale căror aspecte principale au fost legate de expunerea la androgeni prenatali, de testul stimulilor au arătat că femeile homosexuale au în general răspunsuri masculinizate, în timp ce, pe de altă parte, unii bărbați homosexuali au răspunsuri hiper-masculinizate [3] .
Efect asupra ordinii nașterii fraterne

Studiile făcute până în prezent arată că bărbații homosexuali au un procentaj mai mare de frați mai mari în comparație cu heterosexualii [20] , cel puțin conform expertului în sexologie Ray Blanchard . Această considerație a dus la descoperirea efectului ordinii de naștere fraterne, conform căreia cu cât frații mai mari (de la aceeași mamă), cu atât este mai mare posibilitatea exprimării unei orientări homosexuale conștiente atunci când devin adulți.
Estimările indică faptul că există o creștere de aproximativ 33% a probabilității de homosexualitate la un copil de sex masculin cu un frate mai mare [3] . Acest efect se dovedește a fi adevărat numai pentru frații biologici; în timp ce șansele homosexualității masculine nu sunt crescute cu numărul fraților mai în vârstă sau al fraților adoptivi [21] . Se estimează că 1/7 din toți bărbații homosexuali își datorează orientarea sexuală efectului ordinii de naștere fraterne [22] [23] [24] .

Efectul se aplică numai bărbaților homosexuali dreptaci; pe de altă parte, probabilitatea homosexualității la bărbații stângaci sau ambidextri nu crește [26] .
Având în vedere că efectul are un rezultat contingent asupra abilităților manuale și că acesta corespunde unei trăsături prenatale determinate, efectul ordinii nașterii este interpretat a fi mai degrabă de natură biologică decât psihosocială; prin urmare, există în natură și se știe că funcționează în perioada prenatală [21] .
În biologia umană, dexteritatea reprezintă o performanță mai bună, mai rapidă sau mai precisă sau o preferință individuală pentru utilizarea unei mâini, cunoscută sub numele de mână dominantă; mâna mai puțin capabilă sau mai puțin preferată se numește mână nedominantă [25] [27] [28] . Bărbații sunt mai predispuși să exprime o stânga puternic dominantă decât femeile [29] ; se estimează că între 70 și 95% din populația lumii este dreptaci.
În orice caz, mecanismul biologic exact prin care efectul funcționează pe parcursul vieții fetale nu este confirmat cu o certitudine dincolo de orice îndoială. Mecanismul propus prin care se crede că funcționează efectul afirmă că o mamă dezvoltă un răspuns imun împotriva unei substanțe importante în dezvoltarea fetală masculină în timpul sarcinii și că un astfel de efect imunitar devine din ce în ce mai probabil la fiecare făt masculin ulterior [21] .
Prin urmare, se crede că efectul imun provoacă o modificare a dezvoltării creierului prenatal al bărbaților, cu o creștere statistică directă pentru fiecare gestație a unui copil de sex masculin. Ținta răspunsului imun poate fi moleculară (adică proteine legate de cromozomul Y ) la suprafața celulelor creierului fetal masculin, inclusiv locurile unde se află hipotalamusul anterior care a fost legat de orientarea sexuală în alte cercetări [21] .
Tindem să credem că anticorpii produși în timpul răspunsului imun traversează bariera reprezentată de placentă și intră în compartimentul fetal unde se leagă de moleculele Y, modificându-și astfel rolul în diferențierea sexuală și conducând unii bărbați născuți mai târziu să încerce o atracție mai puternică. față de bărbați decât față de femei [21] .
Mecanismul propus continuă să aibă doar dovezi indirecte care să îl susțină; este, de asemenea, singurul mecanism plauzibil propus până acum de comunitatea științifică pentru a explica cum și în ce fel poate opera efectul ordinii nașterii fraterne în uter [21] . Efectul nu se aplică dezvoltării homosexualității feminine [21] .
Instabilitate evolutivă și preferință manuală
Instabilitatea evolutivă se referă la gradul de vulnerabilitate al unui organism atât la stresul de mediu cât și la cel genetic în timpul perioadei de dezvoltare a acestuia [3] [30] [31] ; măsurătorile unei astfel de instabilități pot furniza unele informații despre întreaga istorie evolutivă a organismului însuși.
Asimetria în caracteristicile corporale bilaterale, cunoscută sub numele de asimetrie fluctuantă , este adesea utilizată în cercetarea științifică ca măsură substitutivă a instabilității dezvoltării; el este, de asemenea, înclinat să considere că constanta non-dreaptă (adică o predispoziție la o dezvoltare mai mare a părții stângi a corpului uman ) constituie una dintre măsurile care definesc instabilitatea dezvoltării [30] .

Șansele de a fi caracterizat prin stânga handedness au potențialul de a fi crescută la homosexuali populații. În comparație cu un eșantion de bază heterosexual , o meta-analiză efectuată în 2000 de Lalumiere și colab a unei game întregi de studii anterioare [31] a arătat că bărbații homosexuali sunt cu aproximativ o treime (34%) mai predispuși să nu aibă dreptate -manate, în timp ce pentru femeile lesbiene procentul devine aproape dublu (cu până la 91% mai probabil să fie așa în unele cazuri) [31] .
Ce explicație s-a propus că lipsa „dexterității” [ necorespunzătoare lexicală ] (inclusiv ambidextruism ) este legată în mod corespunzător de homosexualitate prin instabilitate evolutivă [26] ; conform acestei ipoteze, orientarea homosexuală s-ar datora, prin urmare, factorilor de dezvoltare generalizați (adică factori genetici de mediu sau non-hormonali) care schimbă preferințele erotice de la modelul tipic de atracție către sexul opus, începând de la neuro-dezvoltarea fătului [3]. ] [30] .
Dacă această logică a raționamentului ar fi corectă, ar fi de așteptat ca homosexualitatea masculină și feminină să fie asociată și cu alte semne distinctive ale instabilității dezvoltării, cum ar fi asimetria fluctuantă crescută (de exemplu, în amprentele digitale și lungimea urechii )., Încheietura mâinii , degetul și piciorul. ) [3] [30] .
Cu toate acestea, mai multe studii nu au găsit nicio diferență semnificativă în asimetria fluctuantă între heterosexuali și homosexuali [3] [14] [32] [33] , sugerând astfel că homosexualitatea nu reprezintă de fapt o orientare sexuală fenotipică „suboptimală” [3] ; dimpotrivă, s-a constatat că bărbații homosexuali și femeile lesbiene prezintă o asimetrie mai puțin fluctuantă (și, prin urmare, mai puțină instabilitate de dezvoltare) decât bărbații sau femeile heterosexuale, nu mai mult.
Cu alte cuvinte, heterosexualii au o capacitate genomică mai slabă decât homosexualii de a tampona cu succes dezvoltarea și, în consecință, obțin un fenotip normal în condiții de mediu imperfecte[34] [35] . Mai mult, ( „masculine la feminin“ FTM ) homosexuali transsexualilor (adică care sunt atrași de femei) au prezentat o mai mare fluctuantă asimetrie decât ambele grupuri de homosexuali și de control, ceea ce demonstrează că instabilitatea poate explica variatiile identității de gen , dar nu orientare per se [30 ] .

De asemenea, s-a constatat că bărbații homosexuali sunt în general considerați (sub formă de stereotip ) mult mai atrăgători decât bărbații heterosexuali și acest lucru se întâmplă chiar și atunci când evaluatorii nu cunosc orientările sexuale ale bărbaților testați; în timp ce în timpul copilăriei, evaluatorii independenți descriu în mare parte „băieții atipici” (care nu se conformează regulilor sau „ambiguu”) ca fiind mult mai atrăgători decât băieții tipici de genul lor (vezi masculinitatea ) [30] [36] [37] [38] .
În concluzie, ceva din fizionomia copiilor homosexuali , adolescenților și adulților homosexuali îi deosebește pentru a fi considerați mai atrăgători decât toți ceilalți bărbați heteroși: s-a sugerat că acest lucru se datorează faptului că bărbații homosexuali au o asimetrie până la oscilație scăzută. , despre care se știe că este asociat cu o atractivitate fizică crescută [30] [39] [40] .
Prin urmare, mecanismul general propus în calculul instabilității de dezvoltare a lui Lalumiere și al altor persoane este, prin urmare, atât pentru homosexualitatea masculină, cât și pentru cea feminină - incompatibil cu varianța sugerată de dovezile pentru diferitele căi ale homosexualității masculine față de femei [3] [41] .
Cu toate acestea, ipoteza a fost criticată și pentru că este prea generală și pentru că nu a precizat exact ce mecanisme specifice de dezvoltare evolutivă ar fi probabil perturbate [30] ; prin urmare, canalizarea trăsăturii date de orientarea sexuală este mai probabil să fie cauzată de mecanisme specifice de dezvoltare neurologică (care pot include acțiunile androgenului în timpul fazei prenatale), mai degrabă decât de mecanisme generale de dezvoltare neurologică (ca într-adevăr instabilitate de dezvoltare) [3] .
O problemă semnificativă pusă cu măsurarea asimetriei fluctuante este că multe dintre caracteristicile anatomice utilizate arată, de asemenea, un anumit grad de asimetrie direcțională (adică o abatere normală de dezvoltare de la simetrie care merge în aceeași direcție la majoritatea indivizilor ca poziția inimii în stânga latura pieptului ) a cărei direcție și lățime pot varia atât între sexele M / F, cât și între homosexuali și heterosexuali (primii au o asimetrie direcțională comparativ mai mare decât heterosexualii) [14][34] .
Dacă această asimetrie direcțională nu este evaluată cu atenție și eliminată din datele pentru fiecare grup de subiecți, este foarte probabil ca măsura să nu fie în cele din urmă corectă; când seturile de date ale studiilor publicate anterior (care păreau să arate o asimetrie fluctuantă crescută la homosexuali) au fost reanalizate în lumina acestei probleme, a apărut în cele din urmă că homosexualii prezintă o asimetrie fluctuant semnificativ mai mică decât heterosexualii[34] .
Acest lucru sugerează apoi că homosexualii experimentează de fapt o instabilitate de dezvoltare mai mică decât heterosexualii.
Diferențe structurale
Studiile de imagistică post-mortem și de spectrometrie de masă din ultimele două decenii au relevat diferențe structurale atât în structurile cerebrale globale, cât și mai adecvate legate de sexualitate între subiecții heterosexuali și homosexuali.

Hipotalamus
Hipotalamusul este o porțiune a creierului care conține un anumit număr de „nuclei” (grupuri discrete de corpuri celulare în soma neuronului ).
Acum, termenul „nucleu” din neuroanatomie nu trebuie confundat cu aceeași utilizare făcută din acesta în biologia celulară : în al doilea caz se referă la organele găsite în celulele eucariote care conțin materialul genetic al celulei ; în timp ce în prima se referă la grupuri discrete de corpuri de celule neuronale dens împachetate în sistemul nervos central [42] [43] . În secțiunile anatomice, un nucleu apare ca o regiune de substanță cenușie înconjurată de substanță albă .
Se știe că este implicat în diferențe sexuale în comportamentul reproductiv, mediantă a răspunsurilor ciclului menstrual : în special hipotalamusul anterior ajută la reglarea comportamentului sexual tipic bărbaților. La mijlocul anilor 1990, acesta era legat și de identitatea de gen și orientarea sexuală [44] .

Cercetările seminale efectuate de Simon LeVay ar fi descoperit că un nucleu interstițial al hipotalamusului, INAH3 , era dimorf în funcție de orientarea sexuală, dar nu în funcție de sex . Mai exact, INAH3 al bărbaților homosexuali sa dovedit a fi mai mic ca volum decât cel al bărbaților heterosexuali; aceste rezultate au fost obținute din analiza post-mortem a nucleelor hipotalamice ale subiecților homosexuali cunoscuți comparativ cu pacienții heterosexuali [45] .
Cercetările ulterioare au constatat că INAH3 are un volum mai mic la bărbații homosexuali decât la bărbații heterosexuali, acest lucru se datorează faptului că primii au o densitate neuronală mai mare în interiorul acestuia decât al doilea; cu toate acestea, nu există nicio diferență în numărul sau aria secțiunii transversale a neuronilor din INH3 a bărbaților homosexuali comparativ cu heterosexualii [46] . De asemenea, s-a constatat că nu există un efect constatat de infecția cu HIV asupra dimensiunii INAH3, adică nu ia în considerare diferența de volum observată între bărbații homosexuali și heterosexuali [46] .
Hipotalamusul este, de asemenea, legat de orientarea sexuală prin constatări care arată că activitatea aromatazei - o enzimă importantă care transformă androgenul în estrogen - este crescută în regiunea hipotalamică pre-optică a mamiferului în perioada pre- și neonatală . Acest lucru este într-adevăr legat de diferențierea sexuală și poate constitui o bază pentru diferențele sexuale structurale și funcționale care joacă un rol în medierea dezvoltării orientării datorită expunerii hormonale prenatale [44] .

Nucleul suprachiasmatic (SCN) al hipotalamusului anterior se referă și la orientarea sexuală, fiind mai mare și mai alungit la bărbații homosexuali decât la bărbații și femeile heterosexuale. Sub- nucleul celular care conține vasopresină al SCN al bărbaților homosexuali este de două ori mai mare și are de 2,1 ori numărul de celule decât subgrupul care conține vasopresină al SCN la bărbații heterosexuali [44] [47] .

Aceasta ar putea fi o explicație neurologică pentru descoperirea că bărbații homosexuali se trezesc și se retrag zilnic mai devreme decât heterosexualii, deoarece se știe că SCN este implicat în modularea ritmurilor circadiene umane [44] .
În mod similar, într-un studiu model de șobolan , șobolanii masculi tratați cu un inhibitor de aromatază s-au dovedit a prefera femeile atunci când au fost testați în faza târzie, dar au arătat preferințe de împerechere homosexuale atunci când au fost testați în faza întunecată timpurie. orientare la alte specii [44] .
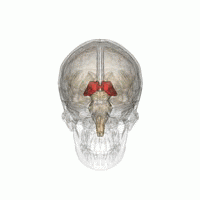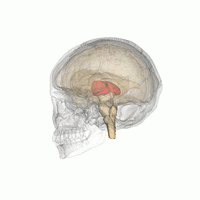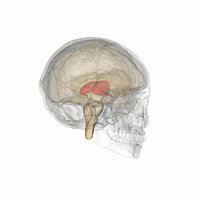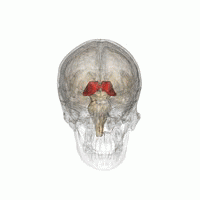
Talamus
Talamusul este o structură ovoidă simetrică a liniei medii din creierul uman, situată între cortexul cerebral și creierul mediu în ambele emisfere cerebrale . Un studiu de imagistică prin rezonanță magnetică a comparat volumele subcorticale ale bărbaților homosexuali și heterosexuali; a constatat că, în timp ce ambele grupuri nu au diferit în ceea ce privește volumul total al creierului, volumul de talamus (în ambele emisfere) a fost mai mare la bărbații heterosexuali [48] .
Un alt studiu a raportat că conectivitatea funcțională care implică talamusul drept și cuneul drept a fost diferită între bărbații homosexuali și heterosexuali și a arătat, de asemenea, corelații cu scorurile scării Kinsey [49] ; în plus, talamusul este implicat în procesul de excitare și recompensă sexuală ; în timpul excitării evocate vizual, atât bărbații heterosexuali, cât și homosexualii au activat talamusul, dar spre deosebire de acesta din urmă, heterosexualii au arătat o activare suplimentară în girusul lingual [48] .

Nucleul bazal al stria terminalis
Nucleul bazal al striei terminale (BNST) este o zonă a sistemului limbic al creierului anterior care este implicată în controlul comportamentului de împerechere; primește intrări neuronale de la amigdala medială și bulbul olfactiv accesoriu și trimite proiecții atât către zona pre-optică medială, cât și către nucleul ventro-medial al hipotalamusului [50] [51] .
Partea centrală a BNST (BNSTc) este cu 44% mai mare la bărbații heterosexuali decât la femeile heterosexuale și 62% la bărbații homosexuali comparativ cu aceștia [52] . BNSTc este mai mare la bărbații homosexuali decât la bărbații heterosexuali, deși diferența de mărime nu este semnificativă statistic [52] . Prin urmare, se presupune că BNSTc al bărbaților homosexuali este „hiper-masculinizat”, deoarece este mai mare decât BNSTc al bărbaților și femeilor heterosexuale [51] .
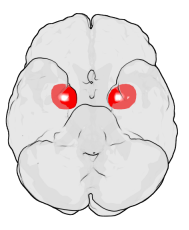
Amigdala
S-a constatat că atât bărbații și femeile homosexuale prezintă conexiuni cu amigdala diferite de cele ale bărbaților și femeilor heterosexuale.
Mai exact, legăturile dintre bărbații homosexuali și femeile heteroase erau mai răspândite în amigdala stângă, în timp ce conexiunile funcționale erau mai răspândite în amigdala dreaptă la bărbații heteroși și lesbiene [53] [54] .
Comisie anterioară

Comisura anterioară , un pachet de fibre de substanță albă care leagă cele două emisfere cerebrale, a fost găsită de Allen și Gorski ca fiind semnificativ mai mare la bărbații homosexuali și la femeile heterosexuale decât la bărbații heterosexuali [44] .
Această constatare oferă o posibilă bază anatomică pentru conexiuni funcționale interhemisferice mai mari la homosexuali, ceea ce explică de ce bărbații homosexuali și femeile heterosexuale prezintă o simetrie funcțională marcată a circuitului lingvistic comparativ cu bărbații heterosexuali care efectuează aceleași teste verbale [53] .
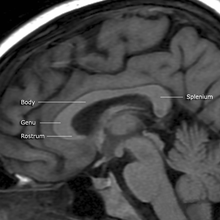
corp calos
Corpul calos (CC), precum și comisura anterioară, este o legătură neuronală importantă care leagă cele două emisfere; cu toate acestea, spre deosebire de comisură (care este prezentă la toate tipurile de vertebrate ), CC este prezentă numai la animalele placentare (inclusiv la oameni) [56] .
Un studiu RMN care compara CC al bărbaților homosexuali și heterosexuali a constatat că toate părțile CC sunt mai mari la persoanele gay[57] .
În special, istmul (o parte a CC prezentă între corpul calos și mușchiul splenius al capului ) este semnificativ mai mare la bărbații homosexuali decât la heterosexuali [55] [56] ; Mărimea CC are o bază genetică puternică, cu rate de moștenire genetică variind de la 82 la 94% [55] . Această asociere a orientării sexuale cu o structură cerebrală foarte ereditară susține teza unei baze genetice și neurobiologice la originea aceleiași orientări [55] .

Substanță gri
Materia cenușie este o parte importantă a sistemului nervos central, care este compus în principal din corpuri de celule neuronale. În timp ce bărbații au, în general, o cantitate mai mare de substanță cenușie și albă decât femeile (datorită masei corporale mai mari a bărbaților și, în consecință, a unei dimensiuni mai mari a creierului), femeile au, în general, un raport mai mare substanță-materie cenușie și straturi mai groase ale acesteia în zone specifice de cortexul cerebral în raport cu bărbații [58] [59] .
S-a constatat că femeile homosexuale au substanțe gri relativ mai puține decât femeile heterosexuale din zona cerebelului ventral , cortexul premotor ventral stâng, cortexul cerebral temporal-bazal și, cel mai semnificativ, cortexul peririnal stâng al lobului temporal . Nu s-a găsit nicio diferență în cantitatea de substanță cenușie între bărbații heteroși și homosexuali [58] .
Questi risultati sono importanti perché la corteccia peririnale si trova vicino alle regioni del cervello ( corteccia entorinale , ippocampo , giro del paraippocampo e amigdala ) coinvolti nell'elaborazione olfattiva e spaziale, che hanno dimostrato di determinare differenze nell'orientamento sessuale - in particolare, sono note nelle donne omosessuali prestazioni superiori alle donne etero nei test di elaborazione spaziale [54] [58] .
La corteccia peririnale stessa è coinvolta in funzioni legate all'elaborazione di stimoli sessuali come l'elaborazione olfattiva, la codifica della memoria e la stessa elaborazione spaziale; è anche coinvolta nel rilevamento dell'identità dell'oggetto. È noto che essa modifica l' attrazione sessuale negli esseri umani e il sistema olfattivo è in grado di differenziare i composti simili al feromone in base all'orientamento sessuale [58] .
La scoperta che le donne omosessuali hanno un modello GM "maschile" mentre di contro gli uomini omosessuali non hanno un modello "femminile" indica che l'omosessualità maschile e il lesbismo non si manifestano allo stesso modo a livello strutturale nel cervello [58] .
Inoltre altre scoperte di caratteristiche sessualmente dimorfiche che sono più maschili nelle donne omosessuali, ma non femminili negli uomini gay , includono le emissioni oto-acustiche, il rapporto dito 2D: 4D e la corporatura. Complessivamente questi risultati suggeriscono che i livelli sessuali atipici dell'azione androgena prenatale possono essere coinvolti nell'origine dell'omosessualità femminile [58] .

Asimmetria cerebrale
La dimensione del telencefalo è un tratto sessualmente dimorfico in cui gli uomini tendono a mostrare asimmetria nei volumi dei loro emisferi, mentre le donne mostrano invece una simmetria volumetrica. È anche un tratto che è assai improbabile possa essere influenzato da schemi appresi socio-ambientali [53] .
Uno studio volumetrico con risonanza magnetica del 2008 ha indicato che uomini gay e donne eterosessuali mostravano volumi emisferici simmetrici mentre le donne omosessuali e gli uomini etero mostravano un'asimmetria verso destra. Questi risultati dimostrano una differenza neurologica globale nelle strutture cerebrali che mostrano caratteristiche sessuali atipiche associate all'orientamento sessuale [54] .
Corteccia cerebrale

La corteccia cerebrale è lo strato più esterno del cervello umano ed è composta da tessuto nervoso . Uno studio RM ha confrontato lo spessore corticale in varie regioni del cervello di uomini omosessuali, uomini eterosessuali e donne eterosessuali: ha scoperto che gli uomini omosessuali avevano cortecce più sottili - rispetto a quelli etero - nella regione laterale orbito-frontale dell'emisfero destro, nonché nelle regioni situate nella corteccia visiva (linguale, pericalcarina e cuneo ) [48] .
Le stesse regioni hanno mostrato una corteccia più sottile nelle donne eterosessuali rispetto agli uomini etero, mentre non sono state riscontrate differenze tra donne eterosessuali e uomini omosessuali. Maschi gay ed etero non differivano nei volumi totali del cervello ed è stato determinato che le differenze riportate nello spessore corticale non erano influenzate dagli anni di educazione o dal volume cerebrale dei soggetti [48] .
Poiché le regioni sopra menzionate mostrano dimorfismo sessuale , gli autori hanno ipotizzato che i processi biologici frequentemente proposti per sottostimare lo stesso, come i meccanismi dipendenti dal gene e dall' ormone sessuale durante lo sviluppo prenatale e postnatale, possano interagire con l'architettura corticale nelle aree visive risultando in differenti spessori corticali nei gay rispetto agli etero [48] .

Inoltre gli uomini omosessuali mostravano cortecce più sottili rispetto agli uomini e alle donne etero sia nella triangular pars destra (nota anche come area 45 di Brodmann ) che nelle regioni temporali inferiori; ciò suggerisce che le differenze cerebrali legate all'omosessualità maschile possono anche essere presenti in regioni che non sono necessariamente considerate come dimorfiche sessuali [48] .

Un altro studio ha dimostrato che lo spessore corticale del triangolare destro pars differisce anche tra le transessuali MtF e gli uomini gay.
Nello specifico la pars triangularis delle persone MtF (e degli uomini eterosessuali) è più spessa rispetto a quella dei gay; inoltre nelle MtF è più spessa anche rispetto a quella degli uomini etero. In particolare, in entrambi gli studi, la regione interessata è la pars triangularis presente nell'emisfero destro) [60] .
Differenze funzionali
Diversi studi hanno iniziato ad esplorare i substrati funzionali e cognitivi dell'orientamento sessuale, in definitiva come una manifestazione comportamentale; l'elaborazione neurale in risposta a stimoli specifici e compiti cognitivi sessualmente "di genere" sono stati individuati per essere correlati con lo specifico orientamento assunto da ogni individuo .
Risposta ai feromoni
Due feromoni umani proposti - il derivato del progesterone 4,16-androstadien-3-one (AND) e un estere -1,3-5 (10), 16-tetraen-3-olo (EST) ( steroide similare all' estrogeno - hanno dimostrato di avere risposte specifiche all'orientamento sessuale nell'attivazione dei circuiti neurali dell' ipotalamo anteriore in soggetti sia omosessuali che eterosessuali. L'ipotalamo anteriore è coinvolto nell'elaborazione delle funzioni riproduttive e recenti evidenze suggeriscono che aiuti a integrare gli stimoli ormonali e sensoriali coinvolti nel comportamento sessuale e nelle sue preferenze [61] .
Recenti esperimenti di imaging a risonanza magnetica funzionale hanno mostrato che la presentazione di AND, trovata nella sudorazione maschile, come stimolo olfattivo produceva risposte olfattive normali in uomini etero e lesbiche , mentre attivava l'ipotalamo anteriore in uomini gay e donne etero [62] .
La proposta EST del feromone, trovata nell' urina delle donne in stato di gravidanza , produce una normale attivazione olfattiva negli uomini gay e nelle donne eterosessuali, mentre d'altra parte le lesbiche e gli uomini etero hanno dimostrato avere risposte ipotalamiche sessualmente correlate [61] .
Gli uomini gay hanno mostrato le stesse risposte funzionali sessualmente correlate a questi stimoli delle donne eterosessuali, mentre le donne omosessuali hanno risposto come gli uomini etero. Questa ricerca condotta da Berglund e Savic indica nel complesso che AND ed EST inducono " effetti specifici della sessualità sul sistema nervoso autonomo " e che gli stimoli hanno prodotto un percorso di risposta che dipendeva dall'orientamento sessuale del soggetto piuttosto che dal sesso derivante dal fenotipo [62] .
Risposta agli stimoli sessuali visivi

L' eccitazione sessuale è un processo altamente coordinato che prepara una persona al comportamento riproduttivo; i cambiamenti diffusi avvengono nello stato neurofisiologico della persona durante l'eccitazione per ottenere risposte adattive. I sistemi dell'attenzione, affettivi e motivazionali dell'individuo interessato vengono ottimizzati in modo tale da consentire la selezione e l'impiego con successo degli stimoli sessuali.
In risposta agli stimoli sessuali visivi gli uomini mostrano eccitamento soggettivo e auto-riferito di categoria specifico; la loro maggiore eccitazione viene rivolta a quelle categorie di persone con cui preferiscono intrattenere un rapporto sessuale : gli uomini omosessuali provano maggiore eccitazione genitale e soggettiva per gli uomini che per le donne (e quindi preferiscono gli stimoli sessuali maschili), mentre per gli uomini eterosessuali avviene l'inverso.
Si ritiene che l' ormone influenzi nel periodo prenatale lo sviluppo delle strutture neurali che regolano il comportamento sessuale; quindi si ritiene che alcuni aspetti dello sviluppo neuro-ormonale negli omosessuali procedano in modo diverso dagli eterosessuali, con conseguenti differenze psicologiche come inneschi distinti (o "stimoli") per l'eccitazione sessuale [63] .
Uno studio del 2007 sulla risonanza magnetica funzionale (fMRI) che ha esplorato i meccanismi neurali dell'eccitazione sessuale negli uomini gay ed etero ha mostrato ai loro soggetti interazioni sessuali composite; hanno dimostrato che entrambi i gruppi maschili attivano le stesse regioni cerebrali dopo che ciascuna è esposta ad uno stimolo sessuale concordante con l' orientamento sessuale del soggetto preso in esame [63] .

Un altro studio di fMRI ha dimostrato che osservando sia gli stimoli visivi erotici etero che quelli gay, solamente quei video corrispondenti all'orientamento sessuale del soggetto producevano modelli di attivazione nelle aree del cervello associate all'eccitazione sessuale. La risposta degli eterosessuali mostrava lo stesso schema dell'elaborazione sessuale neurale la quale provocava la visione gay, mentre la visualizzazione delle immagini dell'orientamento opposto non suscitava la stessa risposta [64] .
È stata quindi riscontrata una correlazione significativa tra l'eccitazione e l'attivazione neurale nell' ipotalamo , una regione chiave del cervello umano per la sua funzione sessuale; anche i valori di eccitazione sessuale auto-riportati erano uguali in entrambi i gruppi. Tuttavia l'entità dell'attivazione ipotalamica era inferiore negli uomini gay rispetto a quelli etero, una caratteristica che è condivisa anche dalle donne etero [64] .
Un ulteriore studio fMRI ha determinato gli schemi di attivazione cerebrale nei soggetti omosessuali ed eterosessuali, esponendoli a stimoli visivi gay, etero e lesbici; hanno quindi rilevato che diversi circuiti neuronali erano attivi nei due gruppi maschili: regioni cerebrali come la circonvoluzione angolare sinistra, il globo pallido destro e il nucleo del caudato sinistro venivano attivate esclusivamente negli uomini omosessuali mentre il giro bilaterale linguale , il giro destro parahippocampale e l' ippocampo destro erano attivato esclusivamente in uomini eterosessuali [65] .
Questi risultati indicano che i circuiti neurali (correlati all'elaborazione di stimoli sessuali visivi) che sono attivi durante l'eccitazione sessuale negli uomini omosessuali ed eterosessuali sono differenti [65] .
Una nuova ricerca fMRI ha mostrato a donne e uomini eterosessuali e omosessuali foto di genitali maschili e genitali femminili ; limitando quindi lo stimolo sessuale visivo alle fotografie dei genitali gli autori hanno ridotto al minimo l'attività neuronale correlata all'elaborazione di vari stimoli quali volti, voci, movimenti corporei e parti del corpo sessualmente eccitanti oltre ai genitali [66] .
Hanno scoperto che lo striato ventrale, il talamo centromediano e la corteccia premotoria ventrale bilaterale hanno mostrato una risposta più forte alle foto del sesso preferito rispetto a quelle corrispondenti del sesso non preferito. Poiché lo striato ventrale e il talamo centromediano sono noti per essere attivati da preferenze innate, la risposta selettiva di queste regioni agli stimoli sessuali preferiti sembra riflettere un modello di risposta predeterminato. Questa nozione è quindi usata a supporto come una delle prove che vogliono l' orientamento sessuale essere di origine prettamente biologica [66] .
Un altro studio di FMRI ha cercato di verificare se i soggetti rispondessero più ai volti (maschio o femmina) a cui erano sessualmente orientati e predissero tale modulazione nel circuito cerebrale del sistema di ricompensa . A uomini e donne eterosessuali e omosessuali sono state mostrate foto di volti maschili e femminili e quindi invitati a valutarne l'attrattiva visiva. Coerentemente con l'ipotesi si scoprì che il circuito di ricompensa dei maschi omosessuali e delle femmine eterosessuali rispondeva di più alle fotografie di volti maschili, mentre i circuiti di ricompensa delle femmine omosessuali e dei maschi eterosessuali rispondevano di più alle fotografie che mostravano volti femminili [67] .
L'interazione tra genere di stimolo (volto maschile o femminile) e orientamento sessuale (omosessuale o eterosessuale) del soggetto era altamente significativa in due regioni cerebrali: il nucleo mediodorsale del talamo (MDT) e la corteccia orbitofrontale mediale (OFC). L'attivazione nell'OFC è notevole perché esso è coinvolto nella rappresentazione del valore di ricompensa di vari stimoli sensoriali, compresi i volti attraenti. Sembra anche che abbia un ruolo importante nell'elaborazione dei segnali facciali necessari per la comunicazione sociale, poiché questa regione possiede neuroni selettivi per il viso e perché i pazienti con lesioni OFC non sono in grado di identificare le espressioni facciali emotive [67] .
La modulazione della risposta ai volti all'interno dell'OFC attraverso l'orientamento sessuale aggiunge ulteriore importanza al suo ruolo nel comportamento sociale; poiché mdT e OFC ricevono le proiezioni neurali l'una dall'altra i modelli di attivazione simili osservati in queste regioni possono essere attribuiti alle loro connessioni anatomiche [67] .
Asimmetria cerebrale funzionale
Differenze nell'elaborazione neurale e nei compiti cognitivi sono state trovate in relazione all'orientamento sessuale. In una recensione del 1987 sulla cognizione, sulla lateralizzazione cerebrale e sull'orientamento, Sanders e Ross-Field hanno suggerito che gli eventi ormonali prenatali porterebbero a asimmetrie cerebrali funzionali correlate allo stesso orientamento [68] .
Alcuni compiti cognitivi sono noti per essere sessualmente dimorfici. La migliore abilità verbale delle donne è associata a una ridotta lateralizzazione delle attività linguistiche, mentre il vantaggio maschile nei compiti spaziali corrisponde alla marcata lateralizzazione cerebrale. Effetti di orientamento sessuale in alcuni di questi compiti sono stati rilevati in diversi studi.
Nel test di Vincent Mechanical Diagrams, una misura del campo di rilevazione del punto divisa dell'asimmetria cerebrale funzionale, gli uomini gay conseguivano lo stesso risultato delle donne eterosessuali ed entrambi con punteggi inferiori rispetto agli uomini etero che mostravano meno asimmetria. Inoltre gli uomini omosessuali mostrano punteggi di Quoziente d'intelligenza delle prestazioni verbali più alti nei sotto-test della scala di Intelligenza degli adulti di Wechsler, in concordanza con i modelli di test eseguiti dalle donne [68] .
In molti altri test, che includevano un compito di lancio mirato da parte del maschio e uno dei test neuropsicologici (il "Purdue Pegboard") con parzialità femminile, le prestazioni degli uomini gay e delle donne etero non mostravano alcuna rilevante differenza statistica gli uni dalle altre, mentre entrambi differivano significativamente dagli uomini eterosessuali [68] .
Inoltre una riduzione dell'asimmetria è stata rinvenuta in uno studio magneto-encefalografico in cui si rileva che le stime di localizzazione di origine di un segnale uditivo evocato da MEG sono emisfericamente simmetriche nelle donne eterosessuali e negli uomini omosessuali, mentre sono asimmetriche negli uomini etero [68] .
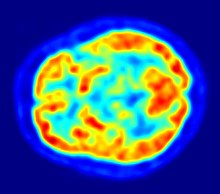
Risposta alla serotonina
La serotonina è un neurotrasmettitore trovato nel sistema nervoso centrale che ha vari ruoli nella regolazione del comportamento sessuale; i suoi agonisti e antagonisti hanno effetti attivanti o inibitori a seconda della loro concentrazione e dell'area cerebrale coinvolta. La fluoxetina è un inibitore selettivo della ricaptazione della serotonina che prolunga l'effetto della stessa sui neuroni [69] .
Kinnunen e altri (2004) hanno somministrato fluoxetina ai loro soggetti di studio per vedere se il cervello viene attivato in una maniera differente negli uomini omosessuali ed eterosessuali attraverso l'azione della serotonina [69][70] ; dopo somministrazione di fluoroxina hanno misurato il metabolismo del glucosio ( fluorodesossiglucosio ) nel cervello utilizzando la tomografia a emissione di positroni (FDG-PET).
Hanno rilevato che la risposta cerebrale alla fluoxetina differisce tra le persone gay e gli uomini etero, cioè i primi mostrano una minore riduzione del metabolismo del glucosio nell' ipotalamo rispetto ai secondi. Inoltre anche altre aree del cervello sono state attivate in modo differenziale: la corteccia prefrontale associativa degli uomini omosessuali mostrava una maggiore attività dopo la somministrazione, mentre quella degli uomini etero non mostrava alcun cambiamento.
Il giro del cunate, il cingolo laterale anteriore e il giro ippocampo bilaterale/parahippocampale degli uomini etero mostravano una maggiore attività, mentre ne veniva osservata una ridotta in porzioni della loro corteccia cingolata anteriore[70] . Questi risultati suggeriscono che gli omosessuali e gli eterosessuali possono non solo differire nel numero totale di neuroni in varie aree del loro sistema nervoso centrale, ma possono anche differire nella distribuzione di certe loro tipologie, come i neuroni serotoninergici e dopaminergici [69] .
Studi relativi
Vari modelli di animali e insetti sono stati utilizzati per esplorare l' orientamento sessuale e le caratteristiche del cervello.
Un esperimento ha coinvolto l'alterazione genetica della Drosophila maschile, inducendola ad assumere strutture cerebrali femminilizzate coinvolte nell'elaborazione del feromone a contatto sessuale dimorfico. I maschi trasformati mostrarono un aumento dei comportamenti di corteggiamento omosessuale nei confronti di mosche maschi di tipo selvatico; fu cioè trovata una correlazione tra il comportamento di corteggiamento e l'espressione del gene alterato nelle regioni del cervello sessualmente correlate [71] .
Studi futuri
Lo sviluppo dell'orientamento sessuale è un argomento tutt'altro che completo. Mentre le neuroscienze hanno compiuto notevoli progressi nel far luce sui meccanismi e le relazioni tra il cervello umano e l'orientamento, è necessario in ogni caso condurre ulteriori ricerche.
Le aree per la ricerca futura includono [3] :
- trovare marcatori per i livelli di steroidi sessuali nel cervello dei feti che mettono in risalto le caratteristiche del precoce sviluppo neurale che porta a determinati orientamenti sessuali;
- determinare la precisa direzione dei circuiti neurali alla base delle preferenze sessuali;
- utilizzare modelli animali per esplorare i fattori genetici e di sviluppo che influenzano l'orientamento sessuale;
- ulteriori studi sulla popolazione, studi genetici e marcatori sierologici per giungere a chiarire e determinare definitivamente l'effetto del sistema immunitario materno;
- studi di neuroimaging per quantificare le differenze relative all'orientamento sessuale nella struttura e nella funzione dei viventi;
- studi neurochimici per indagare il ruolo degli steroidi sessuali sui circuiti neurali coinvolti nell' attrazione sessuale .
Note
- ^ Sexual orientation, homosexuality and bisexuality , su apa.org , American Psychological Association . URL consultato il 10 agosto 2013 (archiviato dall' url originale l'8 agosto 2013) .
- ^ Sexual Orientation , su healthyminds.org , American Psychiatric Association . URL consultato il 1º gennaio 2013 (archiviato dall' url originale il 22 luglio 2011) .
- ^ a b c d e f g h i j k l m n o p q Q Rahman, The neurodevelopment of human sexual orientation , in Neuroscience & Biobehavioral Reviews , vol. 29, n. 7, 2005, pp. 1057-66, DOI :10.1016/j.neubiorev.2005.03.002 , PMID 16143171 .
- ^ Williams TJ, Pepitone ME, Christensen SE, Cooke BM, Huberman AD, Breedlove NJ, Breedlove TJ, Jordan CL e Breedlove SM, Finger-length ratios and sexual orientation ( PDF ), in Nature , vol. 404, n. 6777, Mar 2000, pp. 455-6, DOI : 10.1038/35006555 , PMID 10761903 . URL consultato il 17 agosto 2016 .
- ^ Swaab DF, Sexual differentiation of the human brain: relevance for gender identity, transsexualism and sexual orientation , in Gynecological Endocrinology , vol. 19, n. 6, dicembre 2004, pp. 301-12, DOI : 10.1080/09513590400018231 , PMID 15724806 .
- ^ Brown et al. 2002
- ^ Okten et al. 2002
- ^ Berenbaum SA, Bryk KK, Nowak N, Quigley CA, Moffat S,Fingers as a marker of prenatal androgen exposure , in Endocrinology , vol. 150, n. 11, novembre 2009, pp. 5119-24, DOI : 10.1210/en.2009-0774 , PMC 2775980 , PMID 19819951 .
- ^ John T. Manning, Peter E. Bundred, Darren J. Newton e Brian F. Flanagan, The second to fourth digit ratio and variation in the androgen receptor gene , in Evolution and Human Behavior , vol. 24, n. 6, 2003, pp. 399-405, DOI : 10.1016/S1090-5138(03)00052-7 .
- ^ TJ Williams, ME Pepitone, SE Christensen, BM Cooke, AD Huberman, NJ Breedlove, TJ Breedlove, CL Jordan e SM Breedlove, Finger-length ratios and sexual orientation ( PDF ), in Nature , vol. 404, n. 6777, marzo 2000, pp. 455-456, DOI : 10.1038/35006555 , PMID 10761903 .
- ^ Tortorice, JL, Written on the body: butch/femme lesbian gender identity and biological correlates , in Rutgers Ph.D. Dissertation , 2002.
- ^ McFadden D, Shubel E, Relative lengths of fingers and toes in human males and females , in Hormones and Behavior , vol. 42, n. 4, dicembre 2002, pp. 492-500, DOI : 10.1006/hbeh.2002.1833 , PMID 12488115 .
- ^ Hall LS, Love CT, Finger-length ratios in female monozygotic twins discordant for sexual orientation , in Archives of Sexual Behavior , vol. 32, n. 1, febbraio 2003, pp. 23-8, DOI : 10.1023/A:1021837211630 , PMID 12597269 .
- ^ a b c Rahman Q e Wilson GD, Sexual orientation and the 2nd to 4th finger length ratio: evidence for organising effects of sex hormones or developmental instability? , in Psychoneuroendocrinology. , vol. 28, n. 3, Elsevier Inc., Apr 2003, pp. 288-303, DOI : 10.1016/S0306-4530(02)00022-7 , PMID 12573297 .
- ^ Csathó A, Osváth A, Bicsák E, Karádi K, Manning J, Kállai J, Sex role identity related to the ratio of second to fourth digit length in women , in Biological Psychology , vol. 62, n. 2, febbraio 2003, pp. 147-56, DOI : 10.1016/S0301-0511(02)00127-8 , PMID 12581689 .
- ^ D Putz, Steven JC Gaulin, Robert J. Sporter e Donald H. McBurney, Sex hormones and finger lengthWhat does 2D:4D indicate? ( PDF ), in Evolution and Human Behavior , vol. 25, n. 3, 2004, pp. 182-199, DOI : 10.1016/j.evolhumbehav.2004.03.005 (archiviato dall' url originale il 7 gennaio 2010) .
- ^ Rahman Q, Fluctuating asymmetry, second to fourth finger length ratios and human sexual orientation , in Psychoneuroendocrinology , vol. 30, n. 4, maggio 2005, pp. 382-91, DOI : 10.1016/j.psyneuen.2004.10.006 , PMID 15694118 .
- ^ Kraemer B, Noll T, Delsignore A, Milos G, Schnyder U, Hepp U, Finger length ratio (2D:4D) and dimensions of sexual orientation , in Neuropsychobiology , vol. 53, n. 4, 2006, pp. 210-4, DOI : 10.1159/000094730 , PMID 16874008 .
- ^ Wallien MS, Zucker KJ, Steensma TD, Cohen-Kettenis PT, 2D:4D finger-length ratios in children and adults with gender identity disorder , in Hormones and Behavior , vol. 54, n. 3, agosto 2008, pp. 450-4, DOI : 10.1016/j.yhbeh.2008.05.002 , PMID 18585715 .
- ^ Bogaert AF,Biological versus nonbiological older brothers and men's sexual orientation , in Proceedings of the National Academy of Sciences of the United States of America , vol. 103, n. 28, luglio 2006, pp. 10771-4, DOI : 10.1073/pnas.0511152103 , PMC 1502306 , PMID 16807297 .
- ^ a b c d e f g Bogaert AF e Skorska M, Sexual orientation, fraternal birth order, and the maternal immune hypothesis: a review , in Front Neuroendocrinol , vol. 32, n. 2, 2011, pp. 247-54, DOI : 10.1016/j.yfrne.2011.02.004 , PMID 21315103 .
- ^ Cantor JM, Blanchard R, Paterson AD, Bogaert AF, How many gay men owe their sexual orientation to fraternal birth order? , in Arch Sex Behav , vol. 31, n. 1, febbraio 2002, pp. 63-71, DOI : 10.1023/A:1014031201935 , PMID 11910793 .
- ^ Blanchard R, Bogaert AF, Homosexuality in men and number of older brothers , in Am J Psychiatry , vol. 153, n. 1, gennaio 1996, pp. 27-31, DOI : 10.1176/ajp.153.1.27 , PMID 8540587 .
- ^ Blanchard R e Bogaert AF, Proportion of homosexual men who owe their sexual orientation to fraternal birth order: An estimate based on two national probability samples. , in Am J Hum Biol. , vol. 16, n. 2, 2004, pp. 151-7, DOI : 10.1002/ajhb.20006 , PMID 14994314 .
- ^ a b Holder, MK What does Handedness have to do with Brain Lateralization (and who cares?) . Retrieved 11 August 2012.
- ^ a b Blanchard R, Lippa RA, Birth order, sibling sex ratio, handedness, and sexual orientation of male and female participants in a BBC internet research project , in Archives of Sexual Behavior , vol. 36, n. 2, aprile 2007, pp. 163-76, DOI : 10.1007/s10508-006-9159-7 , PMID 17345165 .
- ^ dominant , su Merriam-Webster Dictionary , Merriam-Webster. URL consultato l'8 marzo 2017 .
« 4 : biology : being the one of a pair of bodily structures that is the more effective or predominant in action • dominant eye • used her dominant hand» . - ^ non- , su Merriam-Webster Dictionary , Merriam-Webster. URL consultato l'8 marzo 2017 .
«Definition of non- 1 : not : other than : reverse of : absence of • non toxic • non linear» . . ("Nondominant" is one of 945 words listed under "non-") - ^ M Papadatou-Pastou, Martin, M, Munafò, MR e Jones, GV, Sex differences in left-handedness: a meta-analysis of 144 studies , in Psychological Bulletin , vol. 134, n. 5, settembre 2008, pp. 677-99, DOI : 10.1037/a0012814 , PMID 18729568 .
- ^ a b c d e f g h Rahman Q e Wilson GD, Born gay? The psychobiology of human sexual orientation , in Personality and Individual Differences , vol. 34, Elsevier, 2003, pp. 1337-1382, DOI : 10.1016/s0191-8869(02)00140-x .
- ^ a b c Lalumière ML, Blanchard R, Zucker KJ, Sexual orientation and handedness in men and women: a meta-analysis , in Psychological Bulletin , vol. 126, n. 4, luglio 2000, pp. 575-92, DOI : 10.1037/0033-2909.126.4.575 , PMID 10900997 .
- ^ Rahman Q, Fluctuating asymmetry, 2nd to 4th finger length ratios and human sexual orientation. , in Psychoneuroendocrinology. , vol. 30, n. 4, Elsevier Inc., maggio 2005, pp. 382-91, DOI : 10.1016/j.psyneuen.2004.10.006 , PMID 15694118 .
- ^ Mustanski BS, Bailey JM e Kaspar S, Dermatoglyphics, handedness, sex, and sexual orientation. , in Arch. Sex. Behav. , vol. 31, n. 1, Feb 2002, pp. 113-22, DOI : 10.1023/A:1014039403752 , PMID 11910784 .
- ^ a b c Simon LeVay, Gay, Straight, and the Reason Why: The Science of Sexual Orientation. , Oxford University Press, 2010, pp. 230-5, ISBN 0-19-975296-6 .
- ^ Martin JT, Puts DA e Breedlove SM, Hand Asymmetry in Heterosexual and Homosexual Men and Women: Relationship to 2D:4D Digit Ratios and Other Sexually Dimorphic Anatomical Traits , in Arch Sex Behav , vol. 37, Mar 2008, pp. 119-132, DOI : 10.1007/s10508-007-9279-8 , PMID 18161017 .
- ^ Innala SM e Ernulf KE, When gay is pretty: physical attractiveness and low homophobia , in Psychol. Rep. , vol. 74, 3 Pt 1, Jun 1994, pp. 827-31, PMID 8058859 .
- ^ Minna Lyons, Detection of Sexual Orientation ( Gaydar ) by Homosexual and Heterosexual Women , in Arch Sex Behav , vol. 43, 2014, pp. 345-352, DOI : 10.1007/s10508-013-0144-7 .
- ^ Zucker KJ, Wild J, Bradley SJ e Lowry CB, Physical attractiveness of boys with gender identity disorder , in Arch. Sex. Behav. , vol. 22, n. 1, Kluwer Academic Publishers-Plenum Publishers, Feb 1993, pp. 23-36, DOI : 10.1007/BF01552910 , ISSN 1573-2800 , PMID 8435037 .
- ^ Rhodes G, Hickford C e Jeffery L, Sex-typicality and attractiveness: are supermale and superfemale faces super-attractive? , in Br J Psychol. , vol. 91, n. 1, Feb 2000, pp. 125-40, DOI : 10.1348/000712600161718 , PMID 10717775 .
- ^ Thornhill R e Gangestad SW, Facial attractiveness , in Trends Cogn Sci , vol. 3, n. 12, Dec 1999, pp. 452-460, DOI : 10.1016/S1364-6613(99)01403-5 , PMID 10562724 .
- ^ Gangestad SW, Bailey JM e Martin NG, Taxometric analyses of sexual orientation and gender identity , in J. Pers. Soc. Psychol. , vol. 78, n. 6, Jun 2000, pp. 1109-21, DOI : 10.1037/0022-3514.78.6.1109 , PMID 10870912 .
- ^ Richard S Snell, Clinical Neuroanatomy , 7th, Lippincott Williams & Wilkins, 2010, p. 34, ISBN 978-0-7817-5993-9 .
- ^ Hal Blumenfeld, Neuroanatomy through clinical cases , 2nd, Sunderland, Mass., Sinauer Associates, 2010, p. 21, ISBN 978-0-87893-613-7 .
- ^ a b c d e f Swaab DF, Hofman MA, Sexual differentiation of the human hypothalamus in relation to gender and sexual orientation , in Trends in Neurosciences , vol. 18, n. 6, giugno 1995, pp. 264-70, DOI : 10.1016/0166-2236(95)80007-O , PMID 7571001 .
- ^ LeVay S, A difference in hypothalamic structure between heterosexual and homosexual men , in Science , vol. 253, n. 5023, agosto 1991, pp. 1034-7, DOI : 10.1126/science.1887219 , PMID 1887219 .
- ^ a b Byne W, Lasco MS, Kemether E, Shinwari A, Edgar MA, Morgello S, Jones LB e Tobet S., The interstitial nuclei of the human anterior hypothalamus: an investigation of sexual variation in volume and cell size, number and density , in Brain Res , vol. 856, n. 1-2, 21 febbraio 2000, pp. 254-8, DOI : 10.1016/S0006-8993(99)02458-0 , ISSN 0006-8993 , PMID 10677635 .
- ^ Swaab DF e Hofman MA., An enlarged suprachiasmatic nucleus in homosexual men. , in Brain Res. , vol. 537, n. 1-2, 24 dicembre 1990, pp. 141-8, DOI : 10.1016/0006-8993(90)90350-K , PMID 2085769 .
- ^ a b c d e f Abe C, Johansson E, Allzen E e Savic I,Sexual Orientation Related Differences in Cortical Thickness in Male Individuals , in Antonella Gasbarri (a cura di), PLoS ONE , vol. 9, n. 12, Dec 2014, p. e114721, DOI : 10.1371/journal.pone.0114721 , PMC 4257718 , PMID 25479554 .
- ^ Hu S, Xu D, Peterson B, Wang Q e He X,Association of cerebral networks in resting state with sexual preference of homosexual men: a study of regional homogeneity and functional connectivity , in PLOS ONE , vol. 8, 2013, p. e59426, DOI : 10.1371/journal.pone.0059426 , PMC 3605412 , PMID 23555670 .
- ^ Aste N, Balthazart J, Absil P, Grossmann R, Mülhbauer E, Viglietti-Panzica C e Panzica GC, <141::AID-CNE1>3.0.CO;2-0 Anatomical and neurochemical definition of the nucleus of the stria terminalis in Japanese quail (Coturnix japonica). , in J Comp Neurol , vol. 396, n. 2, 29 giugno 1998, pp. 141-57, DOI : 10.1002/(SICI)1096-9861(19980629)396:2<141::AID-CNE1>3.0.CO;2-0 , PMID 9634138 .
- ^ a b Aldo Poiani, Animal Homosexuality: A Biosocial Perspective , Cambridge University Press, Aug 2010, p. 213, ISBN 978-1-139-49038-2 .
- ^ a b Zhou, JN, Hofman, MA, Gooren, LJG e Swaab, DF, A sex difference in the human brain and its relation to transsexuality , in Nature , vol. 378, n. 6552, Nov 1995, pp. 68-70, DOI : 10.1038/378068a0 , PMID 7477289 .
- ^ a b c Savic I, Lindström P,PET and MRI show differences in cerebral asymmetry and functional connectivity between homo- and heterosexual subjects , in Proceedings of the National Academy of Sciences of the United States of America , vol. 105, n. 27, luglio 2008, pp. 9403-8, DOI : 10.1073/pnas.0801566105 , PMC 2453705 , PMID 18559854 .
- ^ a b c Hill AK, Dawood K e Puts DA, Biological Foundations of Sexual Orientation , in Handbook of Psychology and Sexual Orientation , illustrated, OUP USA, 2013, 2012, pp. 55-68, ISBN 978-0-19-976521-8 .
- ^ a b c d Witelson SF, Kigar DL, Scamvougeras A, Kideckel DM, Buck B, Stanchev PL, Bronskill M e Black S, Corpus callosum anatomy in right-handed homosexual and heterosexual men , in Arch. Sex. Behav. , vol. 37, n. 6, Dec 2008, pp. 857-63, DOI : 10.1007/s10508-007-9276-y , ISSN 1573-2800 , PMID 17975723 .
- ^ a b Aldo Poiani, Animal Homosexuality: A Biosocial Perspective , Cambridge University Press, Aug 2010, pp. 213-5, ISBN 978-1-139-49038-2 .
- ^ Simon LeVay, Gay, Straight, and the Reason Why: The Science of Sexual Orientation , Oxford University Press, 2010, 2010, pp. 201-5, ISBN 0-19-975296-6 .
- ^ a b c d e f Ponseti J, Siebner HR, Klöppel S, Wolff S, Granert O, Jansen O, Mehdorn HM e Bosinski HA,Homosexual Women Have Less Grey Matter in Perirhinal Cortex than Heterosexual Women , in PLoS ONE , vol. 2, n. 8, 22 Aug 2007, p. e762, DOI : 10.1371/journal.pone.0000762 , PMC 1942120 , PMID 17712410 .
- ^ Aldo Poiani, Animal Homosexuality: A Biosocial Perspective , Cambridge University Press, Aug 2010, p. 220, ISBN 978-1-139-49038-2 .
- ^ Guillamon A, Junque C e Gómez-Gil E,A Review of the Status of Brain Structure Research in Transsexualism , in Arch. Sex. Behav. , vol. 45, n. 7, Springer US, Oct 2016, pp. 1615-1648, DOI : 10.1007/s10508-016-0768-5 , ISSN 1573-2800 , PMC 4987404 , PMID 27255307 .
- ^ a b Berglund H, Lindström P, Savic I,Brain response to putative pheromones in lesbian women , in Proceedings of the National Academy of Sciences of the United States of America , vol. 103, n. 21, maggio 2006, pp. 8269-74, DOI : 10.1073/pnas.0600331103 , PMC 1570103 , PMID 16705035 .
- ^ a b Savic I, Berglund H, Lindström P,Brain response to putative pheromones in homosexual men , in Proceedings of the National Academy of Sciences of the United States of America , vol. 102, n. 20, maggio 2005, pp. 7356-61, DOI : 10.1073/pnas.0407998102 , PMC 1129091 , PMID 15883379 .
- ^ a b Safron A, Barch B, Bailey JM, Gitelman DR, Parrish TB e Reber PJ, Neural correlates of sexual arousal in homosexual and heterosexual men. , in Behav Neurosci. , vol. 121, n. 2, Apr 2007, pp. 237-48, DOI : 10.1037/0735-7044.121.2.237 , PMID 17469913 .
- ^ a b Paul T, Schiffer B, Zwarg T, Tillmann HC Krüger, Sherif Karama, Manfred Schedlowski, Michael Forsting e Elke R. Gizewski, Brain response to visual sexual stimuli in heterosexual and homosexual males , in Human Brain Mapping , vol. 29, n. 6, giugno 2008, pp. 726-35, DOI : 10.1002/hbm.20435 , PMID 17636559 .
- ^ a b Hu SH, Wei N e Wang QD, Patterns of brain activation during visually evoked sexual arousal differ between homosexual and heterosexual men. , in AJNR Am J Neuroradiol. , vol. 29, n. 10, Nov 2008, pp. 1890-6, DOI : 10.3174/ajnr.A1260 , PMID 18768725 .
- ^ a b Ponseti J, Bosinski HA, Wolff S, Peller M, Jansen O, Mehdorn HM, Büchel C e Siebner HR, A functional endophenotype for sexual orientation in humans. , in NeuroImage , vol. 33, n. 3, 15 novembre 2006, pp. 825-33, DOI : 10.1016/j.neuroimage.2006.08.002 , PMID 16979350 .
- ^ a b c Kranz F e Ishai A, Face Perception Is Modulated by Sexual Preference , in Current Biology , vol. 16, n. 1, 2006, pp. 63-68, DOI : 10.1016/j.cub.2005.10.070 .
- ^ a b c d Sanders G, Wright M, Sexual orientation differences in cerebral asymmetry and in the performance of sexually dimorphic cognitive and motor tasks , in Archives of Sexual Behavior , vol. 26, n. 5, ottobre 1997, pp. 463-80, DOI : 10.1023/A:1024551704723 , PMID 9343633 .
- ^ a b c Aldo Poiani, Animal Homosexuality: A Biosocial Perspective , illustrated, Cambridge University Press, Aug 2010, pp. 222-4, ISBN 978-1-139-49038-2 .
- ^ a b Kinnunen LH, Moltz H, Metz J e Cooper M, Differential brain activation in exclusively homosexual and heterosexual men produced by the selective serotonin reuptake inhibitor, fluoxetine , in Brain Res , vol. 1024, n. 1-2, 22 ottobre 2004, pp. 251-4, DOI : 10.1016/j.brainres.2004.07.070 , PMID 15451388 .
- ^ Ferveur JF, Störtkuhl KF, Stocker RF, Greenspan RJ, Genetic feminization of brain structures and changed sexual orientation in male Drosophila , in Science , vol. 267, n. 5199, febbraio 1995, pp. 902-5, DOI : 10.1126/science.7846534 , PMID 7846534 .