Diptera

Dipterele ( Diptera Linnaeus , 1758 ) constituie un ordin al insectelor terestre sau hidrofile, care se scufundă frecvent sau sunt acvatice în etapele pre-imaginale. Sub aspectul sistematic și filogenetice, Diptera , fac parte din Panorpoidea , prima dintre cele trei linii evolutive în care holometaboly a Endopterygotes dezvoltate . Numele se referă la numărul de aripi cu care sunt prevăzute aceste insecte și derivă din greaca veche δις (dis): de două ori și πτερόν (pteron): aripă.
Probabil ordinea, care include aproximativ 120.000 de specii cunoscute, în formele sale primitive a fost prezentă pe Pământ încă din Permian , dar cu dovezi neechivoce din Triasic . Varietatea remarcabilă a mediilor colonizate și relația frecventă cu omul , cu implicații de natură economică, medicală și igienico-sanitară, fac din această ordine una dintre cele mai importante grupări de insecte: ordinul include specii care se încadrează în cele mai importante plăgi care afectează agricultura mondială, specii responsabile de transmiterea unor boli umane grave și, din păcate, celebre, răspândite uneori pe mai multe continente, cum ar fi malaria , și specii omniprezente și comensale ale omului, cum ar fi camerista cu muște foarte obișnuită.
Morfologie
Marea ordine a Dipterelor, căreia îi aparțin muștele, muscații și țânțarii, se caracterizează prin prezența unei singure perechi de aripi, cele din față. Aripile posterioare ale acestor insecte au fost reduse și transformate în balansoare, două buturugi care garantează stabilitatea zborului. Multe specii au larve acvatice care trec prin ciclurile lor de viață în lacuri, râuri și iazuri. Rolul Dipterelor în conservarea ecosistemelor, la fel ca majoritatea formelor de viață din primele verigi ale lanțurilor alimentare, este foarte important: dacă muștele și țânțarii ar dispărea ipotetic, unele specii de păsări, reptile sau amfibieni incapabili să se adapteze ar risca dispariția.
Adult
Adulții sunt mici sau mijlocii, în timp ce cele mari sunt rare: numai anumite diptere din Mydidae și Pantophthalmidae familiile ajunge la anvergura aripilor 95-100 mm. Livrele au culori modeste sau strălucitoare, uniforme sau pestrițe; uneori apare aposematismul , ca de exemplu la Syrphidae . Exoscheletul nu este de obicei foarte consistent. O importanță fundamentală în scopuri taxonomice este prezența și distribuția apendicelor tegumentale, în special a perilor.
 | Reprezentarea schematică a morfologiei unui dipter muscoid. I : cap; II : piept; III : abdomen. 1 : prescripție; 2 : stigmatul anterior; 3 : scuto; 4 : bazal; 5 : caliptra; 6 : scutellum; 7 : coasta aripii (coasta); 8 : aripă; 9 : urită; 10 : bara; 11 : stigmatul posterior; 12 : femur; 13 : tibia; 14 : pinten; 15 : tars; 16 : propleura; 17 : prosterno; 18 : mezopleura; 19 : mesosterno; 20 : metapleura; 21 : metasterno; 22 : ochi compus; 23 : arista; 24 : antena; 25 : palpi maxilari; 26 : buza inferioară; 27 : labellum; 28 : pseudotrahee. |
Șefu
| Reprezentarea schematică a morfologiei capului unui dipter muzcoid | |
 | 1 : labellum; 2 : buza inferioară (labium); 3 : palp maxilar; 4 : buza superioară (labrum); 5 : zona subgenală; 6 : clipe; 7 : zona fronto-orbitală; 8 : peri fronto-orbitali; 9 : peri verticali externi; 10 : peri verticali interni; 11 : peri postcelari; 12 : ocelli; 13 : peri ocelari; 14 : ochi compus; 15 : sutura frontală sau sutura ptilinală; 16 : antena; 17 : arista; 18 : mustăți. |
Capul este bine distinct de torace , cu o îngustare marcată la nivelul gâtului. În formele primitive este prognozat în celelalte este de obicei hipognat . Conformația capsulei craniene variază substanțial, trecând de la Nematocera primitivă la Diptera mai evoluată. În primul, partea dorsal-ventrală a craniului se extinde înainte, în ceea ce privește ochii datorită dezvoltării în lungime a clipeului și a gâtului (numită și zona subgenală ); aparatul bucal este prezent la capătul distal al acestei extensii. În celelalte diptere capul are o formă subgloboză, iar regiunea fronto-clipeală are o dezvoltare limitată prezentându-se ca o zonă delimitată deasupra de ochi și de vârf (sau de ochi numai în formele cu cap holoptic, lateral de ochi și de genă sau obraji și inferior din părțile bucale.În Cyclorrhapha Schizophora , un element morfologic de o importanță deosebită este prezența suturii ptilinale sau a suturii frontale , formată prin reabsorbția ptilinului după fluturare. ruptura pupalei . plic ( puparium ) prin umflarea unei vezicule frontale, ptilinul . După pâlpâire, vezicula este reabsorbită în capsula craniană și o sutură rămâne în regiunea frontală care are o formă de U inversată. sutura separă clar două regiuni : cea superioară este regiunea frontală , care are continuitate cu vârful, orbi asemenea și genele; cea inferioară se numește față sau clipe , conține inserția antenelor și se termină inferior cu marginea epistomală, în corespondență cu care se articulează buza superioară .

Ochii sunt în general bine evidenți, dar ating o dezvoltare notabilă în generalitatea Brachiceri . În acest subordine, acestea sunt marcat convexe și dezvoltate până să ocupe o mare parte a feței laterale. Vârful, între cei doi ochi, poate fi uneori redus la o zonă îngustă care merge de la frunte la regiunea occipitală sau poate dispărea complet datorită contactului direct dintre ochi sau fuziunii lor. Morfologia ochiului compus se caracterizează prin numărul considerabil de omatidii, de ordinul a mii în muscoizi. Ocelii , atunci când sunt prezenți, sunt localizați în vârful vârfului, dispuși în funcție de vârfurile unui triunghi într-o zonă numită stemmaticum sau triunghi ocelar . În scopuri sistematice, prezența, dispunerea și conformarea perilor cefalici sunt importante, cărora li se asociază o terminologie specifică. Părul este împărțit în vibrise , scurte și robuste, și macrochete semnificativ mai lungi . Conform aranjamentului, acestea sunt împărțite în fronto-orbital, vertical, post-vertical, ocelar și postcelar.
Antenele sunt împărțite în două tipuri morfologice de bază, care stau la baza distincției dintre cele două subordine și a denumirii lor. În Nematocera sunt pluriarticulate, de tip filiform sau pene, compuse din 7-15 articole nediferențiate. În Brachicerae sunt compuse din maximum șase articole, dintre care primele trei sunt bine dezvoltate, iar celelalte reduse pentru a forma un apendice filiform, numit arista sau stylus ; peisajul și pedicelul sunt subdezvoltate relativ mici; al treilea antennomer este vizibil mai mare și poartă arista.
Aparatul bucal prezintă, conform grupurilor sistematice, o varietate de conformații atribuibile tipului de supt înțepător (de exemplu Tabanidae și Culicidae ), tipului de supt lambent ( Muscoidea ) și tipului de supt (de exemplu Syrphidae ). Morfologia și funcționalitatea sunt strâns asociate cu regimul alimentar: tipul supt-înțepător apare în sânge -sucking diptere, tipul supt supt în glyciphagous diptere care se hrănesc cu soluții de zahăr cristalizate, tipul supt în glyciphages care , practic , hrana pentru animale pe nectar și polen .
Cufăr
| Reprezentarea schematică a morfologiei toracelui unui dipter muscoid: vedere dorsală (stânga), laterală (dreapta). | |
 |  |
| 1 : pe jumătate deschis; 2 : calus humeral; 3 : notopleuron; 4 : mesoscuto; 5 : calus posterior; 6 : mezoscutellum; 7 : sutură transversală; 8 : postscutello; 9 : metan; 10 : baza aripii; 11 : mezopleuro-tergit sau laterotergit sau katatergit; 12 : bara; 13 : stigmate; 14 : metapleuron; 15 : metacoxa; 16 : hipopleuron sau meron; 17 : mezocoxa; 18 : sutura epimerica; 19 : mezoepimer sau anepimer; 20 : mezoepisternum ventral sau katepisternum; 21 : sutura episterno-precoxală; 22 : procoxa; 23 : sutura pleurală; 24 : mezoepisternum dorsal sau anepisternal; 25 : propleuron. Păr și fire de păr: a : acrostical; dc : dorsal central; ph : posthumeral; om - humeral; ps : presutural; np : notopleural; ia : intralar (postsutural); sa : supralar (postsutural); pa : poștal; psct : scutellari. | |
Particularitatea fundamentală a Dipterelor este remarcabila specializare evolutivă realizată în forma aripilor și în adaptarea morfo-anatomică a toracelui. Cu excepția formelor rare de meiotere (microptere, subacterii sau atteri), Dipterele sunt de obicei înaripate și folosesc aripile ca principal mijloc de locomoție.

Nivelul de specializare anatomică, morfologică și funcțională este de așa natură încât să facă din aceste insecte fluturători formidabili în general, cu o referire specială la agilitate. Toate Dipterele sunt prevăzute cu o singură pereche de aripi funcționale, cele mezotoracice (anterioare). Aripile metatoracice sunt în schimb transformate în basculante . Din acest personaj derivă numele ordinului, din grecescul " dipteros " care înseamnă "cu două aripi". Ca o consecință a acestei structuri morfologice, mezotoraxul reprezintă segmentul cu cea mai mare dezvoltare și complexitate, în timp ce protoraxul și metatoraxul sunt reduse în mod hotărât.
Echilibratoarele sunt organe claviforme utile pentru echilibrarea zborului, constând dintr-o porțiune proximală dilatată conectată la percepția auditivă (printr-o membrană cordotonală) întinsă către o parte distală bogată în peri senzitivi. Omologia dintre aripile posterioare și basculante este demonstrată de mutantul cu patru aripi al muștei Drosophila melanogaster , care este lipsit de ele. Dezvoltarea bilelor variază în funcție de grupul sistematic: la Tipulidae sunt subțiri, dar lungi și clar vizibile, dar în general sunt ascunse de aripi în majoritatea celorlalte grupuri. În Calyptratae , o grupare care include cele mai evoluate diptere, bilele sunt protejate de caliptra inferioară (sau scara toracică ), una dintre cele două scale care formează caliptra .

Aripa mezotoracică este în întregime membrană, complet transparentă și incoloră, sau are pigmentări zonale utile în scopuri de recunoaștere. Suprafața sa este împărțită în trei regiuni: cea mai dezvoltată este regiunea remiges, inclusiv cele mai robuste coaste; posterior există regiunea anală și, în cele din urmă, în tractul posterior-proximal, există o expansiune lobiformă numită alula . În Calyptratae, între alula și torace se află caliptra superioară, numită și scară de aripă sau tegula . Sistemul nervurilor este simplificat, dar este reprezentativ pentru sistemul Comstock-Needham , care a fost conceput la sfârșitul secolului al XIX-lea tocmai pentru a defini terminologia specifică morfologiei aripilor Dipterelor. În Diptera coastele longitudinale tipice ( coasta , subcosta , raza , mijlocul , cubitus ) și ramificațiile relative sunt, prin urmare, bine evidente și distincte, la care se adaugă două coaste anale, dintre care a doua, numită și axilară , separă regiunea anală de clasă. Diferențierile particulare privesc ramificațiile, coastele transversale și forma celulelor, acestea sunt caractere importante pentru determinarea taxonomică.
Picioarele sunt de tip ambulator, relativ subțiri în comparație cu corpul.
Abdomen

Morfologia abdomenului este substanțial determinată de adaptarea morfo-anatomică, la ambele sexe, ca funcție de reproducere. În general, cele zece urite sunt reduse la un număr mai mic de urite aparente datorită modificărilor structurale ale primei și ultimei urite .
În partea anterioară apare de obicei atrofia primei urite și fuziunea a 2-a și a 3-a urotergite . Tergitele și sternitele se pot distinge clar unele de altele, dar apare adesea o dezvoltare diferențială prin care tergitul acoperă sternita până la partea ventrală a abdomenului; cazul extrem apare atunci când expansiunile tergitei fuzionează ventral formând o structură de tub sau inel.
La femele, ultimii uriti devin mai subtiri si alungiti, formand un aparat telescopic extrovertabil, numit ovipozitor de substitutie . Această adaptare morfologică este adesea însoțită de scleroza structurilor terminale din urita a opta, astfel încât ovipozitorul de înlocuire este capabil să pătrundă prin țesuturile organismului care vor găzdui ouăle și larvele.
La mascul, ultimele urite suferă o transformare complexă pentru a forma un aparat, integrat cu organele genitale externe, numit hipopigiu . Gradul și natura modificării structurale se modifică în funcție de grupul sistematic, dar, în general, se manifestă semnificativ odată cu dezvoltarea lobilor forcepsului al nouălea urotergus ( epandru ) și al nouălea urosterno ( hipandru ). Uneori există o răsucire de-a lungul axei abdomenului, rezultând o inversare a pozițiilor epandrului și hipandrului.
Larvă

Majoritatea larvelor Diptera trăiesc într-un mediu acvatic, în interiorul substraturilor organice în descompunere, în interiorul altor organisme ( ciuperci , animale , plante ). Structura lor morfologică prezintă, prin urmare, o simplificare substanțială potrivită pentru un tip de viață neliberă.
Larva Diptera poate fi urmărită înapoi la tipul apodo , adică cu absența picioarelor, chiar dacă uneori, mai ales în larvele acvatice, există apendicele similare pseudopodelor . Capul este în general lipsit de ochi, are un aparat de gură de mestecat modificat, antene cu până la șase articole, mai mult sau mai puțin dezvoltate sau reduse la papile. Capul se poate distinge clar de torace (larve eucefalice ), indistinct de restul corpului ( microcefalic ), scufundat în torace ( criptocefalic ).
Pe baza numărului și a aranjamentului spiralelor traheale , se disting următoarele tipuri de sistem respirator:
- apneustic , cu absența stigmatelor;
- metapneustic , cu o singură pereche de stigme abdominale;
- amfipneustic , cu o pereche de stigme în protorax și una abdominală;
- holopneustic , cu două perechi de stigme toracice și opt perechi de abdominale.

Cel mai frecvent tip, recurent în majoritatea brachicerelor, este cel amfipneustic, în timp ce celelalte tipuri apar mai ales în larvele acvatice.
Cele mai familiare larve, caracteristice Brachycera Cyclorrhapha , au un aspect vermiform, cu o diferențiere redusă a regiunilor corpului (cap, torace, abdomen) până la punctul în care sunt denumite în mod obișnuit, în mod necorespunzător, viermi ; aceste larve au o formă cilindric-conică, mai largă în tractul abdominal, aparat bucal simplificat, reprezentat de două mandibule în formă de cârlig și o serie de sclerite cefalice interne, care formează așa - numitul sistem cefalo-faringian în ansamblu . Spre deosebire de mandibule ale aparatului gurii de mestecat, cârligele aparatului cefalofaringian sunt echipate cu mișcări conform unui plan vertical.
Adaptări morfologice deosebite pot fi găsite în larvele adaptate pentru a trăi într-un mediu acvatic sau ca endoparasitoizi : de exemplu, larvele acvatice ale Syrphidae ( viermi de coadă de șobolan ) au un sifon respirator lung, care le permite să trăiască scufundate în funduri noroioase sau în apele putride, în timp ce cele ale Tachinidae au conducte respiratorii care se deschid în traheele gazdei sau în afara corpului său.
Pupa

Pupele Diptera pot fi obtecte sau exarate . Primele au schițele aripilor și picioarelor vizibile, dar aproape de restul corpului, întrucât întregul este înfășurat într-o singură cuticulă; acestea din urmă au apendicele înfășurate în propria lor cuticulă și, prin urmare, sunt detașabile de restul.
Pupele de la Brachycera Cyclorrhapha se pot atribui tipului evoic ( pupe coartate), deoarece evoluează într-un pseudobozzol, numit puparium , format printr-o modificare morfologică și biochimică a exuviei din ultimul stadiu larvar. Modul în care pupariul este deschis, în actul pâlpâirii, face distincția între două mari grupuri sistematice, Aschiza și Schizophora . Pupele exarate sunt, de asemenea, evoice și sunt identificate cu cele ale Ciclorrafi. Pe de altă parte, pupae obteche sunt în general libere și neprotejate, cu excepția celor de la Simuliidae , care sunt protejate de coconi construiți cu resturi cimentate de mătase .
În ultima fază a vieții lor, pupele din Diptera devin mobile. Această mobilitate se datorează de fapt adultului sub formă de farată , adică deja complet format, dar care încă nu a scăpat din exuvia pupală.
Biologie

Dipterele se reproduc în general prin amfigonie și sunt ovipare , cu cazuri mai puțin frecvente, dacă nu rare, de reproducere prin partenogeneză sau pedogeneză . Pot apărea și viviparitate și ovoviviparitate . Dezvoltarea post-embrionară este holometabolică sau, în unele cazuri, hipermetabolică . O caracteristică recurentă în rândul Dipterelor este scurtimea remarcabilă a ciclului de dezvoltare, un aspect al unei probleme deosebite în cazul Dipterelor dăunătoare sau nocive, deoarece face dificilă controlul acestora în cazul alternării a numeroase generații.
Larvele, de obicei orbe, trăiesc în cele mai dispar medii: în sol , în apă , în materiale în descompunere, în cuiburile insectelor sociale și în interiorul altor organisme; mai puțin frecventă este viața liberă în aer liber. Printre organismele predominant atacate există în special ciuperci , animale și plante, iar relația trofică cu gazda este configurată, după caz, cu prădare (ciuperci, plante), cu parazitism ( mamifere ), cu parazitoidism (alte insecte). Larvele micetofage trăiesc în interiorul minelor săpate între branhii în carnea corpurilor fructifere. Acei fitofagi trăiesc, de asemenea, în interiorul minelor, săpate mai ales în frunze, fructe, tulpini și rădăcini cărnoase sau în interiorul galurilor . Larvele zoofage trăiesc mai ales în cavitățile corpului sau într-o poziție subcutanată și provoacă infecții numite miasă . În cele din urmă, larvele entomofage trăiesc în interiorul larvelor altor insecte și, prin urmare, se comportă ca parazitoizi endofagieni .

Metodele de locomoție ale larvelor apod din Ciclorrafi sunt singulare: aceste larve, adaptate pentru a trăi în interiorul cavităților, se mișcă alternând contracții și extensii ale corpului, pivotând pe pereții cavității în care trăiesc, ajutându-se cu cârligele aparatul cefalic.-faringian și cu zone ambulatorii situate în partea ventrală a corpului. Când sunt în afara mediului lor natural, sunt capabili să sară folosind corpul ca arc: pliază corpul prin apucarea capătului caudal cu piesele bucale, după care îl eliberează brusc îndreptând corpul sărind în urma reacției suprafeței a sustine.
Adulții sunt în general fluturași excelenți: datorită structurii anatomice și morfologice complexe a sistemului de zbor (musculatura directă și indirectă, articularea scleritelor aripilor, basculanților, airbag-urilor) aripa lor este capabilă să efectueze mișcări complexe care dau acestor insecte o remarcabilă agilitate și viteză. În general, Dipterele reprezintă, împreună cu himenopterele , cea mai înaltă expresie evolutivă a funcționalității zborului la insecte, până la punctul în care adulții din diferite specii își petrec o parte considerabilă din viață planând în zbor. Nu mai puțin sofisticată este funcționalitatea pretarsi, ale cărei anexe (cuie, empodiu, arolium) permit parcarea și locomoția pe suprafețe de diferite tipuri, inclusiv pe cele extrem de netede, cum ar fi sticla, chiar și pe poziție inversată.

Dietele adulților, în general, pot fi urmărite înapoi la o dietă lichidă: indiferent de eterogenitatea morfologică, piesele bucale sunt adaptate pentru a lua numai lichide, după perforație sau, mai frecvent, prezente pe substratul alimentar. Aportul are loc prin aspirație în diptere cu piese bucale supt sau piercing-supt sau prin ascensiune capilară , la cei cu piese bucale supt-supt . Printre reapar Diptera adulti frecvent in dieta regim hematofagi , reprezentat atât între nematocere ( de ex. Culicidae ), atât între Brachiceri (ex. Tabanids , Ippoboscidi , Glossinidi , Muscidae Stomoxidini ). Cu toate acestea, regimul dietetic glicifag este mai larg reprezentat. În acest caz , sursa de hrana este reprezentata de nectarul florilor, de mană a Rincoti , de sucuri de legume emise de răni, de fluide și substanțe zaharoase de diferite tipuri. În special, structura anatomo-morfologică complexă și funcționalitatea etichetei aparatului de gură care suge gura, prin intermediul așa-numitelor pseudotrahee , permite acestor diptere să ia zaharuri cristalizate, ca în cazul mierii sau a mierii cristalizate sau zahăr obișnuit. Specificitatea dietei glifagice necesită suplimentarea alimentelor cu surse azotate, care sunt frecvent reprezentate de excremente sau materiale organice în descompunere. Acest aspect are o importanță igienico-sanitară deosebită pentru diptere comensale umane, cum ar fi muște comună: această insectă, care se hrănește simultan cu substraturi nesănătoase și cu alimente destinate oamenilor, este un posibil vector al agenților patogeni transmiși prin alimente.
Importanţă
După cum sa menționat la început, ordinea are o importanță considerabilă în multe domenii, de la științific la medical, de la alimentație la agricultură, de la ecologic la biotehnologie.
Domeniul medical și igienico-sanitar
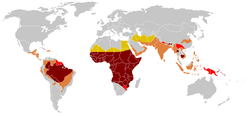 | |
| Harta statistică a riscului de contracție a malariei într-o ședere de o lună | |
| ♦ | Risc ridicat |
| ♦ | Risc mediu |
| ♦ | Risc scazut |
| ♦ | Risc aproape zero |
| ♦ | Absența malariei |
Interesul medical, atât în domeniul uman, cât și în cel veterinar, implică în mod direct dipterii sugeți de sânge și, mai general, zooparaziții vertebratelor . Nocivitatea acestor insecte se manifestă în trei aspecte diferite:
- iritații ale pielii, alergii și alte patologii directe cauzate de activitatea de hrănire a adulților care suge sângele. Activitatea trofică a femelelor Culicidae (țânțari), a Psicodidae ( muște de nisip ) și a altor nematocere se încadrează în acest context. Ocazional, cel puțin în zonele temperate, este activitatea trofică asupra oamenilor de Brachycera care suge sânge, care este în general de interes veterinar. Brachicerele care suge sânge aparțin diferitelor familii, cum ar fi Tabanidele ( muștele calului ), Glossinidele , Hipoboscidele și tribul Stomoxydini , aparținând familiei Muscidi ;
- mioza cauzată de activitatea trofică a larvelor parazitare sau a sarcofagelor. Larvele diferitelor brahicere se dezvoltă ca paraziți sub pielea vertebratelor sau în detrimentul mucoasei cavităților interne sau pur și simplu pe rănile cauzate de traume de diferite tipuri, provocând patologii denumite în mod generic miiază . Dipterele responsabile de miasă aparțin în majoritate familiilor de muscoizi incluși în superfamilia Oestroidea . Interesul acestor insecte este în principal veterinar, dar în regiunile subdezvoltate se extinde și în domeniul medicinei umane. Agenții ocazionali de miasă care afectează membranele mucoase ale sistemului digestiv , pot fi, de asemenea, larvele de diptere neparazite care sunt ingerate accidental de oameni cu ingestia de alimente stricate. Cazuri de acest gen sunt citate în literatură, de exemplu, cu consumul de brânzeturi care conțin larve Piophila casei , care pot provoca boli gastro-intestinale sub formă de colici [1] ;

- transmiterea agenților patogeni infecțioși cauzată de suge de sânge la adulți. Insectele care suge sângele sunt adesea vectori de microorganisme patogene, în special virusuri și protozoare , transmise oamenilor sau animalelor prin injectarea de salivă infectată. Transmiterea poate fi ocazională sau poate presupune proporții de epidemii reale, în unele cazuri extinse la regiuni întregi. În domeniul medicinei umane, unele plasmodiaze sunt, din păcate, renumite, cum ar fi malaria , transmisă de anofeli , și tripanosomiaza africană , mai bine cunoscută sub numele de boală a somnului , transmisă de diferite specii de Glossina , denumite în mod obișnuit muștele tsetse . În domeniul veterinar, febra catarală de origine virală, mai bine cunoscut sub numele de boala limbii albastre, transmise de Culicoides imicola , un nematocere al ceratopogonide familiei, este de o importanță deosebită în Italia .
Interesul igienico-sanitar constă în etologia diferiților muscoizi comensali umani, printre care musca de casă are un rol principal. Această insectă alternează frecventarea unor medii și substraturi nesănătoase (deșeuri, excremente, spații în condiții igienice precare etc.) cu prezența în medii casnice, odihnindu-se pe mâncare, pe vase, pe oameni. Simplul contact sau, mai probabil, deversarea regurgitării și excrementelor este cauza transmiterilor accidentale de microorganisme patogene care provoacă febră sau afecțiuni gastro-intestinale sau către membranele mucoase interne sau externe. Musca este, de asemenea, un posibil vector al agenților tifosului și holerei [2] .
În cele din urmă, merită menționat stresul cauzat, atât de diptere comensale, cât și de sugeți de sânge, la oameni și animale domestice.
Domeniul agricol

Nu mai puțin important este interesul agrar legat de unele grupări sistematice de Diptera. Dipterele fitofage, dăunătoare în stadiul larvelor, sunt prezente în diferite familii, dar în cea mai mare parte aparțin în principal celor din Tephritidae , Agromyzidae și Anthomyiidae , dintre Brachycerae și cele ale Cecidomyiidae din Nematocerae. Conform biologiei larvei, daunele pot fi urmărite în trei tipuri diferite:
- Formarea galelor sau deformări datorate hipertrofiei sau atrofiei cauzate de dezvoltarea larvei endofite. Questo comportamento si riscontra nella generalità dei Cecidomiidi fitofagi e in molti Tefritidi.
- Formazione di mine all'interno dei frutti, a cui segue spesso l'insediamento di microrganismi patogeni. Questo comportamento si riscontra in molti Tefritidi, genericamente chiamati mosche della frutta .
- Formazione di mine all'interno della foglia, con un'estensione che può coinvolgere l'intero lembo fogliare o buona parte di esso, pregiudicando la funzionalità dell'apparato assimilante o causando, nella migliore delle ipotesi, danni di tipo estetico negli ortaggi e nelle piante ornamentali. Questo comportamento si riscontra per lo più negli Agromizidi e negli Antomiidi.

Le specie di maggiore importanza economica rientrano nella famiglia dei Tefritidi e causano danni di grande entità sulle produzioni frutticole su estese regioni, arrivando anche alla perdita dell'intero raccolto e rappresentano autentici flagelli per la loro polifagia, l'aggressività sulle colture e la difficoltà di controllo con metodi di lotta convenzionali. Tali sono ad esempio la Ceratitis capitata (mosca mediterranea della frutta) o la Bactrocera dorsalis (mosca orientale della frutta), quest'ultima presente attualmente solo nel Sud-est asiatico e in Nordamerica . Nella regione mediterranea sono particolarmente temibili la Bactrocera oleae (mosca delle olive) e la Rhagoletis cerasi (mosca delle ciliegie), entrambe considerate i fitofagi più dannosi, rispettivamente, dell' olivo e del ciliegio . Ancora più temibile, nella regione mediterranea, è la Ceratitis capitata , che colpisce gran parte delle produzioni frutticole estive e autunnali nelle stazioni più calde.
Cecidomiidi, Agromizidi e Antomiidi sono statisticamente meno dannosi, tuttavia diverse specie possono avere una spiccata dannosità su determinate colture di particolare importanza in comprensori meno estesi.
Fra i ditteri dannosi vanno citati anche i micetofagi , come gli Sciaridae ei Mycetophilidae , le cui larve invadono numerose i corpi fruttiferi dei funghi commestibili sia spontanei sia coltivati. Gli attacchi in genere non pregiudicano la riproduzione del fungo, tuttavia quelli di una certa entità rendono completamente inutilizzabile il prodotto.
Ambito alimentare ed ecologico

Le larve di molti ditteri sono fitosaprofaghe o zoosaprofaghe e in un ecosistema naturale si collocano all'inizio della catena alimentare del decompositore. Queste larve si ritrovano comunemente in tutti i substrati organici in via di decomposizione, quali rifiuti, lettiere, acque luride, carogne di animali, escrementi, ecc. La stessa mosca domestica, allo stadio larvale, è un organismo detritivoro che si rinviene nei rifiuti in decomposizione. Sotto l'aspetto ecologico i ditteri saprofagi sono organismi utili in quanto intervengono nelle fasi iniziali dei processi di decomposizione della sostanza organica .
Alcuni saprofagi sono invece considerati dannosi quando la loro attività trofica si indirizza sugli alimenti. In questo ambito si citano, fra gli altri, la Piophila casei (mosca del formaggio), dannosa al formaggio , le Drosophila (moscerini dell'aceto), dannose alla frutta, alcuni Sarcophagidae e alcuni Calliphoridae (detti rispettivamente mosconi grigi e azzurri della carne), dannosi alle carni. Oltre al danno diretto, a carico degli alimenti citati, molti di questi ditteri possono essere considerati dannosi anche sotto l'aspetto igienico-sanitario, in quanto possibili responsabili di miasi o vettori di microrganismi patogeni.
Un cenno particolare va fatto alla Piophila casei . Questo dittero, notoriamente responsabile di un'alterazione degenerativa a carico dei formaggi, è sfruttato in contesti locali per una trasformazione guidata che porta alla formazione di una crema piccante. In Sardegna il casu marzu è considerato un prodotto prelibato destinato a un mercato di nicchia, ma prodotti analoghi ottenuti sempre sfruttando la P. casei sono presenti anche in altre regioni in Italia e all'estero, per quanto meno noti del "formaggio marcio" sardo.
Ambito biotecnologico

Alcune famiglie di ditteri sono oggetto di studio e di applicazione come organismi ausiliari, anche su scala commerciale, nel controllo biologico degli insetti fitofagi, sia in lotta biologica sia in lotta integrata . L'attività antagonista dei ditteri entomofagi si svolge in due diversi modi, come predatori e come parassitoidi endofagi, in entrambi i casi a livello di larva.
I ditteri predatori fanno capo alla famiglia dei Syrphidae e sono considerati fra i principali antagonisti degli afidi ; alcune specie sono anche allevate in biofabbrica e commercializzate, ma lo sfruttamento dei Sirfidi si basa per lo più sulle popolazioni indigene, da tutelare con il ricorso a tecniche di conduzione agricola a basso impatto.
Fra gli ausiliari parassitoidi, la famiglia di maggiore importanza è quella dei Tachinidae , le cui larve rientrano fra i principali antagonisti dei Lepidotteri . Altri ditteri parassitoidi, di minore importanza, si annoverano fra le famiglie dei Cecidomyiidae, degli Asilidae , dei Bombyliidae , dei Sarchophagidae .
Nell'ambito delle biotecnologie va infine citata la Drosophila melanogaster , l' organismo modello per eccellenza nel campo della ricerca genetica e in biologica , oggetto di studio da circa un secolo.
Sistematica
Attualmente non esiste una condivisione unanime sulla sistematica dei Ditteri. La tassonomia tradizionale dei Ditteri, fondata su base morfologica, suddivideva l'ordine in due sottordini: - Nematocera (77 famiglie, di cui 35 estinte): antenne lunghe, pronoto distinto dal mesonoto; larve eucefaliche o emicefaliche e spesso acquatiche; - Brachycera (141 famiglie, di cui 8 estinte): antenne corte, pupa entro un pupario fatto dall'ultima esuvia larvale; larve generalmente tozze e con parti boccali ridotte.
I vecchi schemi di classificazione suddividevano a loro volta i Brachycera in - Orthorrapha: ditteri con pupa semplice, obtecta, come quelli delle famiglie di Asilidae e Tabanidae; - Cyclorrapha: pupa in un pupario indurito; a loro volta distinti in Aschiza e Schizophora, a seconda della rispettiva assenza o presenza dello ptilinium.
Tuttavia, la monofilia dei Nematoceri non è più riconosciuta ormai da tempo, e questa organizzazione viene talvolta mantenuta informalmente per pura praticità.
Una recente filogenesi (Wiegmann, Brian M., et al.) conferma la monofilia dell'ordine dei Ditteri, del sottordine dei Brachiceri, come pure del gruppo Cyclorrapha ed evidenzia, d'altro canto, la parafilia dei Nematocera, che sono solo in parte sister group dei Brachycera. Gli autori considerano Mecoptera (mosche scorpione) e Siphonaptera (pidocchi) quali ordini più vicini ai Ditteri.
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Poiché i Brachiceri si sono evoluti dai Nematoceri, che son parafiletici, questi ultimi rappresentano la forma più ancestrale di dittero (effettivamente sono comunemente detti “lower Diptera”). Sempre secondo Wiegeman et al. 2011, le famiglie Deuterophlebiidae (unico genere Deuterophlebia) e Nymphomiidae (unico genere Nymphomyia), dovrebbero essersi speciate per prime, contrariamente alla visione tradizionale secondo cui alla base della filogenesi dei Ditteri ci sarebbero stati, seppure con molta incertezza, i Tipulidae oi Tanyeridae (Gullan&Cranston, 2010). I restanti infraordini di Nematoceri sono rappresentati da Tipulomorpha, Culicomorpha, Psychodomorpha e Bibionomorpha, con questi ultimi identificati come sister group dei Brachiceri. Lo stato monofiletico dei Brachiceri è fortemente supportato sia da dati morfologici sia molecolari, e al suo interno si sarebbero differenziati per primi Eremoneura, Cyclorrapha, Schizophora e Calyptratae. Gli “Aschiza” non formano un gruppo sistematicamente valido, in quanto le loro famiglie stabiliscono un insieme parafiletico di cui i Pipunculidi sono la famiglia più vicina agli Schizophora. Per il resto, negli Schizophora si individuano il gruppo parafiletico degli Acaliptrati (al cui interno le relazioni tra famiglie sono molto poco risolte) e quello monofiletico dei Caliptrati.
Gli studi filogenetici hanno rivelato come sister group dei Drosophilidae, le famiglie Braulidae ei Cryptochetidae. Un risultato, questo, molto interessante dacché ritroviamo come parenti più prossimi del nostro organismo modello, insetti la cui biologia, anatomia, ecologia e fisiologia sono l'antifrasi anche del più rilassato tentativo di archetipizzazione. I Braulidi, meglio noti come pidocchi delle api, sono minuscoli, appiattiti dorso-ventralmente e atteri, e trascorrono la loro vita da commensali di api, al cui corpo aderiscono grazie alle zampe straordinariamente modificate e specializzate. Dei Criptochetidi sono particolari invece le larve, endoparassite di Emitteri, in particolare di cocciniglie. Uno dei punti più critici nella ricostruzione della filogenesi dei Ditteri riguarda le relazioni entro il gruppo degli Schizophora, relazioni che si sta tentando di risolvere anche attraverso l'uso di moderni marcatori molecolari come i miRNA (Wiegemann et al., 2011). In generale, la storia evolutiva di quest'ordine di insetti resta ancora contorta e arzigogolata, e sembra che a complicare la situazione contribuisca un trascorso di episodiche esplosioni di rapidissime radiazioni (Wiegemann et al., 2011), con conseguente compressione nel tempo di segnale filogenetico.
La diversificazione tra Imenotteri, Ditteri e Lepidotteri, e la radiazione stessa dei Ditteri, sembrano risalire al Cretaceo inferiore, contemporaneamente o appena dopo la radiazione delle angiosperme (Musaf et al., 2014). Comunque, nuovi interessanti studi da approfondire suggeriscono, ancora timidamente, di rivedere l'idea della diversificazione delle piante a fiore come precedente a quella degli insetti, e dunque anche dei nostri Ditteri, considerando l'eventualità che, al contrario, l'esplosione in diversità dei consumatori fitofagi, fitomizi e glicifaci abbia anticipato quella delle piante, che hanno utilizzato gli insetti quali propri vettori di diversificazione (Clarcke et el., 2011).
Bibliografia:
- Clarke, JT, Warnock, R., & Donoghue, PC (2011). Establishing a time‐scale for plant evolution. New Phytologist, 192(1), 266-301. - Gullan, PJ, & Cranston, PS (2010). Lineamenti di entomologia. Zanichelli.
- Wiegmann, BM, Trautwein, MD, Winkler, IS, Barr, NB, Kim, JW, Lambkin, C., ... & Yeates, DK (2011). Episodic radiations in the fly tree of life. Proceedings of the National Academy of Sciences, 108(14), 5690-5695.
- Misof, B., Liu, S., Meusemann, K., Peters, RS, Donath, A., Mayer, C., ... & Ott, M. (2014). Phylogenomics resolves the timing and pattern of insect evolution. Science, 346(6210), 763-767.
Note
- ^ Servadei et al. , p. 511 .
- ^ Servadei et al. , p. 523 .
Bibliografia
- Antonio Servadei, Sergio Zangheri, Luigi Masutti. Entomologia generale ed applicata . Padova, CEDAM, 1972.
- Ermenegildo Tremblay. Entomologia applicata . Volume I. 3ª ed. Napoli, Liguori Editore, 1985. ISBN 88-207-0681-4 .
- Ermenegildo Tremblay. Entomologia applicata . Volume III Parte I. 1ª ed. Napoli, Liguori Editore, 1991. ISBN 88-207-2021-3 .
- Ermenegildo Tremblay. Entomologia applicata . Volume III Parte II. 2ª ed. Napoli, Liguori Editore, 2005. ISBN 88-207-2335-2 .
- Ermenegildo Tremblay. Entomologia applicata . Volume III Parte III. 1ª ed. Napoli, Liguori Editore, 1997. ISBN 88-207-2632-7 .
- Aldo Pollini. Manuale di entomologia applicata . Bologna, Edagricole, 2002. ISBN 88-506-3954-6 .
Voci correlate
Altri progetti
-
 Wikizionario contiene il lemma di dizionario « Ditteri »
Wikizionario contiene il lemma di dizionario « Ditteri » -
 Wikimedia Commons contiene immagini o altri file su Ditteri
Wikimedia Commons contiene immagini o altri file su Ditteri -
 Wikispecies contiene informazioni su Ditteri
Wikispecies contiene informazioni su Ditteri
Collegamenti esterni
- ( EN ) Diptera , su Enciclopedia Britannica , Encyclopædia Britannica, Inc.
- ( EN , FR ) Diptera , su Enciclopedia canadese .
- ( EN ) Diptera , su Fossilworks.org .
- Ditteri , su Insetti.org , Mille-Animali.com. URL consultato il 9 dicembre 2008 .
- ( EN ) NL Evenhuis; T. Pape; AC Pont; FC Thompson, BDWD, BioSystematic Database of World Diptera , in Systema Dipterorum , Natural History Museum of Denmark, University of Copenaghen, 2009. URL consultato il 9 dicembre 2008 .
- ( EN ) Brian M. Wiegmann, David K. Yeates, Diptera. True Flies , su tolweb.org , The Tree of Life Web Project, 2007. URL consultato il 20 dicembre 2008 .
- ( EN ) Dipterists Forum. The Society for the study of flies (Diptera) , su dipteristsforum.org.uk . URL consultato il 9 dicembre 2008 .
- ( EN ) FLIES , su Public-Health Pest Control . URL consultato il 9 dicembre 2008 (archiviato dall' url originale il 27 dicembre 2008) .
- ( EN ) FLYTREE. Assembling the Diptera Tree of Life , su inhs.uiuc.edu . URL consultato il 9 dicembre 2008 .
- ( EN ) Diptera.info . URL consultato il 9 dicembre 2008 .
- ( EN ) WRBU, The Walter Reed Biosystematics Unit , su wrbu.si.edu . URL consultato il 9 dicembre 2008 (archiviato dall' url originale l'11 aprile 2009) .
- ( EN ) Division Nematocera , in Australian Faunal Directory , Australian Government, Department of the Environment, Water, Heritage and the Arts. URL consultato il 26 febbraio 2009 .
- ( EN ) Division Orthorrhapha , in Australian Faunal Directory , Australian Government, Department of the Environment, Water, Heritage and the Arts. URL consultato il 26 febbraio 2009 .
- ( EN ) Division Cyclorrhapha , in Australian Faunal Directory , Australian Government, Department of the Environment, Water, Heritage and the Arts. URL consultato il 26 febbraio 2009 .
- ( EN ) Taxon details: Diptera , in Fauna Europaea version 2.6.2 , Fauna Europaea Web Service, 2013. URL consultato il 09-12-2008 .
- ( EN ) Taxon profile: Diptera , in BioLib.cz, Biological Library . URL consultato il 09-12-2008 .
- ( EN ) ITIS Standard Report Page: Diptera , in Integrated Taxonomic Information System . URL consultato il 09-12-2008 .
- Ditteri. Macrofotografia ditteri diptera mosche , su Evasion.it, Fotografie di insetti e aracnidi . URL consultato il 9 dicembre 2008 .
| Controllo di autorità | Thesaurus BNCF 1496 · LCCN ( EN ) sh85049104 · GND ( DE ) 4068207-9 · BNF ( FR ) cb119373647 (data) · BNE ( ES ) XX530866 (data) · NDL ( EN , JA ) 00571460 |
|---|