Chimiotaxie
Chemotaxia ( neologism compus din χημεία cuvintele grecești, chemeia = chimia și τάξις, taxiuri = implementare), un tip de tacticitate, este fenomenul prin care organismele de celule , bacterii și alte organisme uni - sau mai multe -cellulari redirecționarea lor mișcări în în funcție de prezența anumitor substanțe chimice în mediul lor. Acest tip de taxie este foarte important pentru bacterii pentru a găsi hrană (de exemplu, glucoză ) înotând către concentrații maxime de molecule alimentare. La organismele multicelulare, chimiotaxia este critică în stadiile incipiente ale dezvoltării (de exemplu, mișcarea spermei spre ovul în timpul fertilizării), precum și în etapele ulterioare ale dezvoltării (de exemplu, migrarea neuronilor sau limfocitelor ), precum și în condiții normale funcții vitale. Mai mult, s-a recunoscut că mecanismele care permit chemotaxia la animale pot fi subvertizate în timpul metastazei cancerului .
Istoria cercetării chimiotaxiei

Deși migrația celulară a fost observată din cele mai vechi timpuri de dezvoltare a microscopului ( Leeuwenhoek ), prima descriere erudită a chemotaxiei a fost făcută de TW Engelmann (1881), Pfeffer (1884) asupra bacteriilor și HS Jennings (1906) asupra ciliaților. Laureatul Nobel Metchnikoff a contribuit, de asemenea , la acest domeniu de cercetare cu studii privind procesul văzut ca un pas inițial în fagocitoză.
Semnificația chimiotaxiei în biologie și patologie clinică a fost larg acceptată încă din anii 1930. Definițiile fundamentale referitoare la fenomen au fost schițate pentru prima dată în acei ani. Cele mai importante aspecte în controlul calității analizelor chemotactice au fost descrise de H. Harris în anii 1950. În anii 1960 și 1970, revoluția din biologia și biochimia celulară modernă a oferit o serie de noi tehnici disponibile pentru studierea celulelor de răspuns migrator și a fracțiunilor subcelulare responsabile de activitatea chimiotactică. Studiile de pionierat ale lui J. Adler au reprezentat un punct de cotitură semnificativ în înțelegerea întregului proces de transducție a semnalului intracelular la bacterii. [1]
La 3 noiembrie 2006, Dennis Bray de la Universitatea din Cambridge a primit distincția Microsoft European Science Award pentru munca sa de cercetare asupra chimiotaxiei în E. coli. [2]
Filogenie și semnale chimiotactice
Chimiotaxia este unul dintre cele mai elementare răspunsuri fiziologice celulare. Dezvoltarea sistemelor de recepție pentru recunoașterea substanțelor dăunătoare sau benefice în mediul înconjurător a fost esențială pentru organismele unicelulare încă din primele etape de dezvoltare. O analiză globală a activității chimiotactice a protozoarului eucariot Tetrahymena pyriformis și a secvențelor consensuale ale apariției aminoacizilor în supa primordială sugerează că a existat o bună corelație între caracterul chimiotactic al acestor molecule organice relativ simple și dezvoltarea lor pe Pământ. Prin urmare, s-a sugerat că primele molecule au fost aminoacizi puternic chemoatractivi (de ex. Gly, Glu, Pro), în timp ce aceștia din urmă formați s-au considerat că sunt puternic chimico-repelenți (de exemplu Tyr, Trp, Phe). [3]
Chimiotaxia în bacterii
Unele bacterii , cum ar fi E. coli, au numeroși flageli pe celulă (de obicei 4-10). Acestea se pot roti în două moduri:
- Rotația în sens invers acelor de ceasornic aliniază flagelele într-un singur pachet rotativ, astfel încât bacteria să înoate drept.
- Rotația în sensul acelor de ceasornic rupe mănunchiul de flageli, astfel încât fiecare flagel să indice într-o direcție diferită și, prin urmare, bacteria să cadă în loc.
Direcțiile de rotație se referă la un observator din afara celulei care privește flagelele spre celulă.

Comportament
Mișcarea de ansamblu a unei bacterii este rezultatul tumble alternativ și fazele de înot. Dacă te uiți la o bacterie care înoată într-un mediu uniform, mișcarea acesteia va apărea similară cu o mișcare aleatorie, cu înot relativ drept întrerupt de căderi aleatorii care reorientează bacteria. Bacteriile precum E. coli nu pot alege ce direcție să înoate și nu sunt capabile să înoate drept mai mult de câteva secunde din cauza difuziei rotaționale. Cu alte cuvinte, bacteriile „uită” în ce direcție merg. Cu aceste limitări, este remarcabil faptul că bacteriile sunt capabile să-și direcționeze mișcarea pentru a găsi locații favorabile cu concentrații mari de atractanți (de obicei alimente) și pentru a evita repelenții (de obicei otrăvuri).
În prezența unui gradient chimic, bacteriile vor chimiotaxiza sau își vor direcționa toată mișcarea pe baza gradientului. Dacă bacteria simte că se mișcă în direcția corectă (spre atractiv / departe de repulsiv), va continua să înoate drept înainte pentru o perioadă mai lungă de timp înainte de a se prăbuși . Dacă se mișcă în direcția greșită, va cădea mai repede și va căuta o nouă direcție la întâmplare, la întâmplare. Cu alte cuvinte, bacteriile precum E. coli folosesc senzația temporală pentru a decide dacă viața este mai bună sau mai rea. Procedând astfel, bacteria își găsește poziția în cea mai mare concentrație de atractiv (de obicei sursa). Chiar și în concentrații foarte mari, este capabil să distingă diferențe foarte mici de concentrație. Fugind de un agent de respingere, funcționează cu aceeași eficiență.
Pare remarcabil faptul că această „plimbare aleatorie” semnificativă este rezultatul unei alegeri simple între două metode de mișcare aleatorie: adică „căderea” și înotul drept. Într-adevăr, răspunsurile chimiotactice, cum ar fi uitarea unei direcții și alegerea mișcărilor, seamănă cu abilitățile decizionale ale formelor de viață superioare cu creierele care procesează date senzoriale.
Natura elicoidală a catenei flagelare unice este crucială pentru ca această mișcare să apară, iar proteina care formează catena flagelară, flagelina, este destul de similară între toate bacteriile flagelare. Vertebratele par să fi beneficiat de acest lucru, deoarece posedă un imunoreceptor (TLR5) capabil să recunoască această proteină extrem de conservatoare.
La fel ca în multe cazuri în biologie, există bacterii care nu respectă această regulă. Multe bacterii, cum ar fi Vibrio, sunt monoflagelate și au un singur flagel la un singur pol al celulei. Modul în care fac chimiotaxie este diferit. Alții posedă un singur flagel ținut în peretele celular. Aceste bacterii se mișcă rotind întreaga celulă, care arată ca un tirbușon. [4]
Transducția semnalului
Gradienții chimici sunt detectați prin intermediul mai multor receptori transmembranari, numiți proteine metilice care produc chimiotaxie (MCP), care variază în moleculele pe care le disting. Acești receptori se pot lega direct sau indirect de atrăgători sau repelenți prin interacțiuni cu proteinele din spațiul periplasmatic. Semnalele de la acești receptori sunt transmise prin membrana plasmatică în citosol, unde sunt activate proteinele chemotactice (proteinele Che). Aceste proteine modifică frecvența de tumbling și, de asemenea, modifică receptorii.
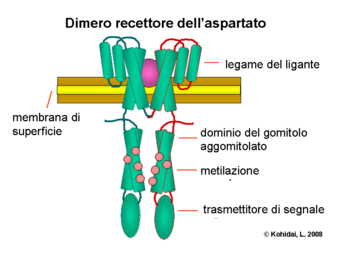
Reglementarea flagelilor
Proteinele CheW și CheA se leagă de receptor. Activarea receptorului de către un stimul extern determină întreruperea cascadei autofosforilative a histidin kinazei, CheA, activă în mod normal în absența unor substanțe atractive. CheA, la rândul său, transferă grupările fosforilate la reziduurile conservate de aspartat ca răspuns la regulatorii CheB și CheY [notă: CheA este o histidin kinază și nu transferă activ grupul fosforilat. Regulatorul de răspuns CheB ia grupul fosforilat de la CheA]. Acest mecanism de transducție a semnalului se numește „Sistem cu două componente” și este o formă comună de transducție a semnalului la bacterii. CheY fosforilat induce căderea prin interacțiunea cu proteina declanșatoare flagelară FliM, determinând rotația flagelului să se schimbe de la sens invers acelor de ceasornic la sensul acelor de ceasornic. Schimbarea stării de rotație a unui singur flagel poate sparge întregul pachet de flageli și poate provoca o cădere . în prezența unor substanțe atractive, pe de altă parte, se constată că receptorii MCP (proteine chemotaxice care acceptă metil) sunt încărcați și CheA nu întrerupe autofosforilarea acestuia. În acest fel, proteina CheY nu este fosforilată și nu mai interacționează cu proteinele flagel FliM. Lipsa de interacțiune între aceste două proteine are ca rezultat o mișcare de „alergare” de către celula bacteriană care va continua să se deplaseze în direcția din care a perceput substanța chimio-atrăgătoare.
Reglementarea receptorilor
CheB, atunci când este activat de CheA, acționează ca o metilesterază, eliminând grupările metil din reziduurile de glutamat de pe partea citosolică a receptorului . Funcționează într-un mod antagonist cu CheR, o metiltransferază, care adaugă reziduuri de metil la aceleași reziduuri de glutamat. Cu cât sunt mai multe reziduuri de metil atașate la receptor, cu atât receptorul este mai sensibil. În timp ce semnalul de la receptor induce demetilarea receptorului într-o buclă de feedback, sistemul este ajustat continuu la nivelurile chimice de mediu, rămânând sensibil la modificări mici chiar și în concentrații chimice extreme. Această ajustare permite bacteriei să „amintească” concentrațiile chimice din trecutul recent și să le compare cu ceea ce se confruntă în prezent, astfel încât să „știe” unde merge deasupra sau sub un gradient . Cu toate acestea, sistemul de metilare singur nu poate explica gama largă de sensibilitate pe care o au bacteriile față de gradienții chimici. Mecanisme de reglare suplimentare, cum ar fi gruparea receptorilor și interacțiunile receptor-receptor, modulează, de asemenea, modelul semnalului.

Chimiotaxie eucariotă
Mecanismul prin care celulele eucariote produc chimotaxia este complet diferit de cel al bacteriilor; cu toate acestea, detectarea gradienților chimici este întotdeauna un pas crucial în proces. Datorită mărimii lor, procariotele nu sunt în măsură să recunoască gradienții de concentrație eficienți, prin urmare, aceste celule își monitorizează și evaluează mediul înotând în mod constant (atacuri consecutive drepte și accidentale). Spre deosebire de procariote, dimensiunea celulelor eucariote oferă posibilitatea de a distinge gradienții și, prin urmare, se realizează o distribuție dinamică și polarizată a receptorilor. Inducerea acestor receptori de către chemoatractanți sau chemorepellants duce la migrarea către sau din substanța chemotactică.
Nivelurile receptorilor, modurile de semnalizare intracelulară și mecanismele efectoare reprezintă toate componente diferite ale tipului eucariot . În organismele eucariote unicelulare, mișcarea ameboidă și ciliul sau flagelul eucariot sunt principalii efectori (de exemplu, Amoeba sau Tetrahymena) [5] [6] . Unele celule eucariote ale vertebratelor mai evoluate , cum ar fi imunocitele , se mută acolo unde sunt necesare în același mod ca organismele eucariote unicelulare. În plus față de celulele imun-competente ( granulocite , monocite , limfocite ), un grup mare de celule considerate anterior fixate în țesuturi, sunt, de asemenea, mobile în anumite condiții fiziologice (de exemplu, mastocite , fibroblaste , celule endoteliale) sau patologice (de ex. metastaze). Chimiotaxia are o semnificație importantă în stadiile incipiente ale embriogenezei, deoarece dezvoltarea straturilor germinale este condusă de gradienții moleculelor semnal.
Motilitate
Spre deosebire de motilitatea din chemotaxia bacteriană, mecanismul prin care celulele eucariote se mișcă fizic nu este clar. Se pare că există mecanisme prin care un gradient chimiotactic extern este detectat și transformat într-un gradient intracelular PIP3, care are ca rezultat un gradient în activarea modului de semnal care culminează cu polimerizarea filamentelor de actină. Terminația de actină în creștere distală dezvoltă conexiuni cu suprafața interioară a membranei plasmatice prin diferite grupuri peptice și duce la formarea pseudopodiei . Ciliul celulei eucariote poate duce, de asemenea, la chemotaxie, dar în acest caz este în principal o inducție dependentă de calciu a sistemului microtubular al corpurilor bazale și pachetul de 9 x 2 + 2 al microtubulilor cililor. Bătaia orchestrată a sute de cili este sincronizată de un sistem submembranar construit între corpurile bazale. Detaliile modului de semnal nu sunt încă pe deplin clare.
Răspunsuri migratorii legate de chimiotaxie
Deși chimiotaxia este cea mai frecventă formă de migrație studiată, există mai multe alte forme de locomoție la nivel celular.
- Chimiochineza este indusă și de moleculele fazei lichide a mediului înconjurător; în orice caz, este răspunsul exprimat într-o taxie non-vectorială aleatorie. Nici amplitudinea, nici frecvența mișcării nu au componente direcționale caracteristice tipice acestui comportament.
- În haptotaxie , gradientul chemoattractantului este exprimat sau legat de suprafață, spre deosebire de calea clasică a chemotaxiei în care gradientul se dezvoltă într-un spațiu solubil. Principala suprafață haptotactică biologic activă este matricea extracelulară (ECM); prezența liganților legați este responsabilă de inducerea migrației transendoteliale și a angiogenezei.
- Necrotaxia reprezintă un tip special de chimiotaxie în care moleculele chimiotratante sunt eliberate din celulele necrotice sau apoptotice . În funcție de natura chimică a substanțelor eliberate, necrotaxia poate acumula sau respinge celule, ceea ce subliniază semnificația fiziopatologică a acestui fenomen.

Receptorii
În cea mai mare parte, celulele eucariote simt prezența stimulilor chimiotactici prin intermediul receptorilor cuplați cu proteine G 7-transmembranare (sau serpentine) heterotrimerice. Această clasă de receptori are o mare semnificație și reprezintă o porțiune semnificativă a genomului . Unii membri ai acestei superfamilii de gene sunt folosiți în viziune (rodopsine), precum și în abilitățile olfactive. receptori de leucotriene (BLT); cu toate acestea, inducerea unui grup mare de receptori de membrană (de exemplu, aminoacizi , insulină , peptide vasoactive) permite, de asemenea, migrarea celulară.
Selecția chimiotactică
Unii receptori chimiotactici sunt exprimați pe suprafața membranei cu caracteristici pe termen lung, deoarece sunt determinați genetic, în timp ce alții au o dinamică pe termen scurt, deoarece sunt asamblați ad hoc în prezența ligantului. Diferitele caracteristici ale receptorilor și liganților de chemotaxie oferă posibilitatea de a alege celulele de răspuns chemotactic cu o analiză chimiotactică simplă. Cu selecția chimiotactică putem determina dacă o moleculă care nu este încă caracterizată acționează printr-o cale de receptor pe termen lung sau scurt. Termenul de selecție chimiotactică este, de asemenea, utilizat pentru a desemna o tehnică care separă celulele eucariote de celulele procariote în funcție de capacitatea lor de reacție la liganții selectivi. [7]

Liganti chimiotactici
Numărul de molecule capabile să exprime răspunsuri chimiotactice este relativ mare și putem distinge moleculele chimiotactice primare și secundare. Principalele grupe de liganti primari sunt urmatoarele:
- Peptide formil: sunt di-, tri-, tetra-peptide de origine bacteriană (vezi gruparea formil de la capătul N al peptidei). Ele sunt eliberate de bacterii in vivo sau după descompunerea celulelor. Un membru tipic al acestui grup este N-formilmetionil-leucil-fenilalanina (fMLF sau fMLP în bibliografie). FMLF bacterian ca o componentă cheie a inflamației are efecte caracteristice de chimiotratare asupra granulocitelor și monocitelor neutrofile.
- Complementul 3a (C3a) și complementul 5a (C5a): sunt produse intermediare ale unei cascade de complemente. Sinteza lor este combinată cu trei moduri alternative (clasice, dependente de lectină și alternative) de activare a complementului printr-o enzimă convertază. Principalele celule țintă ale acestor derivați sunt granulocitele neutrofile, precum și monocitele.
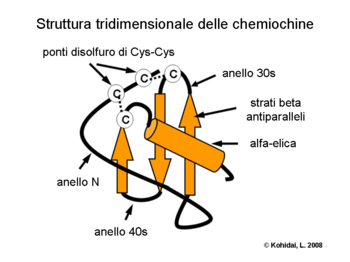
- Chimiochine: aparțin unei clase speciale de citokine. Grupurile lor (C, CC, CXC, CX3C) reprezintă molecule nu numai legate structural și cu un aranjament special de punți disulfidice, dar specificitatea lor ca celule țintă este, de asemenea, diferită: chemokinele CC acționează asupra monocitelor (de exemplu, RANTES), chemokinele CXC sunt specifice pentru granulocitele neutrofile (de ex. IL-8).

Cercetările asupra structurilor tridimensionale ale chemokinelor au arătat că o compoziție caracteristică a unei spirale exprimă secvențele necesare interacțiunii cu receptorii chemokinelor. Formarea dimerilor și creșterea activității biologice a acestora a fost demonstrată cu cristalografia mai multor chemokine (de exemplu IL-8).
- Leucotriene: aparțin grupului de eicosanoizi, care sunt importanți mediatori lipidici ai reacției acidului arahidonic prin 5-lipoxigenază. Componenta lor predominantă este leucotriena B4 (LTB4) care permite aderența, chimiotaxia și agregarea leucocitelor. Efectul caracteristic chimiotratant al LTB4 este indus de proteina G legată de receptorii leucotrienei 7-transmembranari, care sunt puternic exprimate în inflamație și alergie .
Adaptări ale gamelor de acțiune chimiotactice (CRF)
Răspunsurile chimiotactice din interacțiunile ligant-receptor se disting în general prin concentrația (efectele) optimă (e) a ligantului (e). Cu toate acestea, corelația amplitudinii exprimate și raportul celulelor de răspuns în comparație cu numărul total sunt, de asemenea, trăsături caracteristice ale semnalului chemotactic. Studiile asupra familiilor liganților (de exemplu, aminoacizi sau oligopeptide au arătat că există o adaptare a gamei de acțiune (amplitudini; număr de celule de răspuns) și a activităților chimiotactice: mediul chemoatractant este însoțit de game largi de acțiune, în timp ce chemorepelul calități ale unei game limitate de acțiune.

Semnificația clinică
Un potențial migrator mutant al celulelor are o importanță relativ mare în dezvoltarea mai multor simptome clinice și sindroame. Activitatea chimiotactică modificată a agenților patogeni extracelulari (de exemplu, Escherichia coli ) sau intracelulară (de exemplu, Listeria monocytogenes ) este ea însăși o țintă clinică semnificativă. Modificarea capacității chimiotactice endogene a acestor microorganisme de către agenții farmaceutici poate reduce sau inhiba relația infecțiilor sau răspândirea bolilor infecțioase. În afară de infecții, există și alte boli în care chimiotaxia scăzută este factorul etiologic primar, cum ar fi sindromul Chédiak-Higashi în care veziculele intracelulare gigantice inhibă migrația normală a celulelor.
| Tipul bolii | Chtx. crescut | Chtx. scăzut |
|---|---|---|
| Infecții | Inflamații | SIDA , bruceloză |
| Chtx. determină boala | - | Sindromul Chediak-Higashi, sindromul Kartagener |
| Chtx. este afectat | Ateroscleroză , artrită , parodontită , psoriazis , defect de reperfuzie, tumori metastatice | Scleroză multiplă , limfom Hodgkin , infertilitate masculină |
| Intoxicatii | Azbest , benzopiren | Săruri de Hg și Cr, ozon (O 3 ) |
În procesul de publicare
Cercetările privind migrația celulară - așa cum se arată în capitolul „Istoria cercetării chemotaxiei” - necesită aplicarea complementară a tehnicilor clasice și moderne. Arhiva oferă posibilitatea de a prezenta noi date care pot fi evaluate atât în cercetarea de bază, cât și în științele aplicate. În ultimii 20-25 de ani, datorită factorilor menționați mai sus, a existat o creștere a numărului de publicații care se ocupă cu fenomenul de chemotaxie în sine. Cu toate acestea, alte publicații din domeniile geneticii , biochimiei , fiziologiei celulare, patologiei și științelor clinice ar putea prezenta, de asemenea, date despre migrarea celulelor sau, în special, despre chimiotaxia celulară. O curiozitate cu privire la cercetarea migrației constă în faptul că printre numeroasele lucrări asupra taxiei (de exemplu, termotaxie , geotaxie , fototaxie ) cercetările privind chemotaxia arată un raport semnificativ ridicat, care tinde să sublinieze importanța cercetării chimiotaxice atât în biologie, cât și în medicină.

Măsurarea chimiotaxiei
O gamă largă de tehnici este acum disponibilă pentru a evalua activitatea chimiotactică a celulelor sau caracterul chimiotratant și chimio-repulsiv al liganților. Practic, ceea ce este necesar pentru efectuarea măsurătorilor este:
- Gradienții de concentrare se pot dezvolta relativ rapid și pot persista mult timp în sistem
- Activitățile chimiotactice sunt distincte de cele chimiocinetice
- Migrația celulară este liberă pe axa gradientului de concentrație spre sau departe de punctul de concentrație maximă
- Răspunsurile observate sunt rezultatele migrării active a celulelor
Deși un experiment chemotactic ideal nu este încă disponibil, există mai multe protocoale și instrumente care oferă o potrivire bună cu condițiile descrise mai sus. Cele mai frecvent utilizate sunt:
- Experimente pe plăci de agar (de exemplu, camere PP)
- Tehnici cu două camere (de exemplu, camera Boyden, camera Zigmond, camera Dunn, camere cu mai multe canale, tehnici capilare)
- Altele (de exemplu, tehnica labirintului T, tehnica opalescenței, experimentele de orientare)
Notă
- ^ Julius Adler și Wung-Wai Tso, luarea deciziilor în bacterii: răspuns chimotactic al Escherichia Coli la stimuli conflictuali , în Știință , vol. 184, 1974, pp. 1292–4, DOI : 10.1126 / science.184.4143.1292 , PMID 4598187 .
- ^ BBC NEWS | Știință / natură | Studiul de erori pe computer câștigă premiul cel mai mare preluat la 6 noiembrie 2006
- ^ Ligandul chemotactic „optim” - aminoacizi Depus pe 29 septembrie 2008 în Internet Archive . preluat 3 februarie 2008
- ^ Howard C. Berg, E. coli „în mișcare” , Springer-Verlag, NY. , 2003. ISBN 0-387-00888-8
- ^ Anna Bagorda, Carole A. Parent, Chimiotaxia eucariotă dintr-o privire , în J. Cell Science , vol. 121, 2008, pp. 2621-4.
- ^ Laszlo Kohidai, Chemotaxis: răspunsul fiziologic adecvat pentru evaluarea filogeniei moleculelor de semnal , în Acta Biol Hung , vol. 50, 1999, pp. 375-94, PMID 10735174 .
- ^ Laszlo Kohidai și Gyorgy Csaba, chemotaxie și selecție chemotactică indusă cu citokine (IL-8, RANTES și TNF alfa) în Tetrahymena pyriformis unicelulară. , în Cytokine , vol. 10, 1988, pp. 481–6, DOI : 10.1006 / cyto.1997.0328 , PMID 9702410 .
Alte proiecte
-
 Wikționarul conține dicționarul lema « chimiotaxie »
Wikționarul conține dicționarul lema « chimiotaxie » -
 Wikimedia Commons conține imagini sau alte fișiere despre chimiotaxie
Wikimedia Commons conține imagini sau alte fișiere despre chimiotaxie